


Abstract
Background/Aim: We have previously reported that doxorubicin (DXR) showed much higher cytotoxicity against human oral squamous cell carcinoma cell lines compared to normal human mesenchymal normal oral cells (gingival fibroblast, periodontal ligament fibroblast, pulp cell), yielding high tumor-specificity. However, we unexpectedly found that doxorubicin showed potent cytotoxicity against human normal oral keratinocytes and primary gingival epithelial cells. In the present study, we investigated the reproducibility, underlining mechanisms and generality of this unexpected finding. Materials and Methods: Viable cell number was determined by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide method, fine cell structure by transmission electron microscopy and apoptosis induction by western blot analysis. Results: Doxorubicin induced keratinocyte toxicity, regardless of cell density and concentration of FBS in the culture medium. Doxorubicin induced apoptosis (characterized by the loss of cell surface microvilli, chromatin condensation, nuclear fragmentation and caspase-3 activation) in keratinocytes. A total of 11 anticancer drugs showed similar keratinocyte toxicity. Alkaline extract of the leaves of Sasa senanensis Rehder partially alleviated the DXR-induced keratinocyte cytotoxicity by promoting cell growth. Conclusion: The present study suggested that oral keratinocyte toxicity is a novel adverse effect of most anticancer agents.
- Cytotoxicity
- tumor selectivity
- oral carcinoma
- mesenchymal and epithelial normal oral cells
- keratinocyte toxicity
For the exploration of new anticancer drugs, experiments with tumor-bearing animals are mandatory. However, the results obtained with animals have to be carefully evaluated, since drug metabolisms are quite different between animals and humans (1, 2). The other approach is to use cell cultures that mimic whole body. We have established an in vitro assay method of anti-tumor activity, based on the relative cytotoxicity against human oral squamous cell carcinoma (OSCC) cell lines (Ca9-22, HSC-2, HSC-3, HSC-4) and human normal oral mesenchymal cells (gingival fibroblast, HGF; periodontal ligament fibroblast, HPLF; pulp cell, HPC) (3). The validity of this model for estimating the antitumor activity was obtained by our finding that anticancer drugs such as topoisomerase I inhibitors (camptothecin, SN-38) and topoisomerase II inhibitors (etoposide, teniposide), anthracyclines (doxorubicin, daunorubicin, epirubicin, mitoxantrone), mitomycin C, docetaxel, 5-FU and bacterial products (nocobactin NA-a, -b) showed one or two-orders higher tumor-specificity than hundreds of other natural products including tannins, flavonoids and other lower molecular weight polyphenols (3).
We recently found that doxorubicin showed very potent cytotoxicity against human normal oral epithelial cells (mucosa keratinocyte, HOK; primary gingival epithelial cell, HGEP) (4-6). However, the mechanism of keratinocyte toxicity is obscure. In the present study, we investigated whether keratinocyte toxicity is reproducibly observable, regardless the culturing conditions and types of anticancer drugs, and whether it is coupled with apoptosis induction. We also present our preliminary data that searched natural products for their ability to alleviate the keratinocyte toxicity.
Cytotoxic activity of anticancer drugs against human oral malignant and non-malignant cells.
Materials and Methods
Materials. The following chemicals and reagents were obtained from the indicated companies: Dulbecco's modified Eagle's medium (DMEM) from GIBCO BRL, Grand Island, NY, USA; fetal bovine serum (FBS), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), doxorubicin, daunorubicin, camptothecin, SN-38, etoposide and melphalan from Sigma-Aldrich Inc., St. Louis, MO, USA; mitomycin C from Merck KGaA, Darmstadt, Germany; docetaxel from Toronto Research Chemicals, New York, NY, USA; gefitinib from LC Laboratories®, PKC Pharmaceuticals, Inc., Woburn, MA, USA; dimethyl sulfoxide (DMSO) from Wako Pure Chem. Ind., Osaka, Japan; 5-fluorouracil (5-FU) from Kyowa, Tokyo, Japan; methotrexate from Nacalai Tesque, Inc., Kyoto, Japan. Culture plastic dishes and plates (96-well) were purchased from Becton Dickinson (Franklin Lakes, NJ, USA). Alkaline extract of the leaves of Sasa senanensis Rehder (commercial name, Sasahealth) was provided by Daiwa Biological Research Institute Co., Ltd., Kanagawa, Japan.
