

Activated Leukocyte Cell Adhesion Molecule (ALCAM), a Potential ‘Seed’ and ‘Soil’ Receptor in the Peritoneal Metastasis of Gastrointestinal Cancers
 ,
,
Abstract
:1. Introduction
2. Results
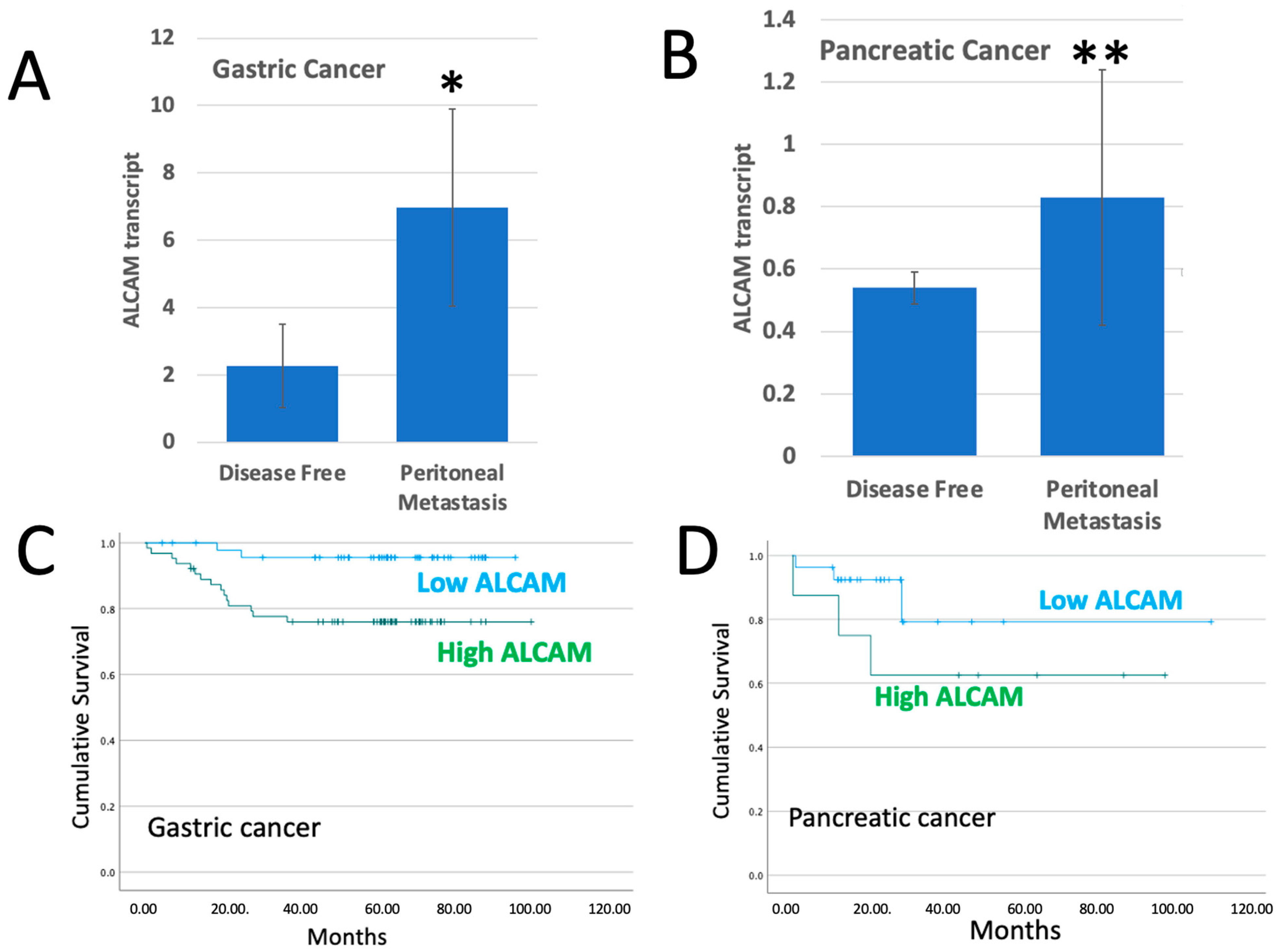
