Article Text

Abstract
Aim: To develop a method for the reliable isolation of adult human macular inner choroidal endothelial cells (ICECs) and to subsequently characterise them for their expression of a range of endothelial cell associated surface markers.
Method: Human ICECs were isolated after manual dissection of maculas from fresh human posterior segments. Following enzyme digestion to form a single cell suspension, the ICECs were isolated using anti-CD31 coated Dynabeads. The isolated cells were grown in culture and examined for typical endothelial cell morphology, surface expression of vWf, CD 31, CD 105, VEGF receptors 1 and 2, and expression of E-selectin after stimulation with TNF-α. The cells were also examined for their ability to form fenestrations and capillary-like tubes in Matrigel.
Results: The method enabled the rapid isolation of viable cells that demonstrated typical endothelial cobblestone morphology in culture. The cells stained positive for CD31, vWf, CD105, VEGF receptors 1 and 2, and E-selectin (after stimulation with TNF-α). The cells stained negative for α smooth muscle actin and fibroblast surface protein. The cells also developed fenestrations when cultured on fibronectin coated plates and formed capillary-like tubes structures when cultured on Matrigel.
Conclusions: This technique isolates cells from the human macular inner choroid that display features consistent with vascular endothelial cells. These cells could subsequently be used to further the understanding of the pathophysiological mechanisms of diseases of the inner choroid, such as choroidal neovascularisation.
- α-SMA, α smooth muscle actin
- ARMD, age related macular degeneration
- EGM, endothelial growth medium
- FITC, fluorescein isothiocyanate
- ICECs, inner choroidal endothelial cells
- MEM, minimum essential medium
- PBS, phosphate buffered saline
- RPE, retinal pigment epithelium
- TEM, transmission electron microscopy
- VEGF, vascular endothelial growth factor
- choroid
- endothelial cells
- macula
- α-SMA, α smooth muscle actin
- ARMD, age related macular degeneration
- EGM, endothelial growth medium
- FITC, fluorescein isothiocyanate
- ICECs, inner choroidal endothelial cells
- MEM, minimum essential medium
- PBS, phosphate buffered saline
- RPE, retinal pigment epithelium
- TEM, transmission electron microscopy
- VEGF, vascular endothelial growth factor
- choroid
- endothelial cells
- macula
Statistics from Altmetric.com
- α-SMA, α smooth muscle actin
- ARMD, age related macular degeneration
- EGM, endothelial growth medium
- FITC, fluorescein isothiocyanate
- ICECs, inner choroidal endothelial cells
- MEM, minimum essential medium
- PBS, phosphate buffered saline
- RPE, retinal pigment epithelium
- TEM, transmission electron microscopy
- VEGF, vascular endothelial growth factor
- choroid
- endothelial cells
- macula
- α-SMA, α smooth muscle actin
- ARMD, age related macular degeneration
- EGM, endothelial growth medium
- FITC, fluorescein isothiocyanate
- ICECs, inner choroidal endothelial cells
- MEM, minimum essential medium
- PBS, phosphate buffered saline
- RPE, retinal pigment epithelium
- TEM, transmission electron microscopy
- VEGF, vascular endothelial growth factor
- choroid
- endothelial cells
- macula
The neovascular or “wet” form of age related macular degeneration (ARMD) is characterised by the growth of new blood vessels from the inner choroid (including the choriocapillaris).1–3 While the initiating event causing wet ARMD is unknown, it is widely acknowledged that the microvascular endothelial cell plays a pivotal part in the disease process.
It is widely accepted however, that endothelial cells derived from different species, vascular organs and from within different vascular beds within those organs display different morphological, biochemical and phenotypical heterogeneity.4,5,6,7,8,9,10,11,12,13,14,15 This suggests that the use of bovine, umbilical vein, or heterogeneous mixtures of human choroidal endothelial cells to study the pathophysiological mechanisms of ARMD16–19 may not be truly representative.
To understand the angiogenic mechanisms important in the development of ARMD, it would be advantageous to study the behaviour of endothelial cells derived from the site where the disease occurs—that is, within the inner choroid of the human macula. In this study, we describe a method that allows the reliable isolation and culture of human macular inner choroidal microvascular endothelial cells (ICECs).