Cell culture. HGF, HPLF and HPC cells, established from the first premolar tooth extracted from the lower jaw of a 12-year-old girl (7), and OSCC cell lines [Ca9-22 (derived from gingival tissue; HSC-2, HSC-3, HSC-4 (derived from tongue)], purchased from Riken Cell Bank, Tsukuba, Japan, were cultured at 37°C in DMEM supplemented with 10% heat-inactivated FBS, 100 units/ml penicillin G and 100 μg/ml streptomycin sulfate under a humidified 5% CO2 atmosphere. HOK cells (purchased from COSMO BIO Co/Ltd., Tokyo, Japan) were cultured in keratinocyte growth supplement (OKGS, Cat, No. 2652). HGEP cells (purchased from CELLnTEC Advanced Cell Systems AG, Bern, Switzerland) were grown in CnT-PR medium (4-6). HOK and HGEP at 7~11 population doubling levels were used in the present study.
Assay for cytotoxic activity. Cells were inoculated at 2.5×103 cells/0.1 ml in a 96-microwell plate (Becton Dickinson Labware). After 48 h, the medium was replaced with 0.1 ml of fresh medium containing different concentrations of single test compound. Cells were incubated further for 48 h and the relative viable cell number was then determined by the MTT method (5, 6). The relative viable cell number was determined by the absorbance of the cell lysate at 540 or 562 nm, using a microplate reader (Biochromatic Labsystem, Helsinki, Finland; Sunrise Rainbow RC-R; TECAN, Männedorf, Switzerland, respectively). Control cells were treated with the same amounts of DMSO and the cell damage induced by DMSO was subtracted. CC50 was determined from the dose–response curve and the mean value of CC50 for each cell type was calculated from triplicate assays.
Western blot analysis. The cells were washed with PBS and processed for western blot analysis, as described previously (8). Antibodies against cleaved caspase-3 (Cell Signaling Technology Inc., Beverly, MD, USA), poly(ADP-ribose) polymerase (PARP) (Cell Signaling Technology Inc.) and glyceraldehyde 3-phosphate dehydrogenase (GAPDH; Trevigen, Gaithersburg, MD, USA) were used as primary antibodies. As secondary antibodies, we used α-rabbit IgG (DAKO Japan) antibodies which were conjugated with horseradish peroxidase.
Fine cell structure. Near confluent HOK cells were inoculated for 0 or 24 h with 10 μM doxorubicin. Cells detached by the cytotoxic action of doxorubicin was recovered by centrifugation. Both adherent and detached cells were then fixed and processed for the observation under the JEM-1210 transmission electron microscope (JEOL, Co., Ltd., Akishima, Tokyo, Japan) (magnification: ×5000) as described previously (4).
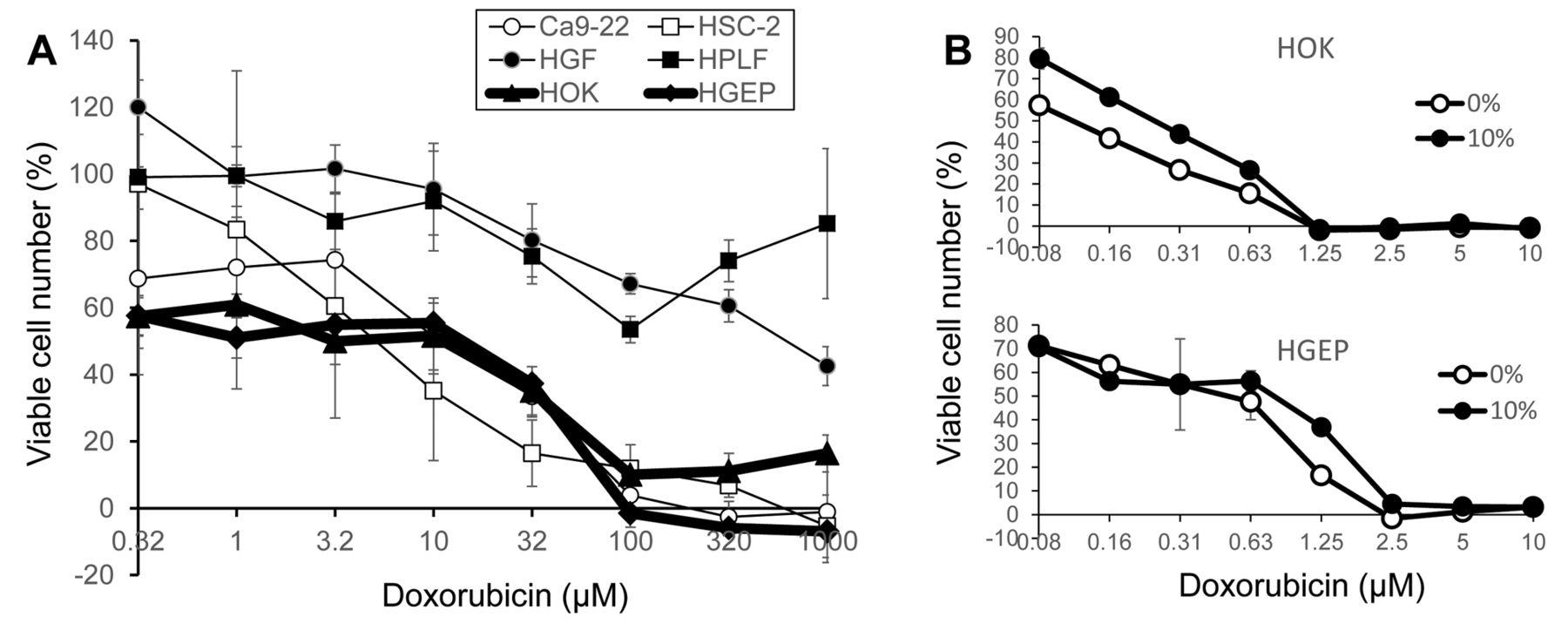