2.1. ALCAM in Tumours Which Developed Peritoneal Metastasis
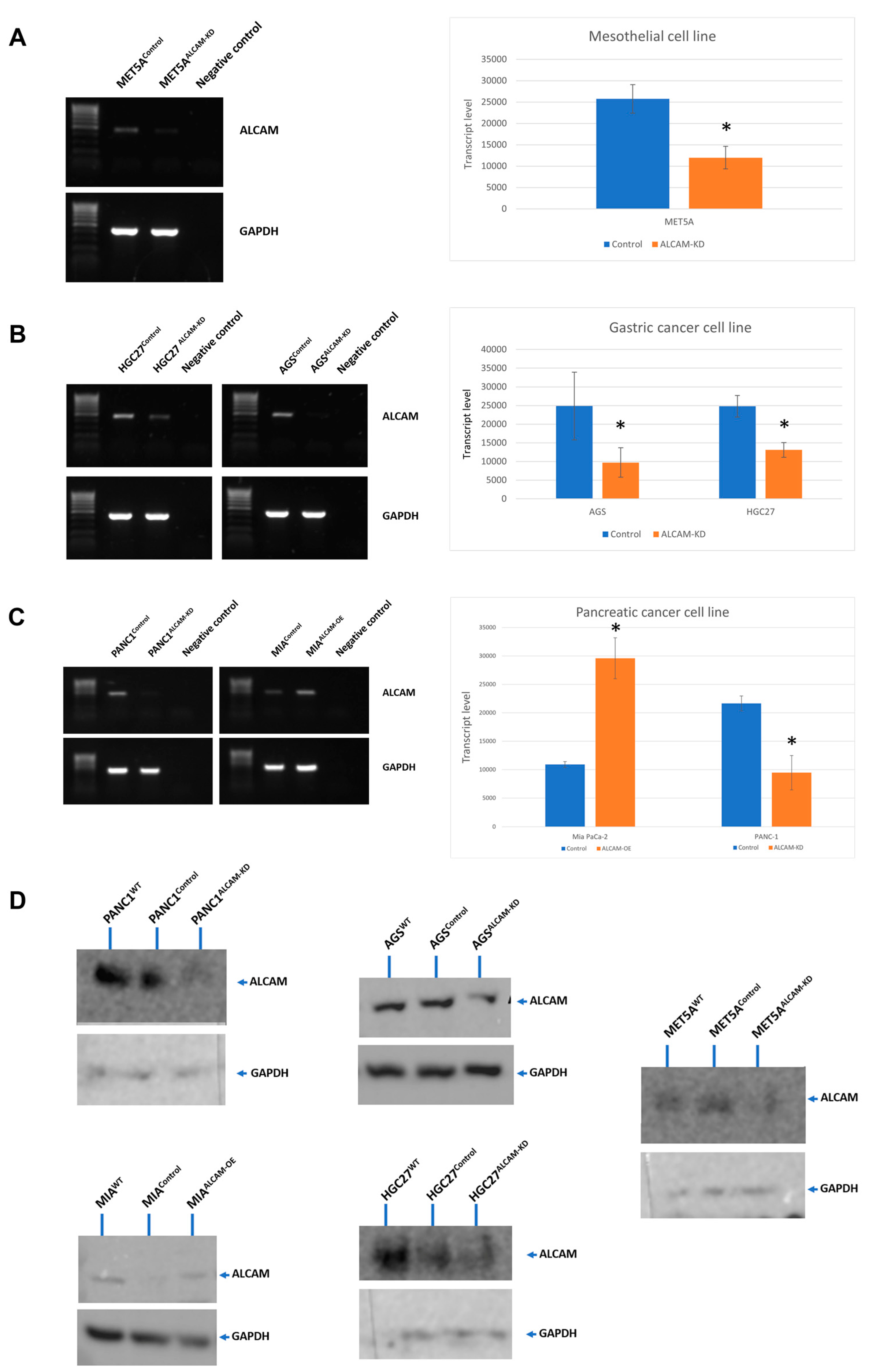
2.2. The Creation of Cell Models with