MATERIALS AND METHODS
Isolation of macular inner choroidal endothelial cells
Human posterior segments free of any known ocular disease were obtained from UK Transplant. The research had the approval of the local research ethics committee (Nottingham 2 LREC). After the iris had been removed, an 8 mm biopsy punch was positioned over the macula and pressure was applied, sufficient to penetrate the full thickness of the sclera. The macular sample was removed and transferred to a petri dish where the retina was discarded and retinal pigment epithelium (RPE) removed by gentle brushing with a sterile spatula and irrigation with sterile phosphate buffered saline (PBS). The choroid was then gently teased from the attached sclera and turned upside down. With the use of a dissecting microscope, the outer choroidal vessels along with adherent pigmented fibrous tissue were peeled off, leaving the relatively non-pigmented inner choroid and choriocapillaris/Bruch’s membrane complex. The tissue was cut into 1 mm pieces and washed three times in minimum essential medium (MEM) (Invitrogen Ltd, Paisley, UK) containing 30 mM HEPES, 0.25 μg/ml amphotericin B, 100 μg/ml streptomycin, 50 μg/ml kanamycin, and 30 μg/ml penicillin (isolation medium). The pieces were incubated in 0.1% collagenase I (Sigma-Aldrich) in MEM for 2 hours at 37°C, with frequent agitatation. The collagenase was neutralised with MEM containing 10% fetal calf serum (Invitrogen Ltd, Paisley, UK). and the mixture filtered through sterile 40 μm and 20 μm filters (Millipore Ltd, Watford, UK). The eluate was centrifuged (75 g) and washed three times in isolation medium, and resuspended in 1 ml of PBS/0.1%BSA. The endothelial cells were then isolated as in the manufacturer’s instructions (Dynal Ltd, Wirral, UK). The Dynabeads were resuspended in endothelial growth medium (EGM2-MV with hydrocortisone omitted, Cambrex Biosciences, Wokingham, Berks, UK) and seeded onto fibronectin coated 35 mm culture dishes (Beckton Dickinson, Oxford, UK). The cells were incubated at 37°C in a humidified atmosphere of 5% CO2 and the medium changed every 2 days. Cells were passaged with 0.025% trypsin and 0.01% EDTA in sterile PBS. All reagents were from Sigma-Aldrich, Poole, Dorset, UK unless otherwise specified.
Endothelial cell characterisation
After growing the cells (passage 1) on 1% gelatin coated glass cover slips (VWR Ltd, Poole, UK) and fixing them in ice cold methanol at −20°C for 20 minutes, a standard two stage immunofluorescence technique was applied,20 using primary antibodies detailed in table 1. (E-selectin expression was examined after exposure to TNF-α (100 pg/ml) (R and D Systems, Abingdon, UK) or PBS for 4 hours before fixation.) The secondary antibody used was rabbit anti-mouse F(ab’)2 fragment, fluorescein isothiocyanate (FITC) conjugated (1:20 dilution) (Dako, Ely, Cambridgeshire, UK) in all cases except vWf, for which swine anti-rabbit F(ab’)2 fragment FITC conjugate was used (1:20 dilution) (Dako). The slides were mounted in glycerol containing 2.5% 1–4 diazabicyclo [2, 2, 2]-octane (DABCO) (Sigma-Aldrich) and observed by confocal fluorescence microscopy (Leica TCS40D, Leica, Milton Keynes, UK). Frozen sections of fresh human placenta were used as a positive control for the vascular endothelial growth factor (VEGF) receptor antibodies, while HUVEC cells were used as a positive control for all other antibodies except for anti-α smooth muscle actin (anti-α-SMA) and anti-human fibroblast, for which human retinal pigment epithelial cells and human Tenon’s capsule fibroblasts were used respectively.
Primary antibodies used in study
Transmission electron microscopy (TEM)
Isolated cells were grown to confluence on 35 mm fibronectin coated culture dishes (Becton Dickinson, Oxford, UK) The cells were fixed by immersion in 2.5% glutaraldehyde (in 0.1 M cacodylate buffer, pH 7.4) for 16–24 hours and processed for TEM as previously described.21,22
In vitro tube formation
A 1:1 mixture of chilled Matrigel (Beckton Dickinson Biosciences, Oxford, UK) and endothelial growth medium (EGM2-MV) was dispensed into pre-chilled wells of a 96 well plate. The Matrigel was allowed to solidify at 37°C for 30 minutes before isolated cells suspended in endothelial growth medium were seeded at a density of 4.8×104 per well. The wells were observed hourly for the formation of tubes. For TEM, the endothelial growth medium was removed and replaced by 2.5% glutaraldehyde (in 0.1 M cacodylate buffer, pH 7.4) for 16–24 hours at 37°C. The fixed Matrigel was then gently removed from the well and processed as described previously.21,22
RESULTS
Human posterior segments of six donors were obtained. The age of the donors (years) and the time from death to endothelial cell isolation (hours) were: 45 (52), 76 (42), 70 (43), 37 (60), 83 (60), and 56 (60).