Doxorubicin induced potent cytotoxicity in human normal oral keratinocytes. (A) Comparison of doxorubicin sensitivity between human oral squamous cell carcinoma (Ca9-22, HSC-2), human normal oral mesenchymal (HGF, HPLF) and epithelial (HOK, HGEP) cells. Addition of as little as 0.32 μM doxorubicin reduced the viability of both HOK and HPEP to approximately 60% of control. The viability declined with increasing concentrations of doxorubicin. Doxorubicin at all concentrations significantly reduced the viability (p<005). (B) Effect of FBS concentration (either 0 or 10%) on the doxorubicin sensitivity. HOK (upper panel) and HGEP (lower panel) cells were treated for 48 h with the indicated concentrations of doxorubicin in specified culture medium in the presence (●) or absence (○) of 10% FBS and the viable cell number was determined by MTT method, and expressed as percent of control (with the same amount of DMSO). Each value represents mean±S.D. values of triplicate determinations. Addition of as little as 0.08 μM doxorubicin reduced the viability of both HOK and HPEP to 60~80% of control. The viability declined with increasing concentrations of doxorubicin. Doxorubicin at all concentrations significantly reduced the viability (p<005).
Tumor-selectivity index calculation. The tumor-selectivity index (TS) was calculated by the following equation: TS=mean CC50 against normal cells/mean CC50 against tumor cells, that is, B/A [HGF+HPLF+HPC (normal mesenchymal) (B) vs. Ca9-22+HSC-2+HSC-3+HSC-4 (tumor epithelial) (A)], and C/A [HOK+HGEP (normal epithelial) (C) vs. Ca9-22+HSC-2+HSC-3+HSC-4 (tumor epithelial) (A)] (Table I). A, B and C are indicated in Table I.
Statistical treatment. Experimental data are the mean±standard deviation (SD). The statistical differences between control and treated groups were evaluated by Student's t-test. A value of p<0.05 was considered to be significant.
Results
Induction of keratinocyte toxicity by doxorubicin. Doxorubicin (DXR) showed higher cytotoxicity against two human normal oral keratinocytes (HOK, HGEP), as well as two human OSCC cell lines (Ca9-22, HSC-2), compared to two human normal oral mesenchymal cells (HGF, HPLF) (Figure 1A).
There was a possibility that higher sensitivity of human oral keratinocytes may be simply due to the absence of FBS, which might have produced higher amounts of unbound doxorubicin. This possibility, however, was eliminated by our finding that the addition of FBS (final 10%) to HOK (upper panel) or HGEP (lower panel) cell culture medium only slightly reduced the cytotoxicity of doxorubicin (Figure 1B).
