
Casticin Inhibits A375.S2 Human Melanoma Cell Migration/Invasion through Downregulating NF-κB and Matrix Metalloproteinase-2 and -1

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
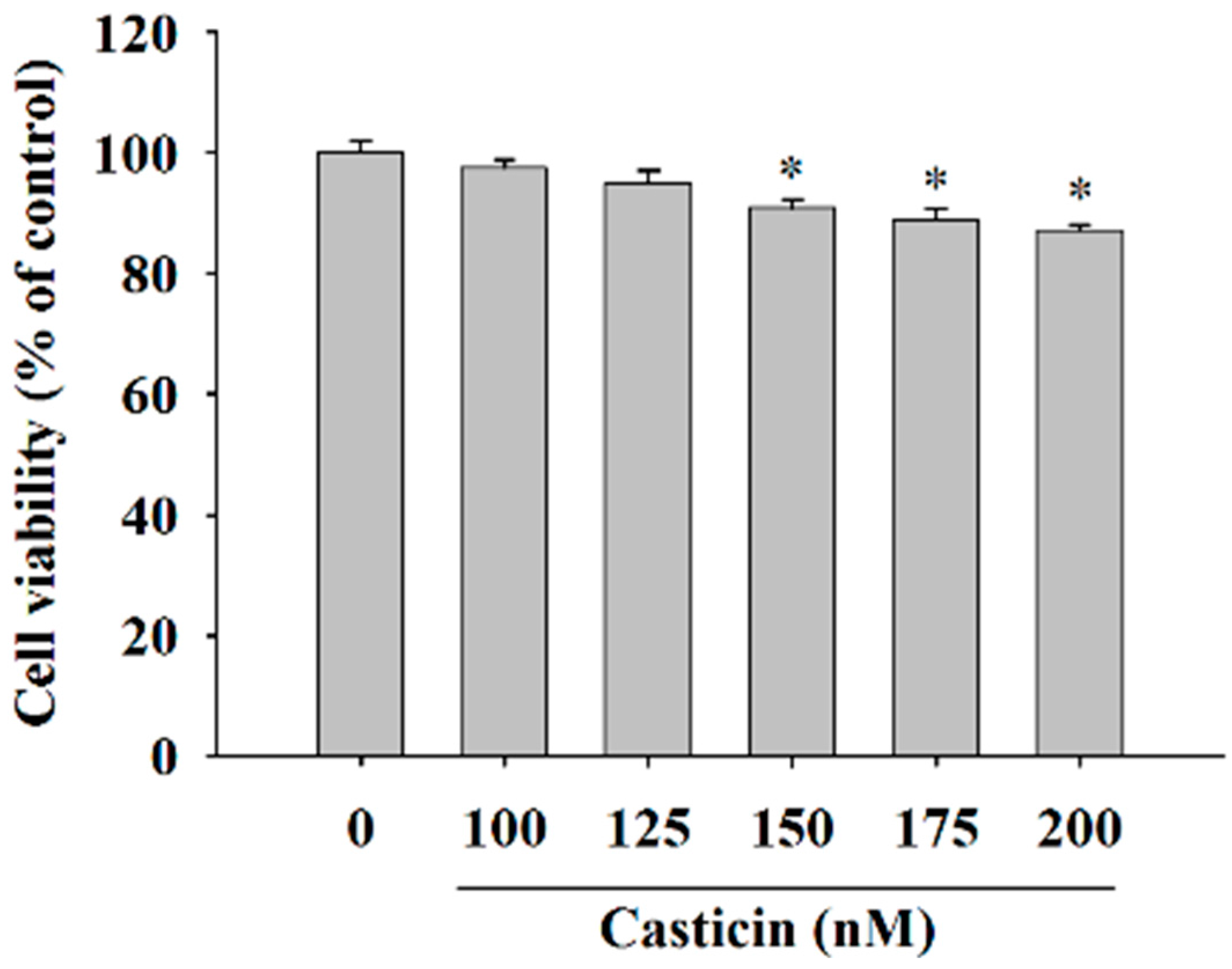
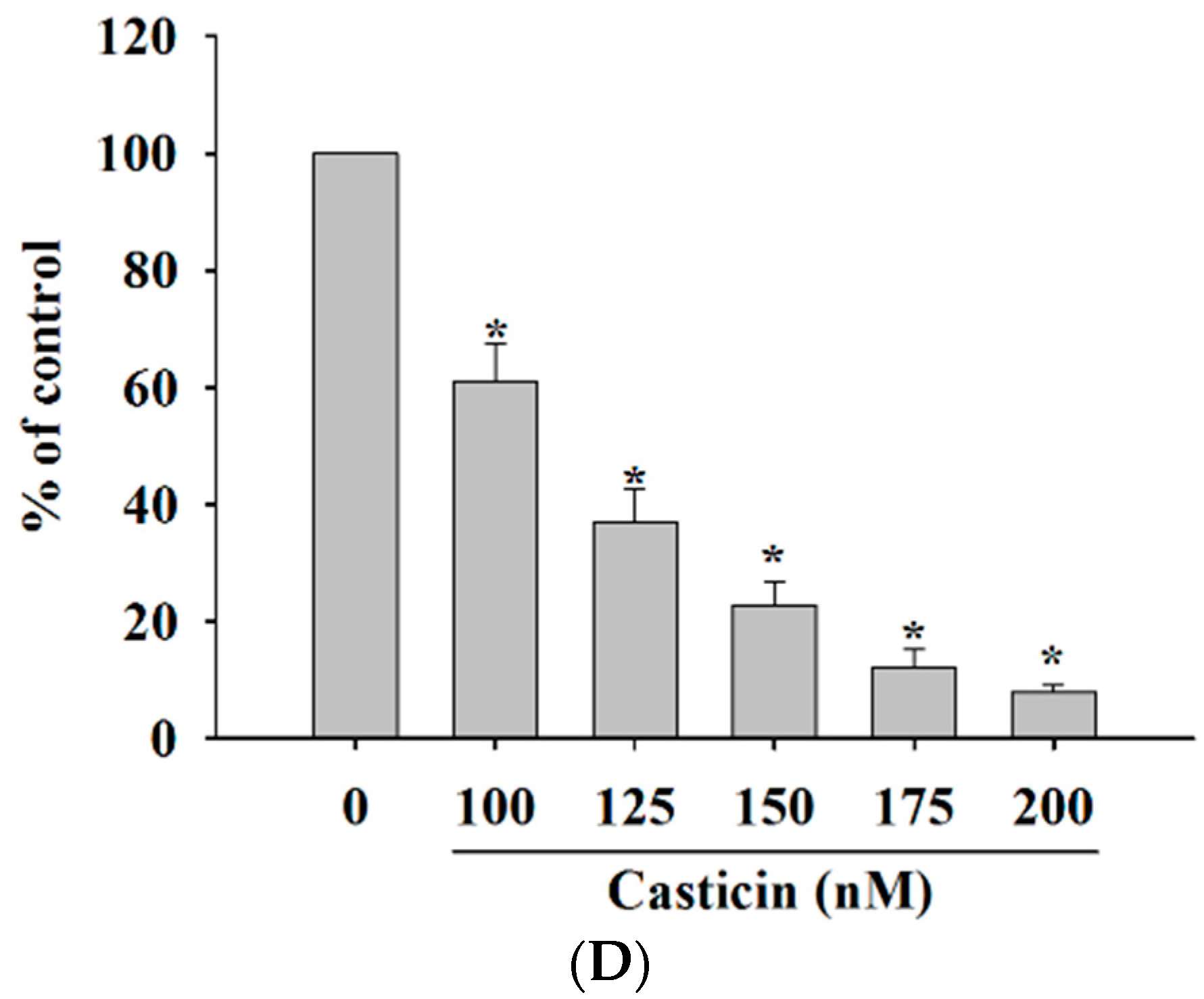
2.1. Casticin Decreased the Cell Viability of A375.S2 Cells
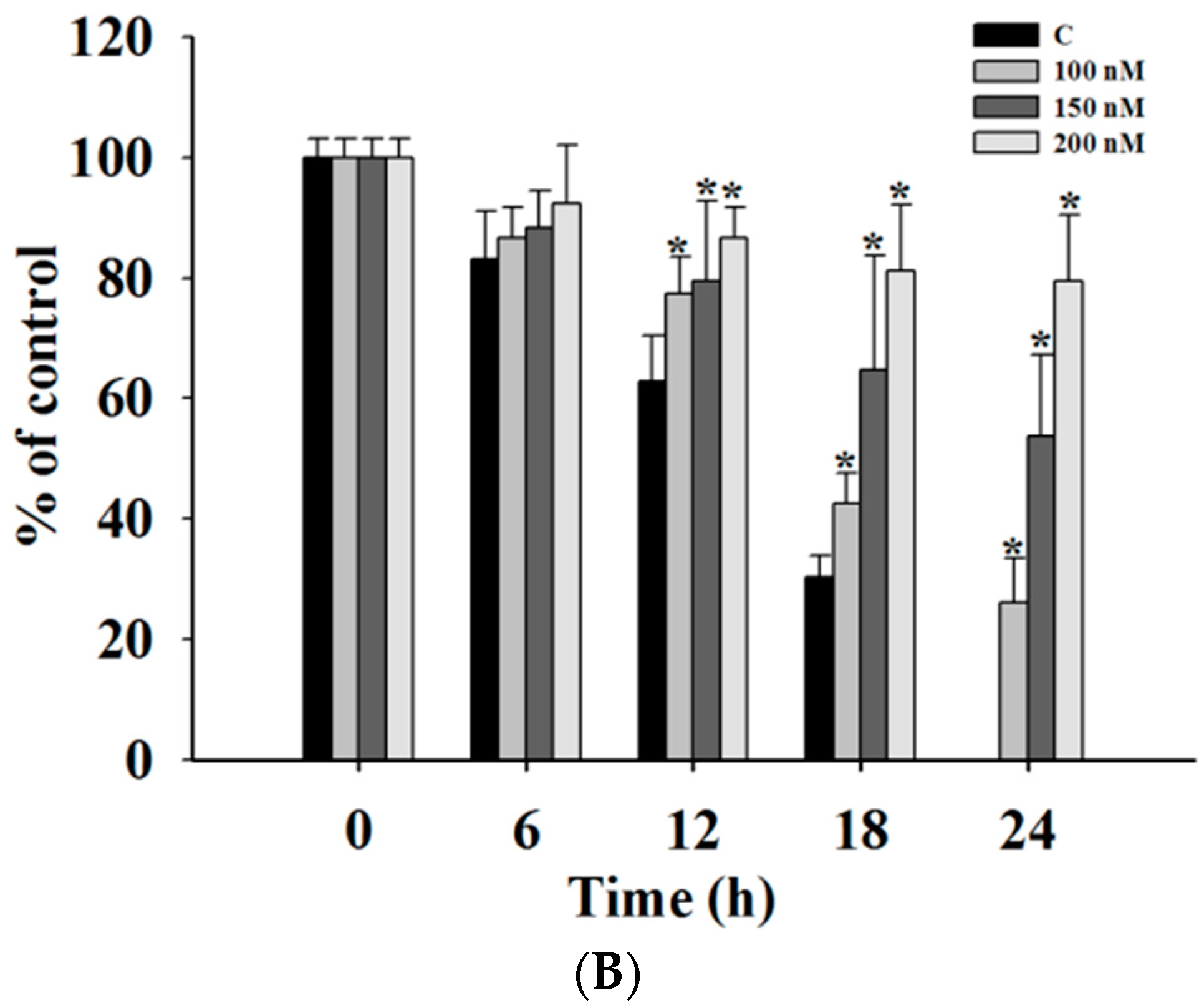