Light microscopy after dissection showed the sample to comprise Bruch’s membrane along with adherent choriocapillary and inner choroidal vessels (fig 1A and B).

Histological section of human submacular choroid stained with toludine blue (A) before and (B) after removal of the outer choroidal vessels. (B) Shows the residual choriocapillaris and inner choroidal vessels attached to Bruch’s membrane.
After isolation, the cells from all donors grew with a cobblestone morphology typical of endothelial cells (fig 2). The isolated cells showed a positive expression for vWf (fig 3A), CD31 (fig 3B), CD105 (fig 3C), VEGF receptor 1 (fig 3D), and receptor 2 (fig 3E), E-selectin before and after stimulation with TNFα (100 pg/ml) (fig 3F and G). The cells were negative for mouse anti-rat isotype control (fig 3H), α-SMA (fig 3I) and anti-fibroblast surface protein (fig 3J). Human RPE cells and Tenon’s capsule fibroblasts showed expression of α-SMA (fig 3K) and fibroblast surface protein (fig 3L), respectively.

Phase contrast photomicrograph of confluent primary culture of ICECs demonstrating a typical cobblestone appearance (20× original magnification).
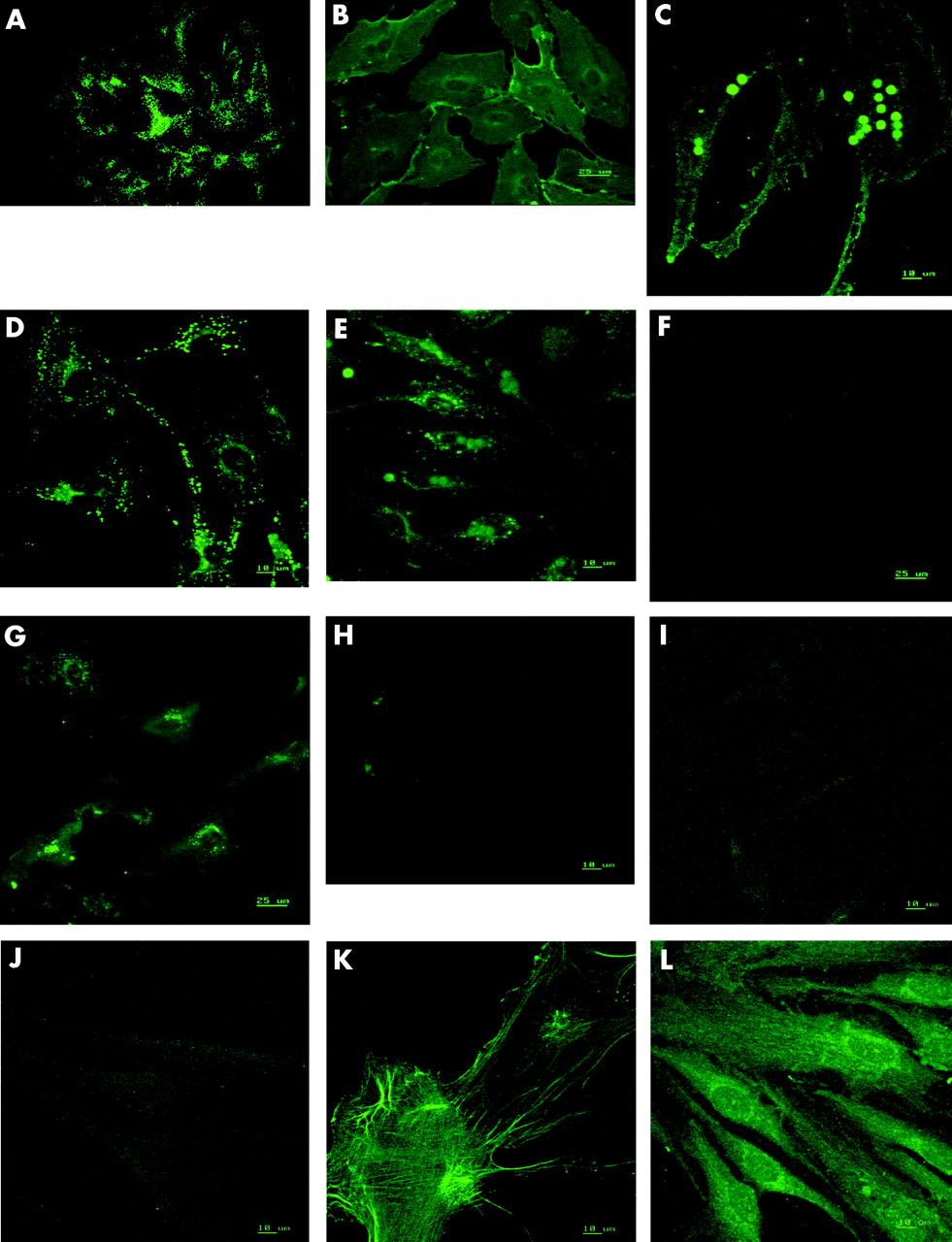
Immunofluorescent photomicrograph of human inner ICECs staining for (A) vWf, (B) CD31, (C) CD 105, (D) VEGF receptor 1, (E) VEGF receptor 2, (F) E-selectin (unstimulated), (G) E-selectin (post-TNF-α stimulation), (H)) mouse anti-rat isotype negative control, (I) α-SMA, (J) fibroblast surface protein, (K) human RPE cells stained for α-SMA, (L) human Tenon’s capsule fibroblasts stained for fibroblast surface protein. (All 63× original magnification.)
