


Abstract
Background/Aim: We previously showed that the use of autophagy inhibitors in combination with chemotherapy can enhance anticancer effects in sarcoma cell lines. In this study, we investigated the combined effect of the autophagy inhibitor chloroquine and the mTOR inhibitor rapamycin on MG63 osteosarcoma cells. Materials and Methods: Effects of chloroquine and/or rapamycin on cell proliferation were assessed by WST-1 assays. Effects of chloroquine and/or rapamycin on the mTOR pathway components, autophagy, and apoptosis were investigated by western blot, flow cytometry, and fluorescence microscopy using immunocytochemical staining of LC3 and Annexin V-FITC/propidium iodide. Results: Rapamycin suppressed cell growth and inhibited the mTOR pathway. Rapamycin promoted autophagy by blocking the mTOR pathway, and chloroquine enhanced apoptosis by blocking autophagy. Conclusion: Chloroquine enhances the effects of rapamycin in inducing apoptosis via autophagy inhibition in MG63 cells. Thus, the combined therapy of chloroquine and rapamycin may be a potent treatment for osteosarcoma.
Osteosarcoma, the most common type of malignant bone neoplasm, shows high morbidity and mortality (1). Despite recent therapeutic advances, recurrence and metastasis continue to pose a challenge for osteosarcoma treatment (2). Recently, several molecular targeted drug therapies have been developed for the treatment of malignant neoplasms (3). These therapies exhibit potent anti-tumour effects by targeting growth factor receptors or intracellular signalling proteins that are associated with tumour growth and metastasis (4).
Mammalian target of rapamycin (mTOR) plays a central role in regulating various cellular functions, such as survival and proliferation (5). Cell growth factors and a rich nutritional environment can activate the PI3K/Akt/mTOR signalling pathway (6), thereby promoting cell growth and proliferation. Recent studies have reported the abnormal activation of mTOR in various cancers, including bone sarcomas (7). Thus, targeting mTOR, which leads to inhibition of these downstream effectors, may be an effective treatment for osteosarcoma. Some studies have suggested that mTOR inhibitors exhibit anti-tumour effects in cancer cells (8, 9). In fact, we have previously shown the anticancer activity of mTOR inhibitor rapamycin in bone and soft tissue sarcomas (10, 11).
Autophagy is a critical cellular process that regulates the degradation of proteins and organelles to provide cellular energy (12). It also serves as a temporary defence mechanism to protect cells from hostile environments (13). Furthermore, autophagy inhibitors can activate apoptosis induced by anticancer drugs and as such improve their effectiveness (14). Chloroquine is an anti-malarial drug (15) that has been used as an autophagy inhibitor. Chloroquine inhibits the fusion of autophagosomes with lysosomes and subsequently blocks their lysosomal degradation, thereby increasing the accumulation of autophagosomes (16). Recent studies have demonstrated the use of chloroquine in combination with anticancer drugs for the treatment of malignant tumours (17, 18). However, combined treatment using chloroquine and rapamycin in osteosarcoma cells has not yet been reported.
In this study, we aimed to analyze the effects of the mTOR inhibitor rapamycin on MG63 osteosarcoma cell growth and autophagy induction. Moreover, we investigated the effects of the combined treatment of rapamycin and chloroquine on apoptosis and autophagy in MG63 osteosarcoma cells. This is the first report on the effects of chloroquine and rapamycin on MG63 osteosarcoma cells.
Materials and Methods
Chemical reagents, cell lines, and cell culture. MG63 cell line, an osteogenic sarcoma cell line, was purchased from the RIKEN BRC through the National Bio-Resource Project of the MEXT (Ibaraki, Japan). Cells were grown in Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich, St, Louis, MO, USA) with 10% foetal bovine serum (FBS; Sigma-Aldrich) and 100 U/ml penicillin/streptomycin solution (Sigma-Aldrich), and were cultured at 37°C with 5% CO2. Chloroquine was obtained from Sigma-Aldrich and rapamycin (CCI-779) was obtained from Calbiochem (San Diego, CA, USA). To examine the effects of rapamycin, cells were treated with 0, 0.4, 2, 10, or 50 μM rapamycin. To examine the effects of chloroquine and/or rapamycin, cells were divided into four groups: i) control (no inhibitor), ii) CQ (20 μM chloroquine), iii) RAP (20 μM rapamycin), and iv) CQ+RAP (20 μM chloroquine + 20 μM rapamycin).