There was another possibility that higher sensitivity of human oral keratinocytes to doxorubicin may be due to its lower cell density, which may have keratinocytes exposed to higher concentrations of doxorubicin per a given number of cells. To test this possibility, OSCC cell lines (Ca9-22, HSC-2), normal oral epithelial (HOK) and mesenchymal (HGF, HPLF) cells were inoculated at very low (LL), low (L), high (H) or very high (HH) cell density, and then treated with increasing concentrations of doxorubicin (Figure 2A-E). When the CC50 values thus obtained were plotted as a function of cell density (absorbance at 540 nm), three different groups of cell populations were separated with each other (Figure 2F). Both keratinocytes and OSCC cell lines were more sensitive to mesenchymal cells, regardless of the cell density, eliminating this possibility.
Induction of apoptosis in keratinocyte by doxorubicin. When HOK cells were incubated for 24 h with 10 μM doxorubicin, some part of the cell monolayer was detached from the culture dish. Observation under TEM demonstrated that the detached cells showed apoptotic morphology (i.e. loss of cell surface microvilli, chromatin condensation, nuclear fragmentation) while attached cells preserved the autophagic morphology (i.e., multivascular body and lamella body) (Figure 3A). Western blot analysis demonstrated that treatment of HOK cells for 24 h with doxorubicin (2 μM) induced the cleavage of PARP and caspase-3, suggesting the activation of caspase-3 (Figure 3B). These data strongly suggest that keratinocyte toxicity by doxorubicin is coupled with induction of apoptosis.

Keratinocyte toxicity induced by doxorubicin did not depend on cell density. Adherent Ca9-22 (A), HSC-2 (B), HOK (C), HGF (D) and HPLF (E) cells were inoculated at very low (LL), low (L), high (H) or very high (HH) cell density, incubated for 48 h with the indicated concentrations of doxorubicin, and then the relative viable cell number (absorbance at 540 m) was determined by the MTT method to calculate the 50% cytotoxic concentration (CC50). Each value represents mean±S.D. of triplicate determinations. Insert (F): CC50 was plotted as a function of cell density (absorbance at 540 nm) of control cells at the time of cell harvest.
Induction of keratinocyte toxicity by many anticancer drugs. We next investigated whether the keratinocyte toxicity is specific to doxorubicin, or a general phenomenon observed in all of anticancer drugs. To discriminate these possibilities, four epithelial OSCC cell lines (CA9-22, HSC-2, HSC-3, HSC-4), three normal mesenchymal cells (HGF, HPLF, HPC) and normal epithelial keratinocytes (HOK, HGEP) were treated with various concentrations of topoisomerase I inhibitors (camptothecin, SN-38, an active metabolite of irinotecan), topoisomerase II inhibitor (etoposide), antitumor antibiotics (doxorubicin, daunorubicin, mitomycin C), antimetabolites (methotrexate, 5-fluorouracil), antimicrotubule (docetaxel), alkylating agent (melphalan) or tyrosine kinase inhibitor (gefitinib) to determine the CC50 values. All these anticancer drugs showed excellent tumor-specificity (B/A) (TS=4~2708) when normal mesenchymal cells (B) were compared to epithelial OSCC cells (A). On the other hand, they showed much lower tumor-specificity (C/A) (TS=0.1~212) when normal keratinocytes (C) were compared to epithelial OSCC cells (A) (Table I). These data demonstrate that all 11 anticancer drugs showed potent keratinocyte toxicity.

Doxorubicin induced apoptosis in HOK human normal oral keratinocyte. (A) Transmission electron microscope (TEM) analysis. HOK cells were treated for 0 (a) or 24 (b, c) h with 10 μM doxorubicin and fine cell structure of adherent (a, b) and detached cells (c) were observed under TEM. Bars indicate 1 μm (a, b) and 2 μm (c). (B) Western blot analysis. HOK cells were treated for 24 h with either vehicle (DMSO) (0.2%) (lane 1), doxorubicin (0.4 and 2 μM) (lane 2 and 3) or actinomycin D (1 μM) (lane 4). Detached and attached cells were combined for western blot analysis.
Discussion
The present system demonstrated that doxorubicin induced very potent toxicity against human oral keratinocytes, regardless of culture conditions (presence or absence of FBS, cell density), accompanied by apoptosis induction (Figure 3). It should be noted that during detachment from the culture plate, cells were committed to apoptosis (Figure 3A-C) while attached cells still preserved the autophagic phenotype (Figure 3A-B). We have previously reported that (E)-3-(4-hydroxystyryl)-6-methoxy-4H-chromen-4-one, having high tumor-specificity (TS=69) and lower keratinocyte toxicity, induced apoptosis in HSC-2 cells after autophagy was suppressed (4). These data suggest that suppression of autophagy triggers apoptosis induction, in accordance with the generally accepted role of autophagy in maintaining the long-term survival of disseminated residual cells (9, 10).
