


Abstract
Background/Aim: Intrauterine growth retardation (IUGR) causes very low birth weight and is related to the morbidity and mortality of the newborn. In our previous study, expression of brain-derived neurotrophic factor (BDNF) was found reduced in the cerebral cortex and dentate gyrus of fetuses with IUGR. BDNF protected cortical neurons against hypoxic injury via activation of the extracellular signal-related kinase (ERK) pathway. The aim of the current study was to observe the immunoreactivity of ERK in mature neurons and proliferating cells. Materials and Methods: Uterine artery ligation was performed at 17 days of gestation (dg). Rat fetuses were obtained at 21 dg using cesarean section. Fetuses were designated either to the growth retardation (GR) group when removed from the horn with uterine artery ligation, or to the control group when removed from the other horn with the untied artery. Immunohistochemistry was performed with primary antibodies on paraffin-embedded forebrain sections. Results: The density and proportion of cells expressing PCNA, ERK, and phosphate ERK in the subventricular zone (SVZ) was not different between the control and GR group. The density and proportion of NeuN- and phosphate ERK-positive cells in the cerebral parietal cortex was lower in the GR group, compared to the control group. Conclusion: Although IUGR had no effect on the proliferation of cells in the SVZ, it reduced neuronal survival in the cerebral parietal cortex, which was associated with the decrease of pERK-positive cell density and proportion in the cerebral cortex.
Intrauterine growth retardation (IUGR) causes very low birth weight (1) and is related to the morbidity and mortality of newborn infants. A majority of pregnancies with IUGR symptoms result in stillbirth or neonatal death (2). Furthermore, IUGR is generally associated with placental insufficiency (3) and is caused by abnormalities in the umbilical artery (4). The common symptoms in fetuses undergoing placental insufficiency include malnutrition, hypoxemia, and hypoglycemia (5).
The uterine artery ligation animal model, with placental insufficiency and IUGR (6), is a good representation of the human IUGR condition (7). This model has been used to demonstrate metabolic and hemodynamic abnormalities during cardiac (8), vascular (9), and neurological development (10, 11). Some studies state that the growth of the majority of infants with IUGR is normalized by the age of 2-3 years (12, 13). However, other studies showed that IUGR caused neurological disorders such as cerebral palsy (14), schizophrenia (15), and problems with learning/memory (16).
IUGR is also associated with neurogenesis, which is a process that encompasses the proliferation, growth, migration, and survival of neuronal cells. The number of neurons was reduced in the hippocampus and cerebellum of postnatal fetuses (17, 18). Further, morphological changes were seen during the prenatal period in IUGR fetuses (19) and the neocortical volume was reduced in IUGR neonates (20).
In our previous study (21), cell survival was reduced in the cerebral cortex and dentate gyrus of fetuses with hypoxia. In addition, the expression of brain-derived neurotrophic factor (BDNF) was also decreased in the same regions (21). BDNF plays an important role in neuronal cell proliferation and survival (22). Another study demonstrated that BDNF protected cortical neurons against hypoxic injury via activation of the extracellular signal-related kinase (ERK) pathway (23). ERK activation promoted the survival of pheochromocytoma 12 (24) and retinal ganglion cells (25).
The aim of this study was to observe the immunoreactivity of ERK and phosphate-ERK (pERK) in mature neurons and proliferating cells.
Materials and Methods
Animal surgery. The Chosun University Institutional Animal Care and Use Committee approved all animal experiments (approval number: CIACUC2014-A0026). Sprague-Dawley (SD) rats were supplied from a certified breeder (Damul Laboratory Animals, Republic of Korea). The rats were mated for three days. Female rats were then checked vaginally to determine the pregnancy status. Uterine artery ligation was performed as reported in a previous study (26). Briefly, animals were anesthetized by intramuscular injection with zoletil (10 mg/kg; Virbac, France) and xylazine (0.15 mg/kg; Bayer, Germany) at 17 days of gestation (dg). After shaving below the umbilicus, we performed a midline incision under aseptic surgical conditions. The uterine horn was found, and the uterine artery at the cervical level was ligated with silk sutures (4/0). After the procedure, the skin on abdominal region was sterilized using povidone-iodine solution (Green Medical, Republic of Korea). All animals were bred in the same environment.