2.2. Casticin Inhibits the Motility of A375.S2 Cells
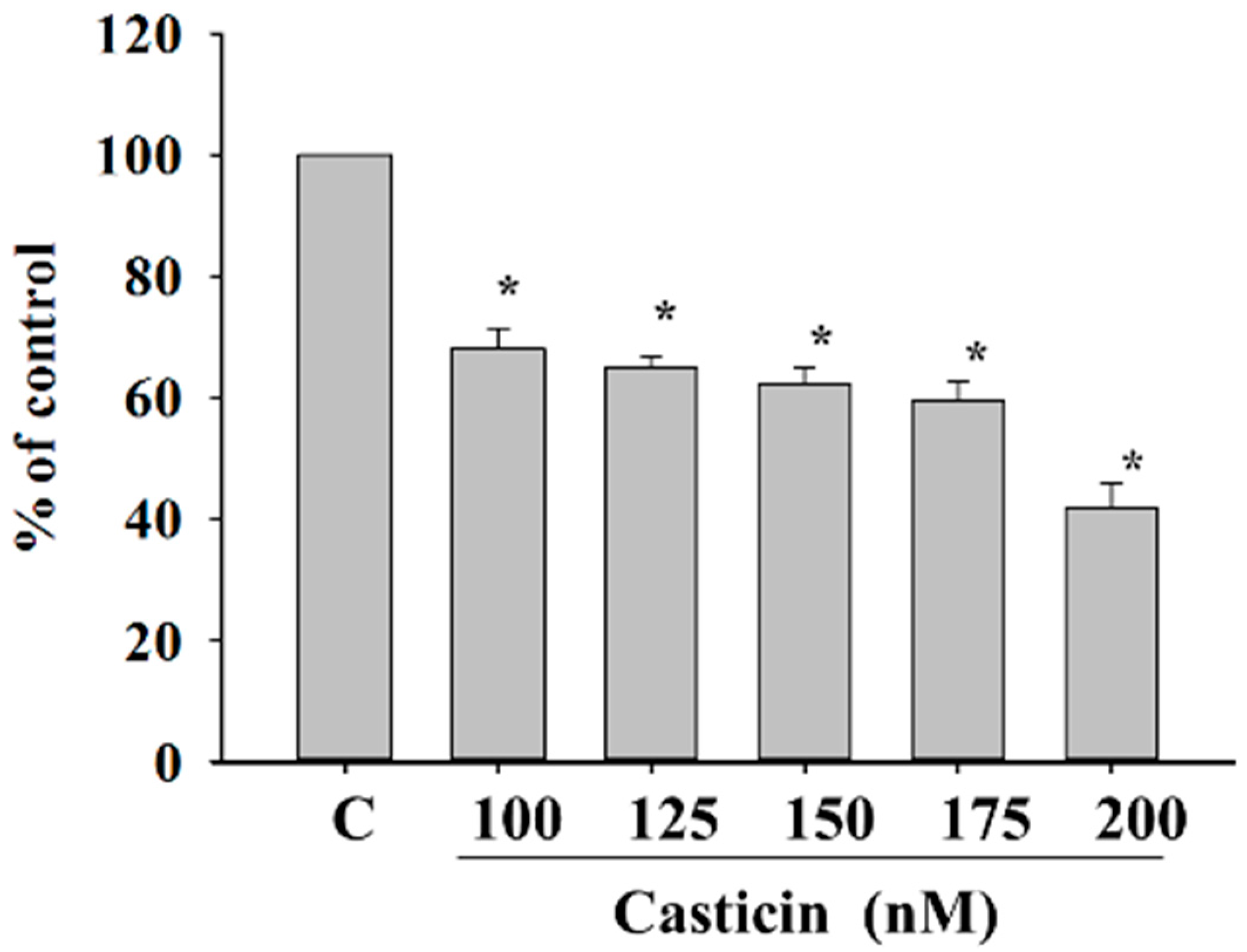
2.3. Casticin Inhibit Adhesion of A375.S2 Cells
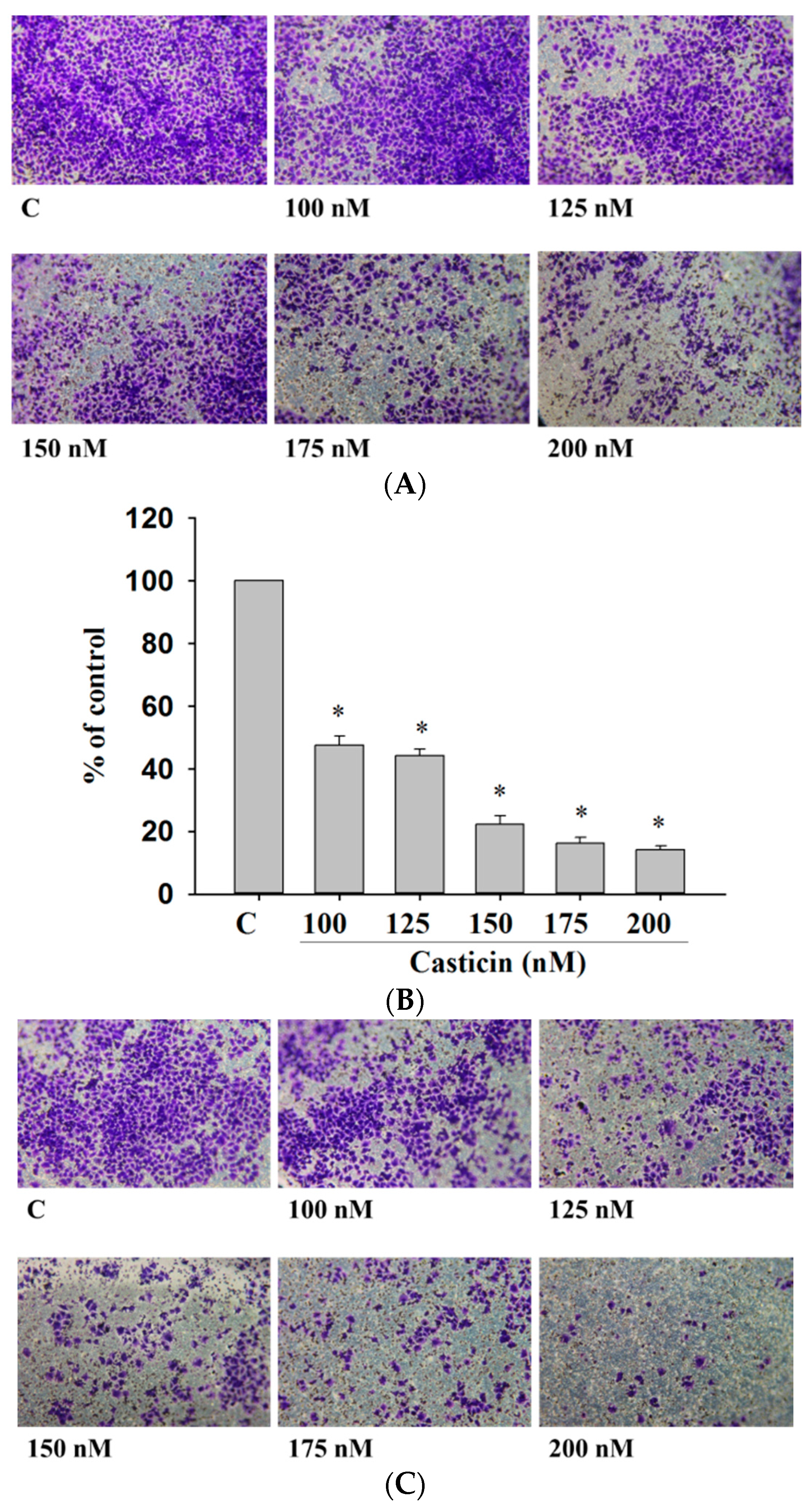
2.4. Casticin Inhibited the Cell Migration