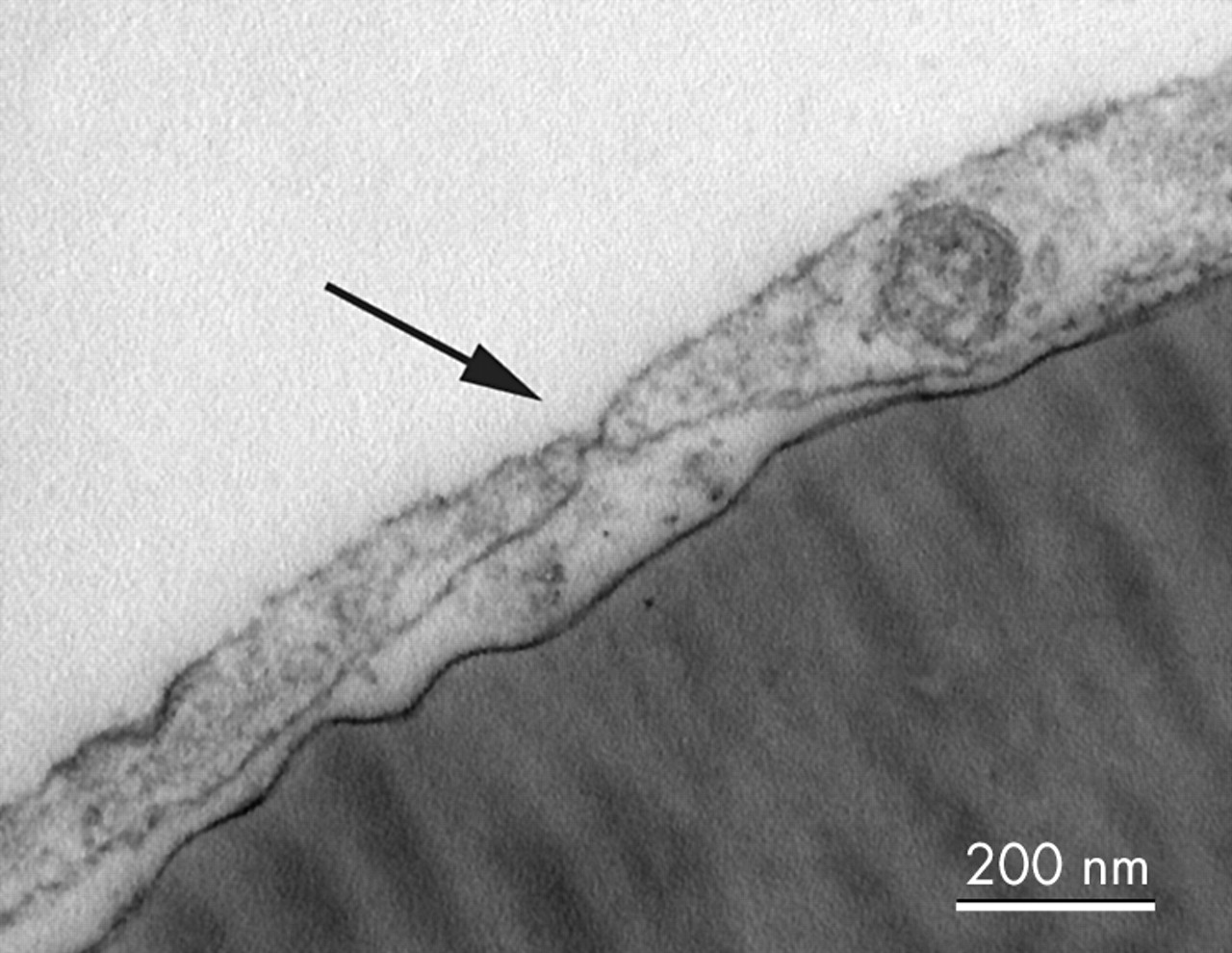
Fenestrations were found scattered throughout the cells on TEM (fig 4). Cells seeded onto Matrigel formed capillary-like tube structures within 3 hours (fig 5), and TEM showed that these structures possessed a lumen (fig 6) and each lumen was surrounded by up to three cells joined by junctional complexes.