The present study demonstrated that doxorubicin induced apoptosis also in human normal oral keratinocytes, like that has previously been published in various cancer cells (11-13). This indicates the induction of apoptosis itself does not guarantee the anti-tumor potential. A simple search in pubmed with two key words, flavonoids and apoptosis, got over nearly 8000 publications, even though most of flavonoids showed lower levels of tumor-specificity (TS values=1.3~3.2) (3). Taken together, tumor-specificity, rather than apoptosis-inducing activity, should be used for the evaluation of antitumor activity.
We found that most of anticancer drugs showed keratinocyte toxicity (Table I). This points out that keratinocyte toxicity is new adverse effect of anticancer agents, in addition to cardiomyopathy (11). Further study is required to elucidate why human oral keratinocytes are more sensitive to anticancer drugs compared to human mesenchymal cells.
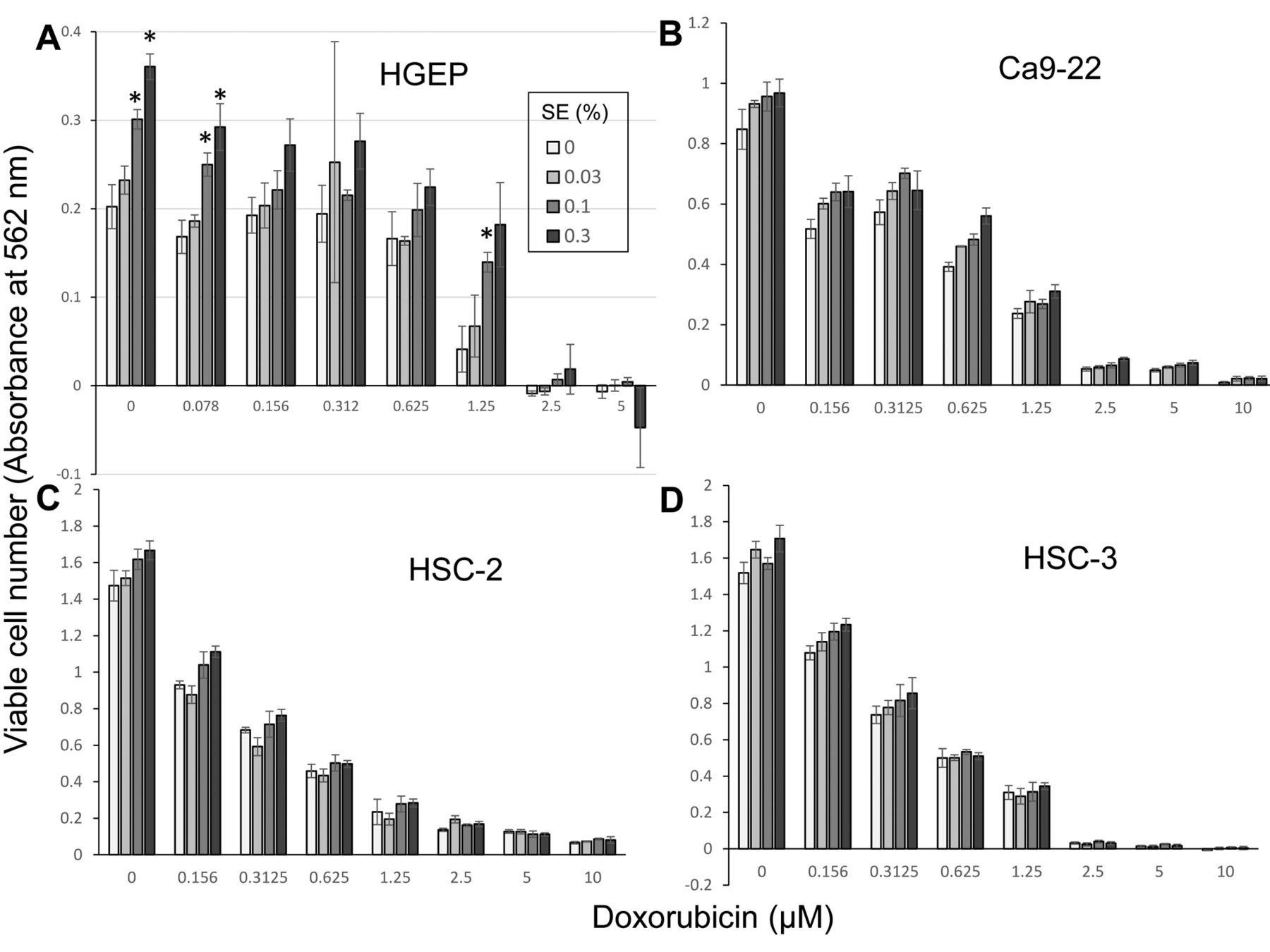
It, thus, became very important to search for natural products that can alleviate the doxorubicin-induced keratinocyte toxicity. We found that alkaline extract of the leaves of Sasa senanensis Rehder (SE), OTC drug, having prominent antiviral (14) and anti-UV activity (15), significantly alleviated the DXR-induced cytotoxicity against HGEP, by its growth-promoting activity (Figure 4A). On the other hand, the growth promoting effect of SE against OSCC cell lines, Ca9-22) (B), HSC-2 (C) and HSC-3 (D) was significantly less (Figure 4). Further study is required to identify the active ingredient of SE that promote the growth of HGEP.