Cell proliferation assay. The cell proliferation reagent WST-1 was obtained from Sigma Aldrich. Cells were seeded in 96-well culture plates at a density of approximately 1×104 cells/well and were cultured for 48 h. To examine the effects of rapamycin, the medium was subsequently replaced with a fresh medium containing, for each case, different concentrations of rapamycin, and cells were cultured for 24 or 48 hours. To examine the effects of chloroquine and/or rapamycin, the cells were pre-treated with or without chloroquine for 12 hours, they were then washed twice with phosphate-buffered saline (PBS), and they were finally treated with or without rapamycin for 24 h. After 2 h of incubation with WST-1, the optical density at 420 nm was recorded using a microplate reader (Molecular Devices, Sunnyvale, CA, USA). Percentages of cell viability were calculated using the following formula: Cell viability (percentage of control)=(sample OD - blank OD)/(control OD - blank OD) ×100%. We performed at least three independent tests.
Western blotting. Cells were seeded at approximately 6×105 cells/well in 6-well plates. After 48 h of culture, cells were treated for 24 h to examine the effects of rapamycin, of chloroquine, and their combination. Following treatment, the cells were lysed in lysis buffer (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, 1 mM Na2EDTA1 mM EGTA, 1% Triton, Cell Signalling Technology, Beverly, MA, USA) on ice for 20 min. The protein samples were separated by 10% sodium dodecyl sulphate polyacrylamide gel electrophoresis (Wako, Tokyo, Japan) and under reducing conditions. The proteins were electrophoretically transferred to nitrocellulose membranes (GE Healthcare Bio-Sciences, Piscataway, NJ, USA) and sequentially incubated with primary and anti-rabbit secondary antibodies in iBind solution (iBind Western System, Life Technologies, Carlsbad, CA, USA) for 2.5 hours. Primary antibodies for anti-4E-BP1, phospho-4E-BP1, p70S6k, phospho-p70S6k, cleaved-caspase 3, cleaved-caspase 9, cleaved-PARP, and a-tubulin were purchased from Cell Signaling Technology. Primary antibodies for microtubule-associated protein light-chain3 (LC-3), anti- Atg12-Atg5 complex, and p62/SQSTM1 were purchased from MBL Co. (Nagoya, Japan). All antibodies were used at a dilution of 1:1,000. Labelled proteins were determined using a Novex® AP Chemiluminescent Detection Kit (Life Technologies) and a LAS-1000 plus image analyser (Fujifilm Co., Tokyo, Japan).
Detection of apoptosis using flow cytometry. Cells were plated at 6×105 cells/well in 6-well plates and were cultured for 48 hours. To examine the effects of chloroquine and/or rapamycin, cells were treated for 24 hours and were stained with Annexin V-FITC and propidium iodide (PI) using the Annexin V-FITC kit (Beckman Coulter, Inc., Brea, CA, USA) for 10 minutes. Apoptosis was detected using an FC500 flow cytometer (Beckman Coulter). Data are representative of three independent experiments.
Immunocytochemical staining for LC3. Cells were plated at 1×106 cells/well in 6-well plates on 25-mm circular coverslips and were cultured for 24 h. To examine the effects of chloroquine and/or rapamycin, cells were treated for 24 h and were fixed in 4% paraformaldehyde in phosphate buffer for 30 min. Then, the cells were sequentially incubated with anti-LC3 antibody (MBL) for 1 h and with anti-IgG secondary antibody (Alexa Fluor® 488, code no. A11008; Invitrogen, Carlsbad, CA, USA) for 30 minutes. Samples were then examined under an epifluorescence microscope (FSX100, Olympus Optical Co., Ltd., Tokyo, Japan) with a 50× objective lens.
Apoptosis detection by morphological analysis. Cells were plated, cultured, and treated as described above. Then, the cells were stained with Annexin V-FITC, PI, and Hoechst 33342 using the Promokine Apoptotic/Necrotic/Healthy Cells detection kit (PromoCell GmbH, Heidelberg, Germany), according to the manufacturer's instructions, and were incubated for 15 min. Stained cells were viewed with an epifluorescence microscope (FSX100, Olympus Optical Co.).
Statistical analysis. We performed statistical analyses using the GraphPad Prism 5 software (GraphPad, San Diego, CA, USA). Proliferation assay results were analysed with one- or two-way analysis of variance, followed by a post-hoc analysis. We considered p<0.05 as statistically significant.