and Invasion of A375.S2 Cells
2.5. Casticin Downregulates the Proteolytic Activity and Protein Expression of MMP-2
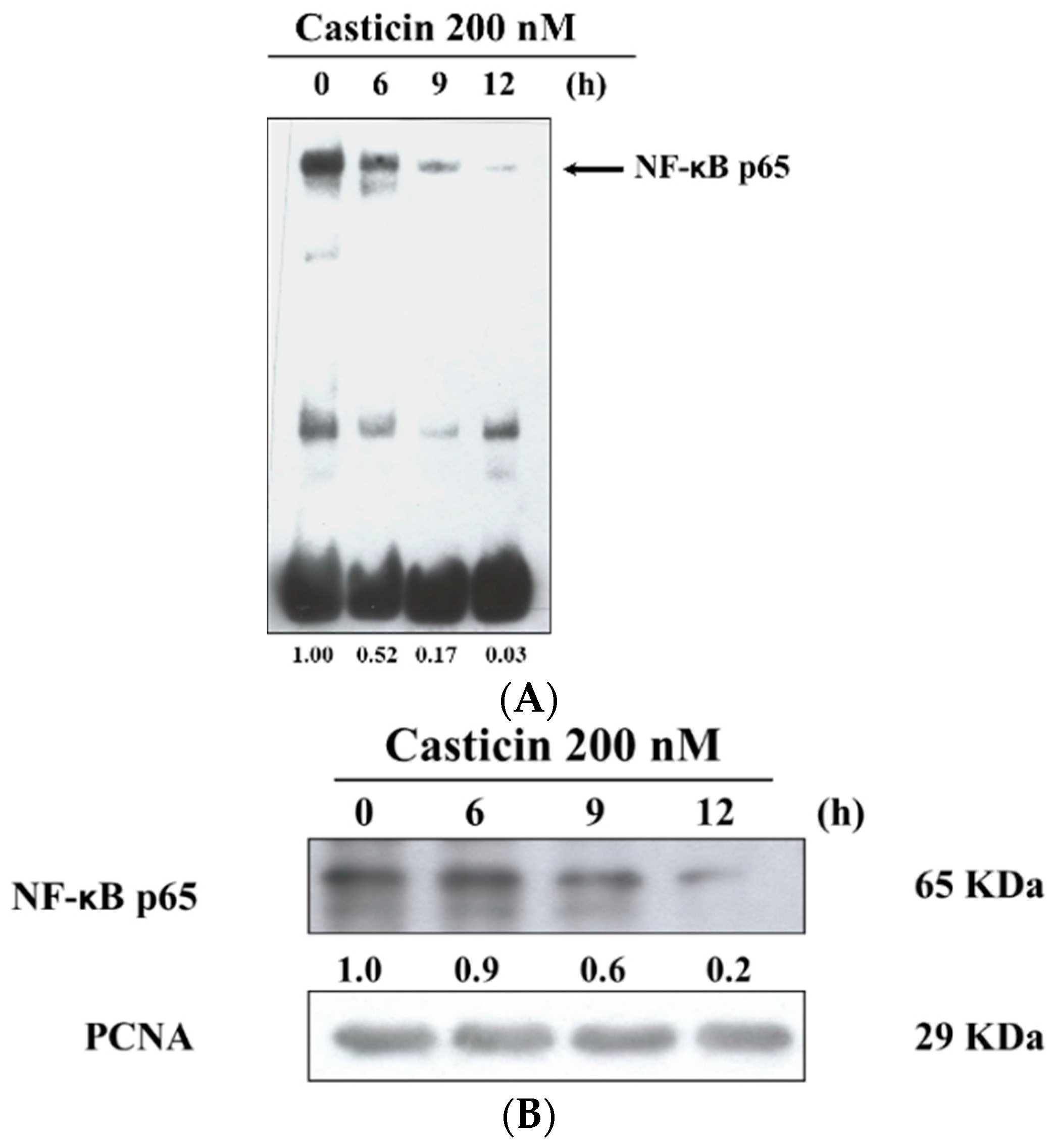
2.6. Casticin Inhibits the Binding and Protein Expression of NF-κB
2.7. Casticin Alters the Levels of Proteins Associated with Migration and Invasion of A375.S2 Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Cell’s Viability Assays