Tissue preparation. At 21 dg, we reopened the surgical site of pregnant rats (n=14) and obtained fetuses by cesarean section. Fetuses that were removed from the horn with the artery ligation were designated to the growth retardation (GR) group (n=45), while fetuses that were removed from the other horn with the untied artery served as the control group (n=47). We measured body and brain weights, crown-rump length, and the brain-to-body-weight ratio of both fetus groups. The brains of the fetuses were removed, fixed in 4% paraformaldehyde (PFA) solution, and washed after two days in distilled water. The brains were dehydrated in a series of graded ethanol and embedded in paraffin. Serial coronal sections were cut at 12 μm thickness and mounted on gelatin-coated slides (Fisher Scientific, Pittsburgh, PA, USA).
Immunohistochemistry. The sections were deparaffinized, washed with 0.1 M phosphate buffered saline (PBS; pH 7.4), and heated in a microwave for 10 min with 0.01 M sodium citrate buffer (pH 6.0) for antigen retrieval. After cooling for 40 min, the sections were soaked in 0.3% hydrogen peroxide solution for 20 min to block endogenous peroxidase. After washing with PBS, sections were then immersed with primary antibody solutions and incubated overnight at 4°C. The next day, the sections were rinsed with PBS and incubated with the appropriate secondary antibody solutions, which matched the type of primary antibody. The immunoreactivity (IR) of cells in the sections was visualized using an avidin-biotin-peroxidase (ABC) detection system (Vectastain ABC Elite Kit, Vector Laboratories). Counterstaining was performed with thionin. The sections were mounted with PolyMount mounting medium (Polysciences, USA).
The primary antibodies used were as follows: mouse anti-hexaribonucleotide-binding protein 3 (NeuN, 1:100; Millipore, USA), mouse anti-proliferating cell nuclear antigen (PCNA, 1:1,000; Sigma-Aldrich, USA), rabbit anti-ERK (1:50; Santa Cruz, USA), and goat anti-p-ERK (1:50; Santa Cruz, USA).
Body and brain weight and crown-rump length of control and GR groups at 21 days of gestation.
Quantification. Immunoreactivity of the sections was visualized using a light microscope (Olympus BX41, USA) connected to a digital CCD camera. Coronal sections separated by a distance of 240 μm were obtained from each animal. The density of cells expressing NeuN, PCNA, ERK, and pERK was determined by counting the number of immunoreactive cells in a defined field (objective: 400×), as described previously (27), in five randomly selected areas in the granular layer of cerebral parietal cortex and SVZ. The proportion of these cells was expressed as a percentage of the total number of cells in each region. IR cells were manually counted using double blind protocols.
Statistical analysis. All data were analyzed using Statistical Package for Social Sciences (Information Analysis Systems, SPSS, USA). All measurements were compared between the control and GR groups using Student's t-tests. The statistical significance was set at p<0.05.
Results
At 21 dg, body and brain weights in the GR group were decreased relative to those of the control group. The ratio of brain-to-body weight in GR fetuses was also increased relative to that of controls (Table I). At 21 dg, the density and proportion of PCNA-IR cells in the subventricular zone (SVZ) was the same in the control and GR groups (Figure 1). The density and proportion of NeuN-IR cells in the cerebral parietal cortex was lower in the GR group, in comparison to the control group (Figure 2). There were no differences in the density and proportion of ERK-IR cells in the cerebral cortex and SVZ, between the control and GR groups (Figure 3). Although the density and proportion of pERK-IR cells of the GR group did not differ from the control group in the SVZ, it decreased in the cerebral parietal cortex (Figure 4).
Discussion
Unilateral artery ligation was used to induce hypoxia in pregnant rats at 17 dg. This time-point is the onset of the period of rapid fetal growth and matches the third trimester in human pregnancy (28). Uterine artery ligation clearly decreased uterine blood flow and caused fetal weight reduction (29), which resulted in neuronal cell death (30). Further, impaired neuronal cell bodies were observed in the hippocampus and cerebral cortex (31), and were reduced in the entorhinal cortex (32). Hypoxic injury, like uterine artery ligation, was also associated with oligodendrocyte maturation (33) and defects in myelination (34). These deficits were related with neurobehavioral problems, including motor hyperactivity, posture abnormalities, and working memory deficit (35, 36).