Results
Chloroquine enhances rapamycin-mediated suppression of MG63 cell proliferation. Rapamycin inhibited MG63 cell proliferation in a dose and time-dependent manner (Figure 1A). Furthermore, compared to the RAP group, the CQ+RAP group showed a significantly higher inhibition of MG63 cell proliferation after 24 h treatment (p<0.05, Figure 1B).
Western blot analysis demonstrated that chloroquine inhibits rapamycin-induced autophagy and enhances rapamycin-induced apoptosis in MG63 cells. Rapamycin induced a dose-dependent decrease in the phosphorylation of 4E-BP1 and p70S6k, the downstream molecules of the Akt/mTOR pathway, in MG63 cells. Furthermore, rapamycin increased the levels of LC-3II and the Atg12-Atg5 complex, which are autophagy-related proteins, and decreased the level of p62/SQSTM1 in a dose-dependent manner (Figure 2A). p62/SQSTM1 protein is observed in inclusion bodies containing ubiquitinated protein aggregates, which are degraded by autophagy (19). Notably, the expression levels of LC-3II and p62/SQSTM1 in the CQ+RAP group were higher than those in the RAP group. In addition, the levels of apoptosis-related proteins (cleaved caspase-3, cleaved caspase-9, and cleaved PARP) were increased in the RAP group and further markedly increased in the CQ+RAP group, compared to the CQ group (Figure 2B).

Results of the WST-1 cell proliferation assay in MG63 cells. (A) Rapamycin inhibited MG63 cell proliferation in a dose-dependent and time-dependent manner (*p<0.05). (B) MG63 cell proliferation in the CQ+RAP group was lower than that in the RAP group (*p<0.05).

Western blotting analysis. (A) Rapamycin induced a dose-dependent inhibition of the mTOR pathway, and a dose-dependent increase of autophagy-related proteins. (B) The levels of LC-3II and p62/SQSTM1 in the CQ+RAP group were higher than those in the RAP group. Apoptosis-related protein levels were slightly increased in the RAP group and notably increased in the CQ+RAP group.
Flow cytometry analysis confirmed that chloroquine enhanced rapamycin-induced apoptotic cells. Annexin V-FITC-positive and PI-negative cells were considered early apoptotic cells, and Annexin V-FITC-positive and PI- positive cells were considered late apoptotic cells (Figure 3A). Apoptotic cells were significantly increased in the CQ+RAP group, compared to those in the Rap group (Figure 3B).
Immunocytochemical staining for LC3 revealed that chloroquine inhibited rapamycin-induced autophagy. LC3-positive puncta (autophagosomes) were increased in the CQ and RAP groups but were markedly increased in the CQ+RAP group (Figure 4A).