In conclusion, the present paper suggests that keratinocyte toxicity may be the new adverse effect of most of anticancer agents.

Protective effect of alkaline extract from the leaves of Sasa senanensis Rehder (SE) on doxorubicin cytotoxicity. HGEP (A), Ca9-22 (B), HSC-2 (C) or HSC-3 (D) cells were incubated for 48 h with the indicated concentrations of doxorubicin in the presence of 0, 0.03, 0.1 or 0.3% SE, and the viable cell number (absorbance at 562 mm) was determined by the MTT method. Each value represents mean±S.D. of triplicate determinations. Asterisks indicate statistical significance at p<0.05.
Acknowledgements
This work was partially supported by KAKENHI from the Japan Society for the Promotion of Science (JSPS) (16K11519).
Footnotes
This article is freely accessible online.
Conflicts of Interest
The first Author (HS) was supported by the Daiwa Biological Research Institute Co., Ltd., Kanagawa, Japan. The authors wish to confirm that such financial support has not influenced the outcome or the experimental data.
- Received December 14, 2016.
- Revision received January 30, 2017.
- Accepted January 31, 2017.
- Copyright© 2017, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Potentiation of Anticancer Activity of G2/M Blockers by Mild Hyperthermia
- Prominent Anti-UVC Activity of Lignin Degradation Products
- Inhibition of Neurotoxicity/Anticancer Activity of Bortezomib by Caffeic Acid and Chlorogenic Acid
- Antitumor Effects and Tumor-specificity of Guaiazulene-3-Carboxylate Derivatives Against Oral Squamous Cell Carcinoma In Vitro
- In Vitro Assessment of Antitumor Potential and Combination Effect of Classical and Molecular-targeted Anticancer Drugs
- Quantitative Structure-Cytotoxicity Relationship of 2-Arylazolylchromones and 2-Triazolylchromones
- Quantitative Structure-Cytotoxicity Relationship of Azulene Amide Derivatives
- Quantitative Structure-Cytotoxicity Relationship of Pyrano[4,3-b]chromones
- Quantitative Structure-Cytotoxicity Relationship of 3-(N-Cyclicamino)chromone Derivatives
- Partial Protection of Paclitaxel-induced Neurotoxicity by Antioxidants
- Quantitative Structure-Cytotoxicity Relationship of 2-(N-cyclicamino)chromone Derivatives
- Quantitative Structure-Cytotoxicity Relationship of Furo[2,3-b]chromones
- In Vitro Anti-tumor Activity of Azulene Amide Derivatives
- In Vitro Antitumor Activity of Alkylaminoguaiazulenes
- Protection of Differentiating Neuronal Cells from Amyloid {beta} Peptide-induced Injury by Alkaline Extract of Leaves of Sasa senanensis Rehder
- Quantitative Structure-Cytotoxicity Relationship of Cinnamic Acid Phenetyl Esters
- Quantitative Structure-Cytotoxicity Relationship of 2-Azolylchromones
- Quantitative Structure-Cytotoxicity Relationship of Aurones
- Quantitative Structure-Cytotoxicity Relationship of Newly Synthesized Piperic Acid Esters
- Search for New Type of Anticancer Drugs with High Tumor Specificity and Less Keratinocyte Toxicity