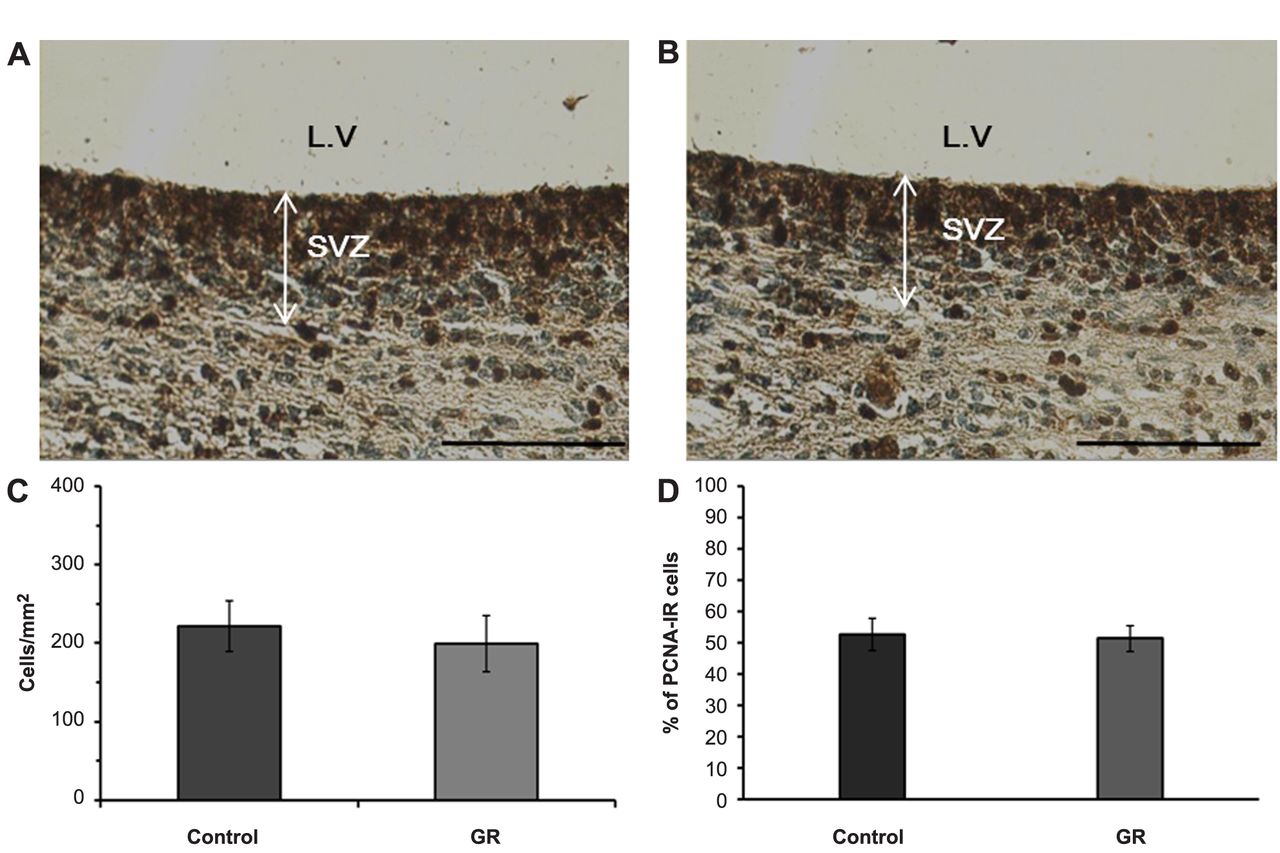
Representative photomicrographs of PCNA-IR cells in the SVZ. PCNA-IR cells were observed as dark-brown color in the control (A) and GR (B) groups. Scale bars=100 μm. The density (C) and proportion (D) of PCNA-IR cells in the SVZ of control and GR fetuses at 21 days of gestation. No significant difference in the density and proportion of IR cells was observed between the two groups. Values are expressed as the mean±SEM.