Annexin V-FITC/PI staining confirmed apoptosis induction after combined chloroquine and rapamycin treatment. Several early apoptotic cells (Annexin V-FITC+/PI−) and many late apoptotic cells (Annexin V-FITC+/PI+) were observed in the CQ+RAP group (Figure 4B). This result was consistent with that from the flow cytometry analysis.
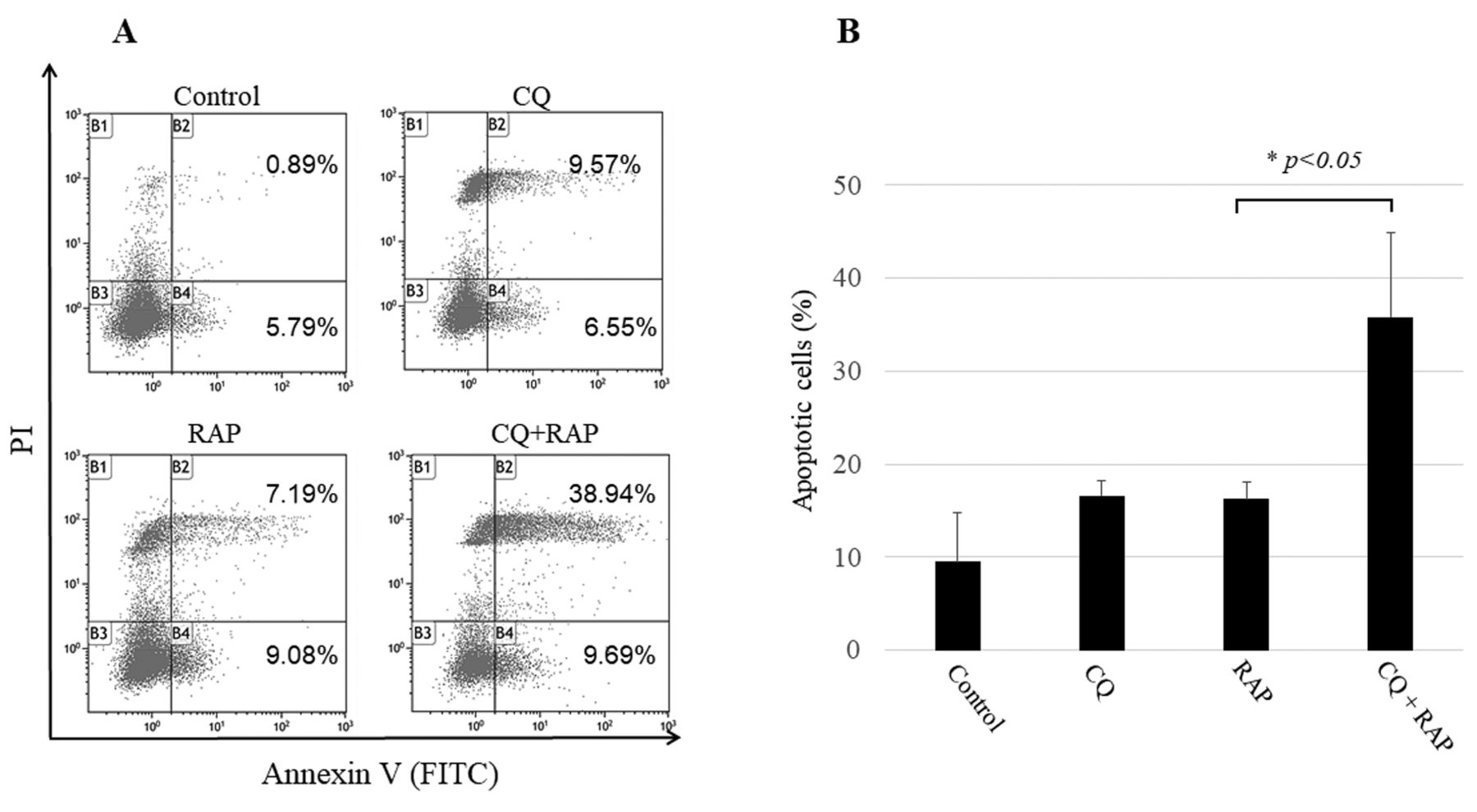
Apoptosis detection by flow cytometry using Annexin V-FITC/PI staining. (A) MG63 cells were treated with 20 μM CQ and/or 20 μM RAP for 24 h and were stained with Annexin V-FITC/PI. (B) The histogram shows a comparison of the percentages of apoptotic cells within the treatment groups. The percentages of apoptotic cells are presented as mean±SD of three experiments. The percentage of apoptotic cells in the CQ+RAP group was significantly higher than that in the other groups (*p<0.05).

Morphological analysis by fluorescence microscopy. (A) The CQ and RAP groups showed increased LC3-positive puncta (arrows: autophagosomes). However, the CQ+RAP group showed a further increase in LC3-positive puncta (B) Apoptotic cells were detected by Annexin V-FITC and PI staining assay. Annexin V-FITC (green): early apoptotic cells (open arrows); PI (red): late apoptotic cells (closed arrows); Hoechst 33342 (blue): nuclei of living cells. Several apoptotic cells can be observed in the CQ+RAP group (Scale bars=50 μm).
Discussion
Although a combination of autophagy inhibitors and anti-cancer drugs has been shown to be effective for cancer therapy, the combined effect of the autophagy inhibitor chloroquine and the mTOR inhibitor rapamycin has not been previously reported. In this study, we investigated the effects of rapamycin on MG63 osteosarcoma cell growth and autophagy induction. Moreover, we studied the effects of the combined treatment of rapamycin and chloroquine on the apoptosis and autophagy of MG63 osteosarcoma cells in vitro. We found that rapamycin inhibited cell growth and induced autophagy in MG63 cells, however, the combined rapamycin and chloroquine treatment strongly inhibited cell proliferation and increased the number of apoptotic cells autophagy inhibition.
Rapamycin was previously shown to block the mTORC1-dependent phosphorylation of 4E-BP1 and the p70S6K phosphorylation (20). In this study, we found that rapamycin inhibited cell proliferation in a dose and time-dependent manner and decreased the 4E-BP1 and p70S6K phosphorylation in a dose-dependent manner. These results indicate that rapamycin successfully inhibits the mTOR pathway in MG63 cells.