4.4. Wound Healing Mobility Assay
4.5. Adhesion Assay
4.6. Cell Migration and Invasion Assay
4.7. Gelatin Zymography Assay
4.8. Nuclear Extract Preparation.
4.9. Western Blotting Assay
4.10. Electrophoretic Mobility shift Assay (EMSA)
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bravo-Cordero, J.J.; Hodgson, L.; Condeelis, J. Directed cell invasion and migration during metastasis. Curr. Opin. Cell Biol. 2012, 24, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.; Ward, E.; Brawley, O.; Jemal, A. Cancer statistics, 2011: The impact of eliminating socioeconomic and racial disparities on premature cancer deaths. CA Cancer J. Clin. 2011, 61, 212–236. [Google Scholar] [CrossRef] [PubMed]
- Webb, C.P.; Vande Woude, G.F. Genes that regulate metastasis and angiogenesis. J. Neurooncol. 2000, 50, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Mukhtar, H. Cancer and metastasis: Prevention and treatment by green tea. Cancer Metastasis Rev. 2010, 29, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.J.; Kim, Y.; Kang, H.; Sull, J.W.; Kim, Y.S.; Jang, S.W.; Ko, J. Inhibitory effect of Trolox on the migration and invasion of human lung and cervical cancer cells. Int. J. Mol. Med. 2012, 29, 245–251. [Google Scholar] [PubMed]
- Tsai, J.P.; Hsiao, P.C.; Yang, S.F.; Hsieh, S.C.; Bau, D.T.; Ling, C.L.; Pai, C.L.; Hsieh, Y.H. Licochalcone A suppresses migration and invasion of human hepatocellular carcinoma cells through downregulation of MKK4/JNK via NF-kappaB mediated urokinase plasminogen activator expression. PLoS ONE 2014, 9, e86537. [Google Scholar] [CrossRef] [PubMed]
- Tryggvason, K.; Hoyhtya, M.; Pyke, C. Type IV collagenases in invasive tumors. Breast Cancer Res. Treat. 1993, 24, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Koshikawa, N.; Giannelli, G.; Cirulli, V.; Miyazaki, K.; Quaranta, V. Role of cell surface metalloprotease MT1-MMP in epithelial cell migration over laminin-5. J. Cell Biol. 2000, 148, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Lochter, A.; Bissell, M.J. An odyssey from breast to bone: Multi-step control of mammary metastases and osteolysis by matrix metalloproteinases. APMIS 1999, 107, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.C.; Zhou, M.; Shankavaram, U.; Peng, G.; Wahl, L.M. Differential regulation of lipopolysaccharide-induced monocyte matrix metalloproteinase (MMP)-1 and MMP-9 by p38 and extracellular signal-regulated kinase 1/2 mitogen-activated protein kinases. J. Immunol. 2003, 170, 6244–6249. [Google Scholar] [CrossRef] [PubMed]
- Lynch, C.C.; Matrisian, L.M. Matrix metalloproteinases in tumor-host cell communication. Differentiation 2002, 70, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Gialeli, C.; Theocharis, A.D.; Karamanos, N.K. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. FEBS J. 2011, 278, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Ko, W.G.; Kang, T.H.; Lee, S.J.; Kim, N.Y.; Kim, Y.C.; Sohn, D.H.; Lee, B.H. Polymethoxyflavonoids from Vitex rotundifolia inhibit proliferation by inducing apoptosis in human myeloid leukemia cells. Food Chem. Toxicol. 2000, 38, 861–865. [Google Scholar] [CrossRef]
- Lee, C.; Lee, J.W.; Jin, Q.; Lee, H.J.; Lee, S.J.; Lee, D.; Lee, M.K.; Lee, C.K.; Hong, J.T.; Lee, M.K.; Hwang, B.Y. Anti-inflammatory constituents from the fruits of Vitex rotundifolia. Bioorg. Med. Chem. Lett. 2013, 23, 6010–6014. [Google Scholar] [CrossRef] [PubMed]
- Sohn, S.H.; Ko, E.; Oh, B.G.; Kim, S.H.; Kim, Y.; Shin, M.; Hong, M.; Bae, H. Inhibition effects of Vitex rotundifolia on inflammatory gene expression in A549 human epithelial cells. Ann. Allergy Asthma Immunol. 2009, 103, 152–159. [Google Scholar] [CrossRef]
- Mainardi, T.; Kapoor, S.; Bielory, L. Complementary and alternative medicine: Herbs, phytochemicals and vitamins and their immunologic effects. J. Allergy Clin. Immunol. 2009, 123, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Yamamoto, M.; Yanaka, T.; Ito, Y.; Nohara, T. Ten new labdane-type diterpenes from the fruit of Vitex rotundifolia. Chem. Pharm. Bull. 2001, 49, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhou, Q.; Liu, C.; Tao, M.L. Drug screening study using glioma stem-like cells. Mol. Med. Rep. 2012, 6, 1117–1120. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yang, Y.; Tian, L.; Sheng, X.F.; Liu, F.; Cao, J.G. Casticin-induced apoptosis involves death receptor 5 upregulation in hepatocellular carcinoma cells. World J. Gastroenterol. 2011, 17, 4298–4307. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Fang, Z.; Wu, Y.; Zhang, H. [Chemical constituents of the fruits of Vitex trifolia L]. Zhongguo Zhong Yao Za Zhi 1996, 21, 167–168. [Google Scholar] [PubMed]