Transmission electron micrograph of human ICECs grown on a collagen IV coated Thermanox coverslip showing a typical fenestration (arrow) (bar = 200 nm).

Phase contrast photomicrograph of human ICECs 3 hours after seeding onto Matigel showing typical capillary-like tubes (20× original magnification).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Transmission electron micrograph of a lumen of a capillary-like tube surrounded by ICECs. The cells are joined by tight junctions (higher magnification in inset) and project villi into the lumen (bar = 2 μm).
DISCUSSION
In view of the known heterogeneity of endothelial cells,4,5,6,7,8,9,10,11,12,13,14,15 it would appear logical to study endothelial cells derived from the macular inner choroidal/choriocapillaris area of humans when studying the disease mechanisms of ARMD. While methods have been described for the isolation of human choroidal endothelial cells that contain a mixture of both inner and outer choroidal endothelial cells,23,24 to date it has not been possible to reliably isolate human macular inner choroidal/choriocapillaris endothelial cells.
We describe a method to isolate human macular ICECs cells that utilise observations made around 100 years ago. In 1892, Nuel described a natural cleavage plane within the human choroid between the layers of Haller (inner choroid) and Sattler (outer choroid),25 while in 1912, Salzmann noted that dissection of the choroid was easiest by teasing apart the layers from the outside.26 With the aid of a dissecting microscope and a microdissection technique, we easily peeled the large outer choroidal vessels from the underlying inner choroid/choriocapillaris complex. The subsequent isolation of endothelial cells was achieved using anti-CD31 coated paramagnetic beads. This purification technique replaces the laborious manual method of cell sweeping or the reliance on preferential EC proliferation by using endothelial cell specific culture medium and produces isolates with a purity in excess of 99%.23,27–29
When grown in microvascular endothelial culture medium, the isolated cells formed typical cobblestone patterns consistent with their endothelial cell lineage.
The cells were found to stain strongly for CD31, a ubiquitous endothelial cell marker important in cell-cell adhesion, vWf, VEGF receptors 1 and 2, and CD105. The latter is a subunit of the TGF-β receptor found on proliferating vascular endothelial cells and has been suggested as a suitable antigen for antibody directed attack (immunotherapy) on the proliferating endothelial cells within choroidal neovascularisation complexes.30,31 The cells also showed expression of E-selectin (CD62E) after stimulation with TNF-α, a feature that is reported to be unique to endothelial cells.32 There appeared to be no contamination with RPE cells or fibroblasts.
In this study, the isolated cells also formed fenestrations when grown on fibronectin and formed capillary-like tubes when cultured in Matrigel. The lumen of these tubes was surrounded by two or more cells joined by junctional complexes, suggesting that the cells had not simply aggregated together randomly.
In summary, we have developed a method to isolate and culture human ICECs. During early passages, these cells possess the morphological characteristics of vascular endothelial cells, form fenestrations and capillary tube-like structures, and express a number of surface markers consistent with their endothelial cell origin. These cells may be useful in studying the pathophysiology and cellular mechanisms involved in choroidal neovascularisation.
Acknowledgments
We are grateful to the Kelman Foundation, USA, the British Eye Research Foundation (formerly the Iris Fund), and QMC University Hospital Trust for grant funding this project. The authors thank S Anderson and I Ward, School of Biomedical Sciences, University of Nottingham, for their assistance in conducting the confocal microscopy of the cells and I Zambrano, Eye Bank, Manchester Royal Infirmary, for the provision of the donor eye tissue.
REFERENCES
Footnotes
-
Competing interests: none declared