Autophagy is an important physiological process that regulates the degradation of proteins and organelles and sustains homeostasis. Autophagy plays a protective role in regulating cellular homeostasis under unfavourable conditions, such as exposure to environmental stress (21, 22). Autophagy has also been shown to play an important role in cancer cells with respect to their response to cellular stress and increased metabolic demands due to fast cell growth (23). Furthermore, cancer cells can activate autophagy as a defence mechanism to resist chemotherapy (24). Indeed, our western blot results showed that rapamycin increased the level of the autophagy markers, LC3-II and Atg5-Atg12 complex, and decreased the level of p62/SQSTM1 in MG63 cells in a dose-dependent manner.
In contrast, we found that the levels of LC-3II and p62/SQSTM1 in cells treated with chloroquine and rapamycin were both higher than those in cells treated with rapamycin alone. This was because chloroquine inhibits autophagic flux causing accumulations of autophagosomes. Similarly, immunocytochemical analysis revealed a marked increase in the number of LC3-positive puncta in cells treated with rapamycin and chloroquine, compared to the cells treated with rapamycin alone. These results may be attributed to the inhibition of the late phase of autophagy by chloroquine and the subsequent increase in autophagosome accumulation, which leads to increased LC3-II and p62/SQSTM1 levels and LC3-positive puncta. The LC3 turnover assay is a similar method, and one of the principal ones, currently used to measure autophagic flux (25). Taken together, our findings indicate that chloroquine inhibited the rapamycin-induced autophagy.
Recently, some studies have suggested that blocking autophagy can trigger apoptosis in cancer cells (14, 26). Therefore, autophagy inhibition may be a potent therapeutic approach for osteosarcoma. Chloroquine, which is being widely investigated as an autophagy inhibitor, blocks the later step of autophagy by inhibiting lysosome activity (27). Because cancer cells maintain high cell growth via autophagy-derived energy, chloroquine can sensitise cancer cells through autophagy inhibition. Recent reports have demonstrated the effective use of chloroquine as an autophagy inhibitor for cancer therapy. A combination treatment of androgen deprivation and chloroquine appear to exhibit a synergistic apoptotic effect in prostate cancer cells (28). Furthermore, chloroquine was reported to potentiate the anti-cancer effect of fluorouracil in colon cancer cells (29). In our study, cell proliferation was strongly inhibited after treatment with rapamycin and chloroquine, indicating that chloroquine improves the rapamycin-induced suppression of MG63 cell growth. Furthermore, western blot analysis revealed increased caspase-3 and PARP levels in cells treated with rapamycin and chloroquine both, compared to cells treated with rapamycin alone. In addition, we observed a large number of apoptotic cells treated with both rapamycin and chloroquine using flow cytometry analysis and numerous Annexin V-FITC-positive cells using the Annexin V/PI staining assay. These results indicate that apoptosis induction was considerably higher when the combined treatment with rapamycin and chloroquine was used, compared to the effect of the individual drug treatments or in the control cells. Thus, our study showed that autophagy may be activated as a defence mechanism of MG63 cells in response to rapamycin and that inhibiting autophagy by co-treatment with chloroquine promoted the activation of apoptotic cell death.
In conclusion, we demonstrated that rapamycin promotes autophagy in MG63 cells by blocking the mTOR signalling pathway and chloroquine enhances apoptotic cell death by blocking autophagy. A limitation of this study is that we conducted experiments with only one cell line, and so we plan to perform similar experiments with other sarcoma cell lines in the future. Taken together, this study shows that autophagy may be a protective mechanism adopted by MG63 cells in response to rapamycin and that combination therapy of chloroquine and rapamycin may be a potent treatment for osteosarcoma.
Footnotes
Conflicts of Interest
The Authors declare no conflicts of interest.
- Received December 27, 2018.
- Revision received January 10, 2019.
- Accepted January 17, 2019.
- Copyright© 2019, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Targeting Autophagy With the Synergistic Combination of Chloroquine and Rapamycin as a Novel Effective Treatment for Well-differentiated Liposarcoma
- Chloroquine Combined With Rapamycin Arrests Tumor Growth in a Patient-derived Orthotopic Xenograft (PDOX) Mouse Model of Dedifferentiated Liposarcoma
- Rapalink-1 and Hydroxychloroquine Exhibit an Additive Effect in Undifferentiated Pleomorphic Sarcoma by Inducing Apoptosis