In the current study, immunoreactivity was observed in two regions: the SVZ and cerebral parietal cortex. The SVZ is a neurogenic zone that responds to hypoxic injury during the late stage of brain development (37). This area is associated with neuronal cell proliferation (38). The parietal cortex is more vulnerable to insult than other cortices. For example, although the primary motor cortex in rats with placental insufficiency did not show neuronal density reduction (11), the parietal cortex in rats with a similar condition showed structural abnormality (39). Finally, neuronal survival in the cerebral cortex of fetuses has been previously associated with the ERK pathway (40).
In the current study, we used a PCNA antibody to determine cell proliferation in the SVZ, since it is a nuclear protein since it is an essential element that binds polymerase-d during leading-strand DNA synthesis (41). PCNA expression appears during the G1 phase and is maximal during the S phase (42). Although the density of PCNA-IR cells in the GR group was a little lower than in the control group, this difference was not statistically significant. Previous studies indicate the reduction of neural progenitor cells after hypoxic damage and the presence of immature neuronal cells or apoptotic cells (38, 43). Tolcos et al. (27) found that although cell proliferation increased, the weight of the brain was decreased. These results suggest the variability of the intrauterine micro-environment.
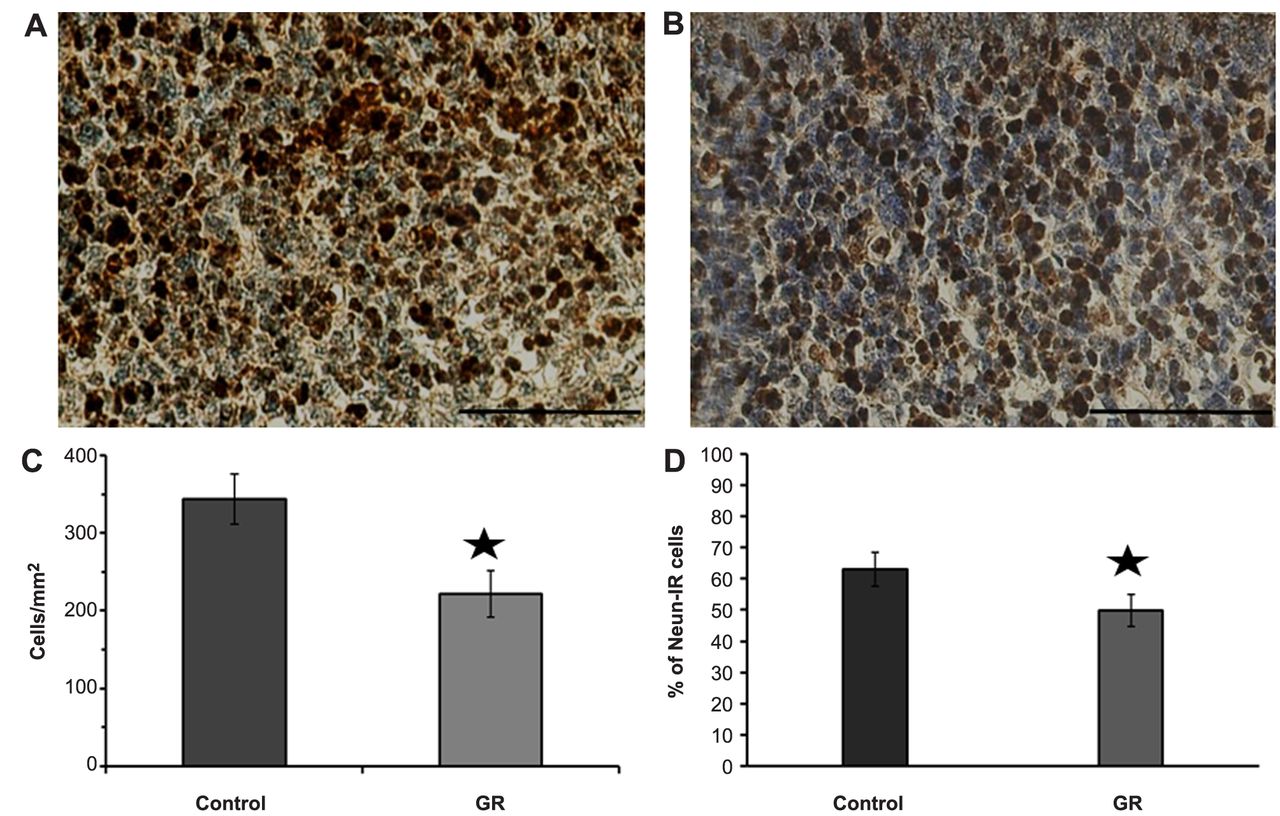
Next, we used NeuN antibody to determine the density of mature neurons in the cerebral parietal cortex. NeuN is a monoclonal antibody, and its nuclear antigen is detected during neurogenesis progress and is restricted to neurons (44). The density of NeuN-IR cells in the cerebral parietal cortex decreased in the GR group. This result was similar to our previous study, using a guinea pig model that showed chronic placental insufficiency (21, 45). Similarly, the density of NeuN-IR cells was significantly reduced in the developing chick brain (46). These results suggest that the survival of neurons in the cerebral cortex was reduced in GR fetuses.
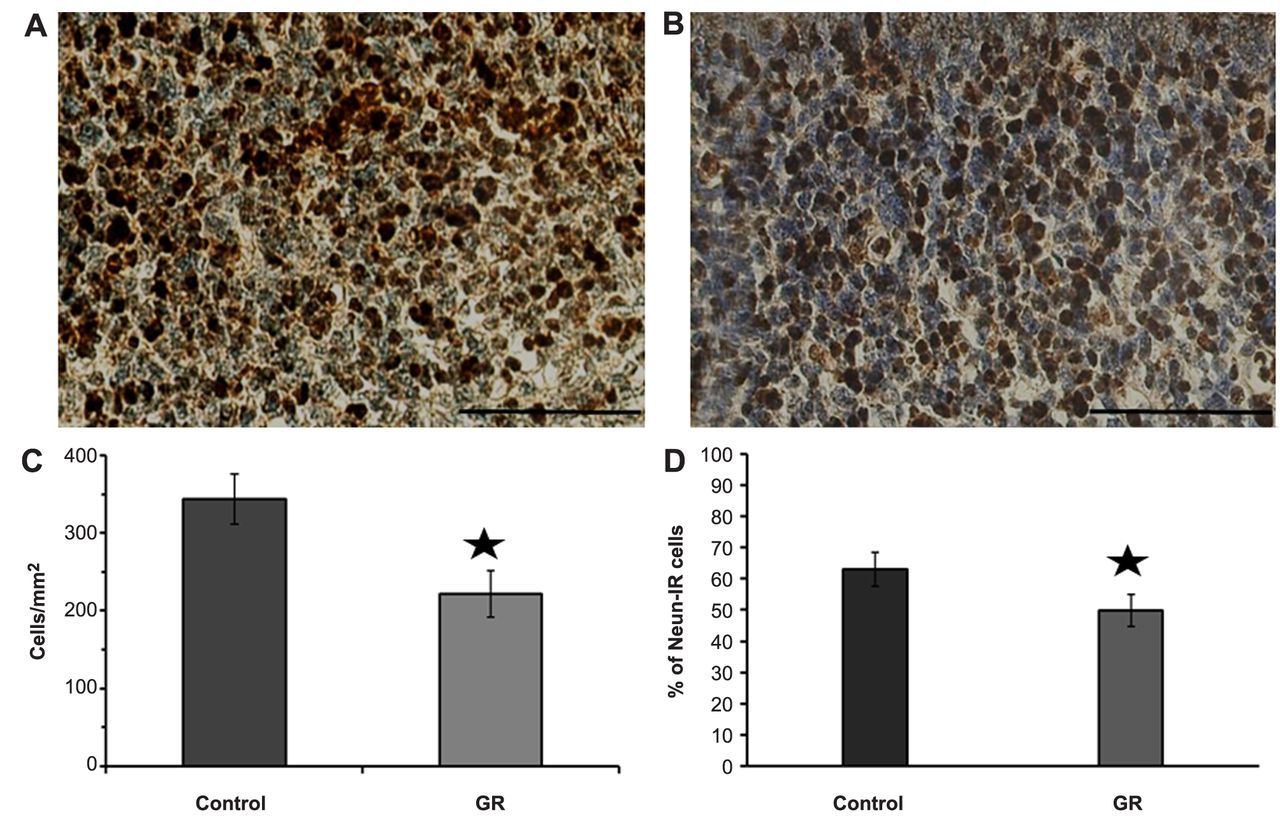
Representative photomicrographs of NeuN-IR cells in the granular layer of the l parietal cortex. NeuN-IR cells (dark-brown color) were scattered in both the control (A) and GR (B) groups. Scale bars=100 μm. The density (C) and proportion (D) of NeuN-IR cells in the cerebral parietal cortex of control and GR fetuses at 21 days of gestation. The density and proportion of IR cells was lower in the GR group than in the control group. Values are expressed as the mean±SEM. *p<0.05.