- Lin, S.; Zhang, H.; Han, T.; Wu, J.Z.; Rahman, K.; Qin, L.P. In vivo effect of casticin on acute inflammation. Zhong Xi Yi Jie He Xue Bao 2007, 5, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Cao, J.; Tian, L.; Liu, F.; Sheng, X. Induction of apoptosis by casticin in cervical cancer cells through reactive oxygen species-mediated mitochondrial signaling pathways. Oncol. Rep. 2011, 26, 1287–1294. [Google Scholar] [PubMed]
- Kikuchi, H.; Yuan, B.; Yuhara, E.; Takagi, N.; Toyoda, H. Involvement of histone H3 phosphorylation through p38 MAPK pathway activation in casticin-induced cytocidal effects against the human promyelocytic cell line HL-60. Int. J. Oncol. 2013, 43, 2046–2056. [Google Scholar] [PubMed]
- Zhou, Y.; Tian, L.; Long, L.; Quan, M.; Liu, F.; Cao, J. Casticin potentiates TRAIL-induced apoptosis of gastric cancer cells through endoplasmic reticulum stress. PLoS ONE 2013, 8, e58855. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Cao, X.; Liu, Z.; Guo, H.; Ren, K.; Quan, M.; Zhou, Y.; Xiang, H.; Cao, J. Casticin suppresses self-renewal and invasion of lung cancer stem-like cells from A549 cells through down-regulation of pAkt. Acta Biochim. Biophys. Sin. 2014, 46, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, M.I.; Jalil, S.; Nawaz, S.A.; Khan, K.M.; Tareen, R.B. Antiinflammatory and lipoxygenase inhibitory compounds from Vitex agnus-castus. Phytother. Res. 2009, 23, 1336–1339. [Google Scholar] [CrossRef] [PubMed]
- Mesaik, M.A.; Murad, S.; Khan, K.M.; Tareen, R.B.; Ahmed, A.; Choudhary, M.I. Isolation and immunomodulatory properties of a flavonoid, casticin from Vitex agnus-castus. Phytother. Res. 2009, 23, 1516–1520. [Google Scholar] [CrossRef] [PubMed]
- Li, W.X.; Cui, C.B.; Cai, B.; Wang, H.Y.; Yao, X.S. Flavonoids from Vitex trifolia L. inhibit cell cycle progression at G2/M phase and induce apoptosis in mammalian cancer cells. J. Asian Nat. Prod. Res. 2005, 7, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Liou, C.J.; Len, W.B.; Wu, S.J.; Lin, C.F.; Wu, X.L.; Huang, W.C. Casticin inhibits COX-2 and iNOS expression via suppression of NF-kappaB and MAPK signaling in lipopolysaccharide-stimulated mouse macrophages. J. Ethnopharmacol. 2014, 158, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Jung, K.H.; Lee, H.; Park, S.; Choi, W.; Bae, H. Casticin, an active compound isolated from Vitex Fructus, ameliorates the cigarette smoke-induced acute lung inflammatory response in a murine model. Int. Immunopharmacol. 2015, 28, 1097–1101. [Google Scholar] [CrossRef] [PubMed]
- Koh, D.J.; Ahn, H.S.; Chung, H.S.; Lee, H.; Kim, Y.; Lee, J.Y.; Kim, D.G.; Hong, M.; Shin, M.; Bae, H. Inhibitory effects of casticin on migration of eosinophil and expression of chemokines and adhesion molecules in A549 lung epithelial cells via NF-kappaB inactivation. J. Ethnopharmacol. 2011, 136, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.J.; Yen, G.C. The in vitro and in vivo experimental evidences disclose the chemopreventive effects of Ganoderma lucidum on cancer invasion and metastasis. Clin. Exp. Metastasis 2010, 27, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix metalloproteinases: Regulators of the tumor microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Babykutty, S.; Suboj, P.; Srinivas, P.; Nair, A.S.; Chandramohan, K.; Gopala, S. Insidious role of nitric oxide in migration/invasion of colon cancer cells by upregulating MMP-2/9 via activation of cGMP-PKG-ERK signaling pathways. Clin. Exp. Metastasis 2012, 29, 471–492. [Google Scholar] [CrossRef] [PubMed]
- Dung, T.D.; Feng, C.C.; Kuo, W.W.; Pai, P.; Chung, L.C.; Chang, S.H.; Hsu, H.H.; Tsai, F.J.; Lin, Y.M.; Huang, C.Y. Suppression of plasminogen activators and the MMP-2/-9 pathway by a Zanthoxylum avicennae extract to inhibit the HA22T human hepatocellular carcinoma cell migration and invasion effects in vitro and in vivo via phosphatase 2A activation. Biosci. Biotechnol. Biochem. 2013, 77, 1814–1821. [Google Scholar] [CrossRef] [PubMed]