ERK 1/2 are members of the mitogen-activated protein kinase (MAPK) family (47), which elicits an intracellular signaling cascade, i.e., the Ras-Raf-MEK-ERK signaling pathway, which is involved in the development of neurons (48). Activation of the ERK pathway was modulated by extracellular stimuli, and affected cell proliferation and differentiation (49). In the current study, the density of pERK in the SVZ was the same between control and GR groups. Since the neuronal cell loss in IUGR is normalized after a few years, it is possible that the ERK pathway triggers neural progenitor cell differentiation into neural cells (50). Therefore, simply increasing the activation ERK in an attempt to compensate for hypoxic damage is not enough. The density of pERK and mature neuronal survival was reduced in the cerebral parietal cortex in the GR group, in comparison to the control. Since the activation of ERK by phosphorylation promoted cell survival in cortical neurons (23), we suggest that the reduction of pERK immunoreactivity is associated with the decreased neuronal survival in the cerebral cortex. Neurotrophin factors protected cortical neurons by ERK activation (51). ERK activation has effects on early human neurogenesis (52) and cancer clinicopathologic (53). So, the results about ERK phosphorylation are associated with therapeutic window (54). There are two representative protection mechanisms that are elicited by ERK-activated BDNF, the inhibition of pro-apoptotic proteins (55) and the induction of CRE-binding protein phosphorylation (56).
Conclusion
IUGR induced by uterine artery ligation leads to reduction of neuronal survival in the cerebral parietal cortex, but not to the reduction of proliferation in SVZ. In the cerebral cortex, the reduction in neuronal survival is associated with the decrease of cells expressing pERK.
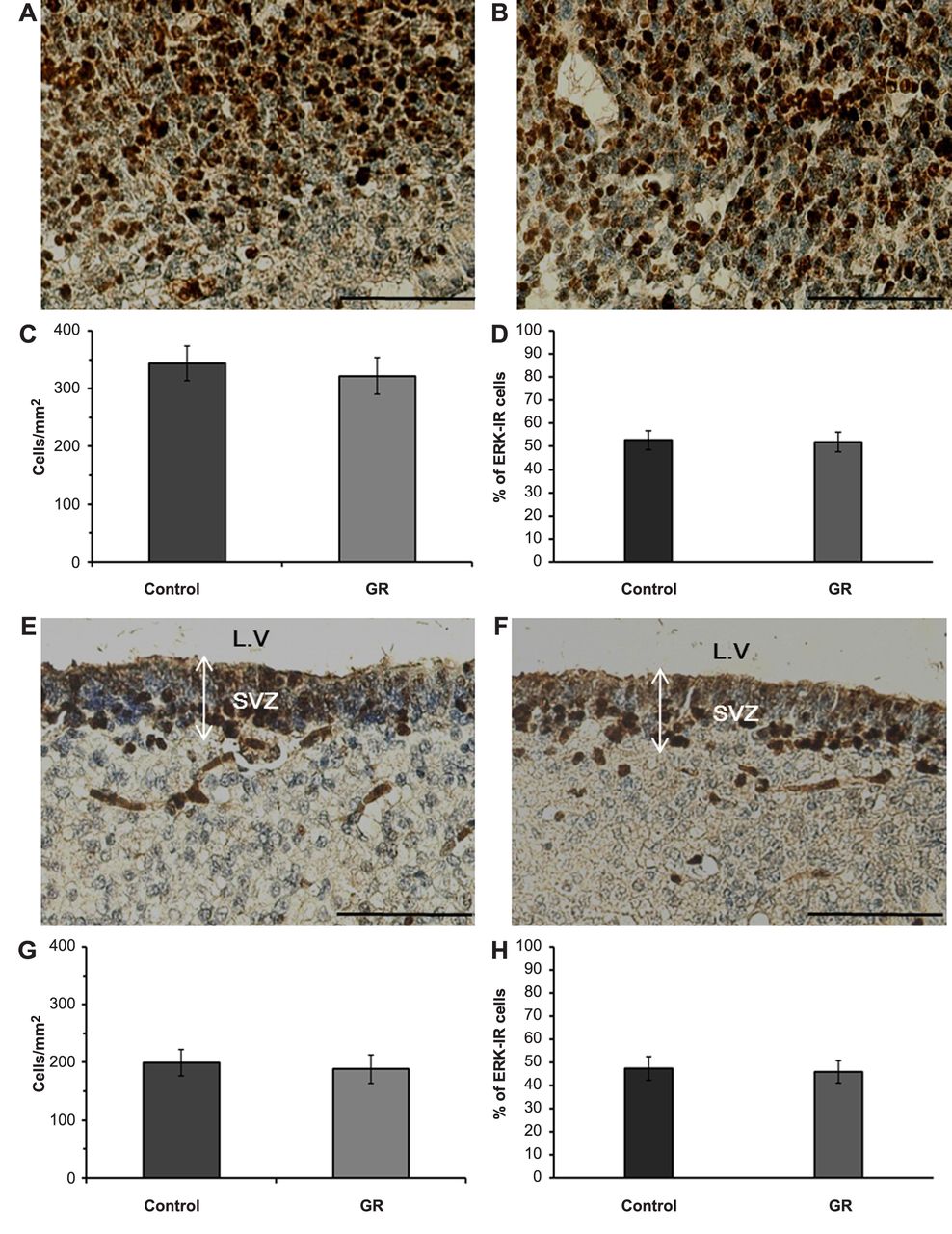
Representative images of ERK-IR cells (dark-brown) in the cerebral parietal cortex (A: control B: GR) and SVZ (E: control F: GR) at 21 days of gestation. Scale bars=100 μm. The density (C: cerebral cortex G: SVZ) and proportion (D: cerebral cortex H: SVZ) of ERK-IR cells in the control and GR groups at 21 days of gestation. No significant differences in the density and proportion of ERK-IR cells in the cerebral parietal cortex and SVZ were observed between the two groups.
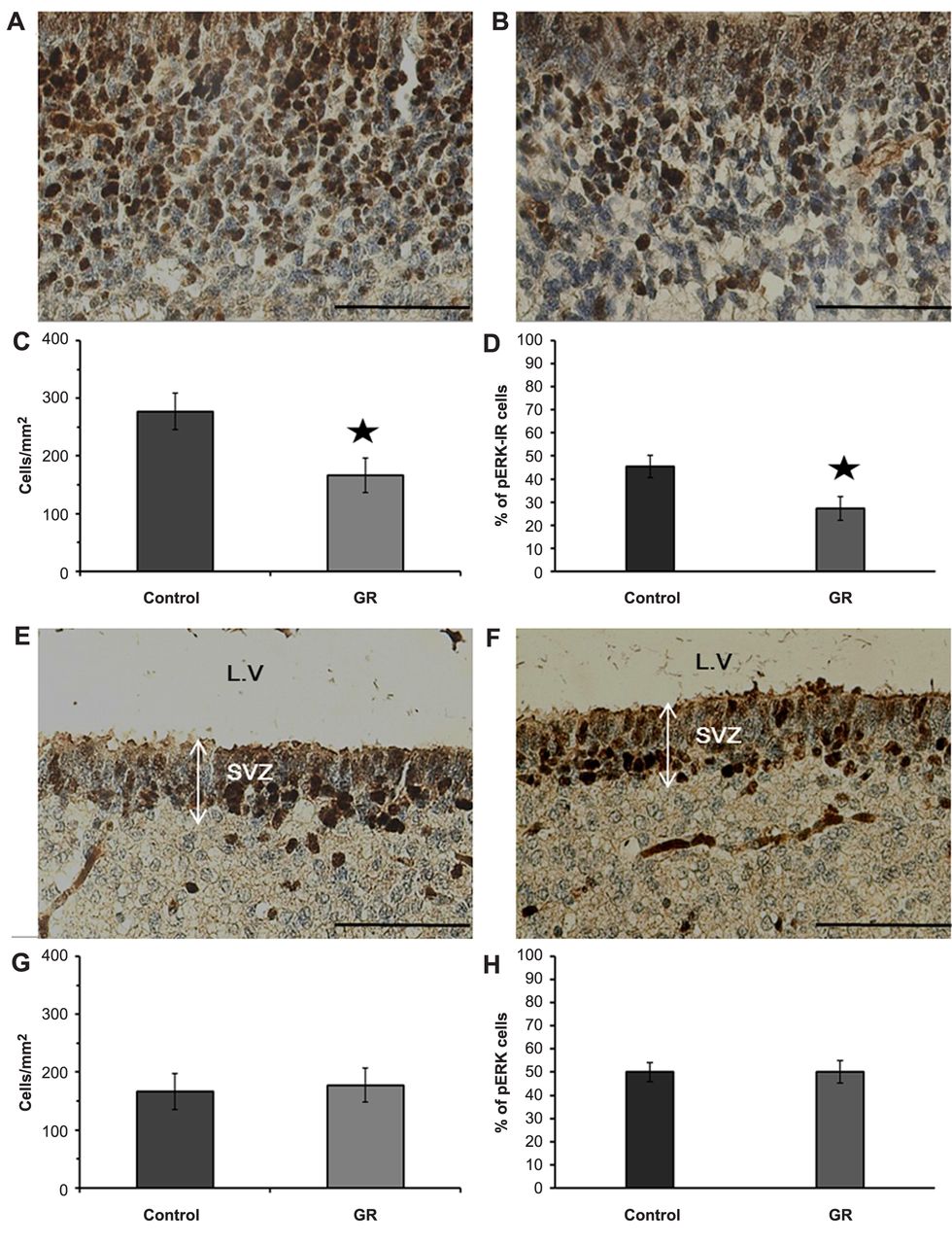
Analysis of pERK-IR cells (dark-brown) in the cerebral parietal cortex (A: control B: GR) and SVZ (E: control F: GR) at 21 days of gestation. Scale bars=100 μm. The density (C: cerebral cortex G: SVZ) and proportion (D: cerebral cortex H: SVZ) of pERK-IR cells in the control and GR groups at 21 days of gestation. The density and proportion of pERK-IR cells were decreased in the cerebral parietal cortex of GR fetuses, relative to those of controls. Values are expressed as the mean±SEM. *p<0.05.
Acknowledgements
This study was supported by research funds from Chosun University (2015).
Footnotes
Authors>' Contributions
JYH designed the study. KST participated in the surgical procedures. KDJ analyzed the obtained data. CYY and CMS performed the immunohistochemistry. All Authors read and approved the final manuscript.
This article is freely accessible online.
Conflicts of Interest
The Authors declare that no competing interests exist.
- Received December 10, 2019.
- Revision received December 21, 2019.
- Accepted January 5, 2020.
- Copyright© 2020, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Dexmedetomidine Stereotaxic Injection Alleviates Neuronal Loss Following Bilateral Common Carotid Artery Occlusion via Up-Regulation of BDNF Expression
- Differential Expression of Pax6 Following Bilateral Common Carotid Artery Occlusion
- Differential Expression of BDNF and BIM in Streptozotocin-induced Diabetic Rat Retina After Fluoxetine Injection