- Garg, P.; Sarma, D.; Jeppsson, S.; Patel, N.R.; Gewirtz, A.T.; Merlin, D.; Sitaraman, S.V. Matrix metalloproteinase-9 functions as a tumor suppressor in colitis-associated cancer. Cancer Res. 2010, 70, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.B.; Krueger, J.S.; Kondapaka, S.B.; Diglio, C.A. Mitogen-activated protein kinase (MAPK) regulates the expression of progelatinase B (MMP-9) in breast epithelial cells. Int. J. Cancer 1999, 82, 268–273. [Google Scholar] [CrossRef]
- Liotta, L.A.; Tryggvason, K.; Garbisa, S.; Hart, I.; Foltz, C.M.; Shafie, S. Metastatic potential correlates with enzymatic degradation of basement membrane collagen. Nature 1980, 284, 67–68. [Google Scholar] [CrossRef] [PubMed]
- Anand, M.; Van Meter, T.E.; Fillmore, H.L. Epidermal growth factor induces matrix metalloproteinase-1 (MMP-1) expression and invasion in glioma cell lines via the MAPK pathway. J. Neurooncol. 2011, 104, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, C.; Li, L.; Shanmugam, R.; Bhat-Nakshatri, P.; Jayaprakasan, V.; Baldridge, L.A.; Gardner, T.; Smith, M.; Nakshatri, H.; Cheng, L. Nuclear factor-kappaB is constitutively activated in prostate cancer in vitro and is overexpressed in prostatic intraepithelial neoplasia and adenocarcinoma of the prostate. Clin. Cancer Res. 2004, 10, 5501–5507. [Google Scholar] [CrossRef] [PubMed]
- Price, J.T.; Tiganis, T.; Agarwal, A.; Djakiew, D.; Thompson, E.W. Epidermal growth factor promotes MDA-MB-231 breast cancer cell migration through a phosphatidylinositol 3′-kinase and phospholipase C-dependent mechanism. Cancer Res. 1999, 59, 5475–5478. [Google Scholar] [PubMed]
- Pasparakis, M.; Courtois, G.; Hafner, M.; Schmidt-Supprian, M.; Nenci, A.; Toksoy, A.; Krampert, M.; Goebeler, M.; Gillitzer, R.; Israel, A.; et al. TNF-mediated inflammatory skin disease in mice with epidermis-specific deletion of IKK2. Nature 2002, 417, 861–866. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-M.; Velmurugan, B.K.; Kuo, W.-W.; Chen, Y.-S.; Ho, T.-J.; Tsai, C.-T.; Ye, C.-X.; Tsai, C.-H.; Tsai, F.-J.; Huang, C.-Y. Inhibitory effect of alpinate Oxyphyllae fructus extracts on Ang II-induced cardiac pathological remodeling-related pathways in H9c2 cardiomyoblast cells. Biomedicine 2013, 3, 148–152. [Google Scholar] [CrossRef]
- Wu, S.H.; Hsiao, Y.T.; Kuo, C.L.; Yu, F.S.; Hsu, S.C.; Wu, P.P.; Chen, J.C.; Hsia, T.C.; Liu, H.C.; Hsu, W.H.; et al. Bufalin Inhibits NCI-H460 Human Lung Cancer Cell Metastasis In Vitro by Inhibiting MAPKs, MMPs, and NF-kappaB Pathways. Am. J. Chin. Med. 2015, 43, 1247–1264. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.L.; Lin, J.H.; Lien, J.C.; Hsu, S.C.; Chueh, F.S.; Yu, C.C.; Wu, P.P.; Huang, Y.P.; Lin, J.G.; Chung, J.G. The crude extract of Corni Fructus inhibits the migration and invasion of U-2 OS human osteosarcoma cells through the inhibition of matrix metalloproteinase-2/-9 by MAPK signaling. Environ. Toxicol. 2015, 30, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Casticin are available from the Sigma-Aldrich Corp. (St. Louis, MO, USA).









© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.-Y.; Lien, J.-C.; Huang, Y.-P.; Liao, C.-L.; Lin, J.-J.; Fan, M.-J.; Ko, Y.-C.; Hsiao, Y.-P.; Lu, H.-F.; Chung, J.-G. Casticin Inhibits A375.S2 Human Melanoma Cell Migration/Invasion through Downregulating NF-κB and Matrix Metalloproteinase-2 and -1. Molecules 2016, 21, 384. https://doi.org/10.3390/molecules21030384
Wu Z-Y, Lien J-C, Huang Y-P, Liao C-L, Lin J-J, Fan M-J, Ko Y-C, Hsiao Y-P, Lu H-F, Chung J-G. Casticin Inhibits A375.S2 Human Melanoma Cell Migration/Invasion through Downregulating NF-κB and Matrix Metalloproteinase-2 and -1. Molecules. 2016; 21(3):384. https://doi.org/10.3390/molecules21030384
Chicago/Turabian StyleWu, Zih-Yun, Jin-Cherng Lien, Yi-Ping Huang, Ching-Lung Liao, Jen-Jyh Lin, Ming-Jen Fan, Yang-Ching Ko, Yu-Ping Hsiao, Hsu-Feng Lu, and Jing-Gung Chung. 2016. "Casticin Inhibits A375.S2 Human Melanoma Cell Migration/Invasion through Downregulating NF-κB and Matrix Metalloproteinase-2 and -1" Molecules 21, no. 3: 384. https://doi.org/10.3390/molecules21030384