Altered Levels of ALCAM
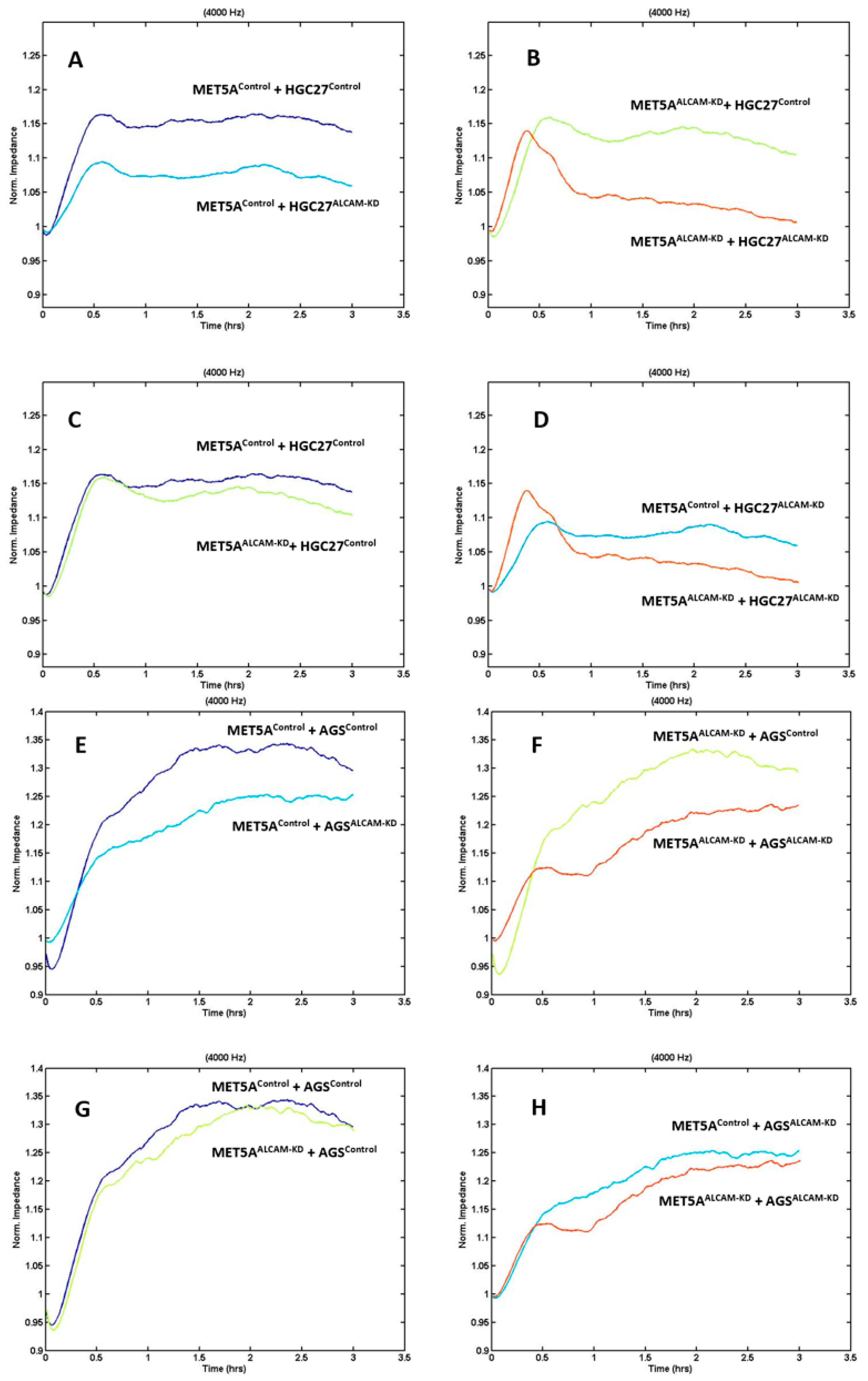
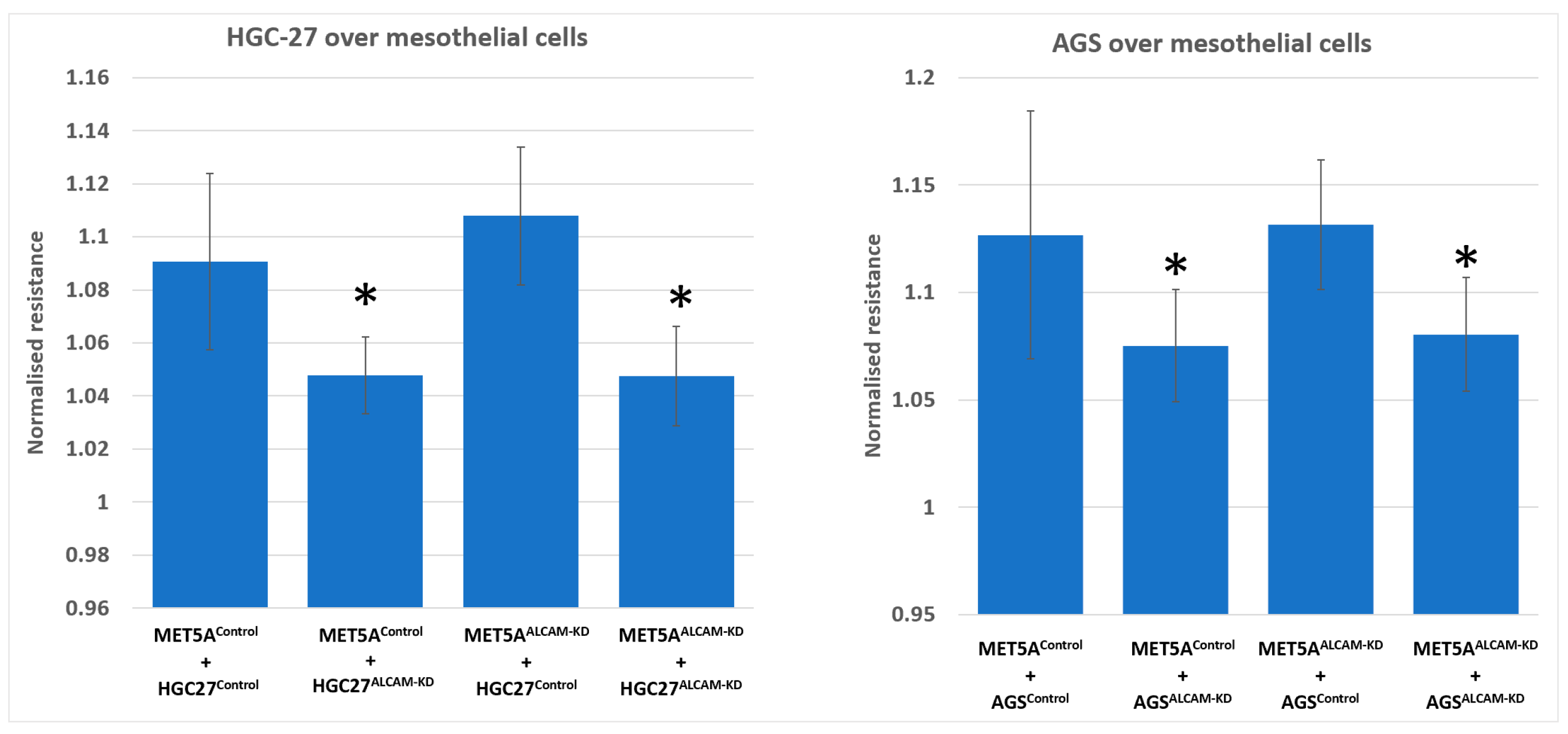
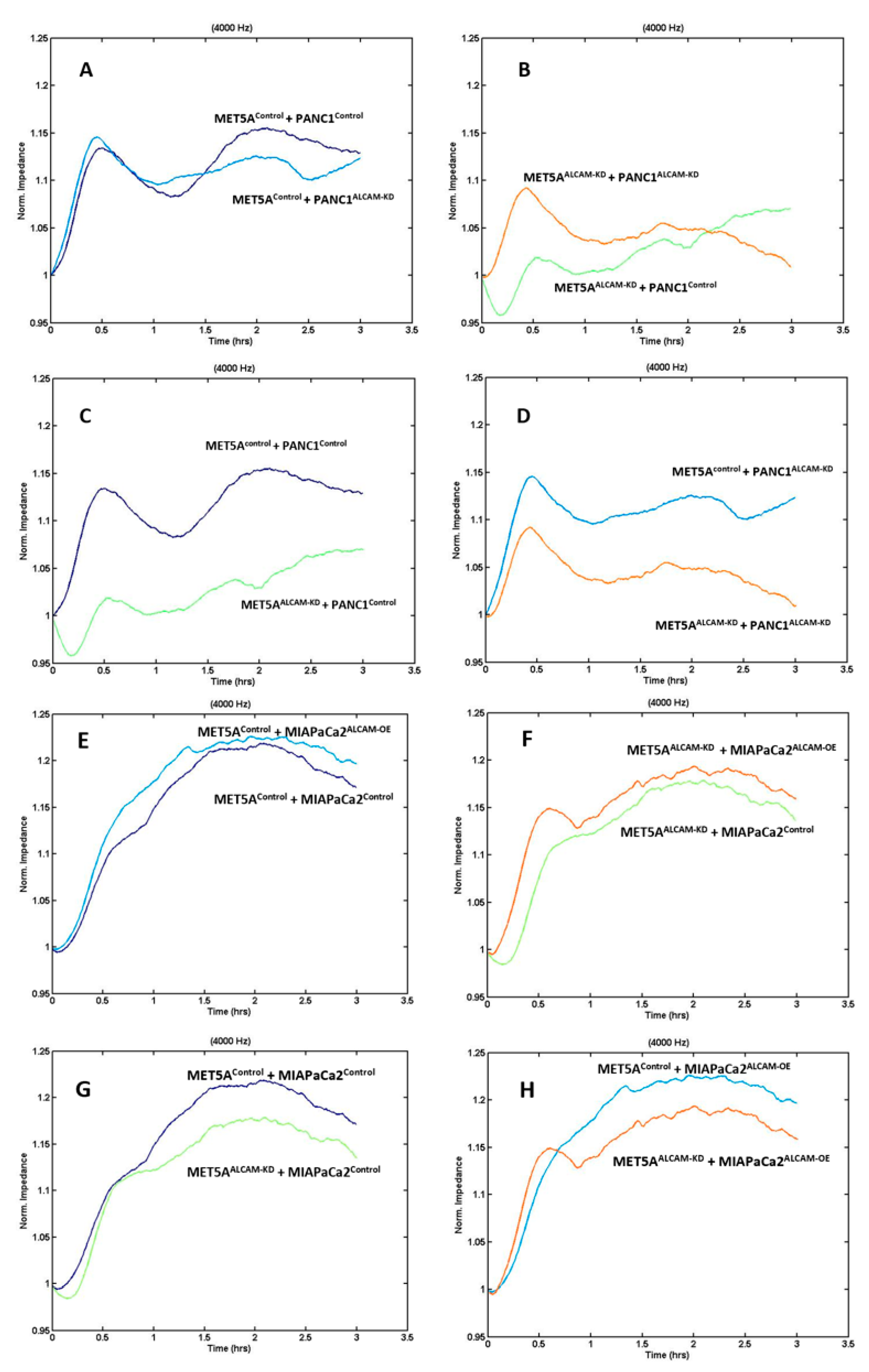
2.3. Dynamic Monitoring of Gastric Tumour-Mesothelial Interaction Assocaited with ALCAM Level Alteration
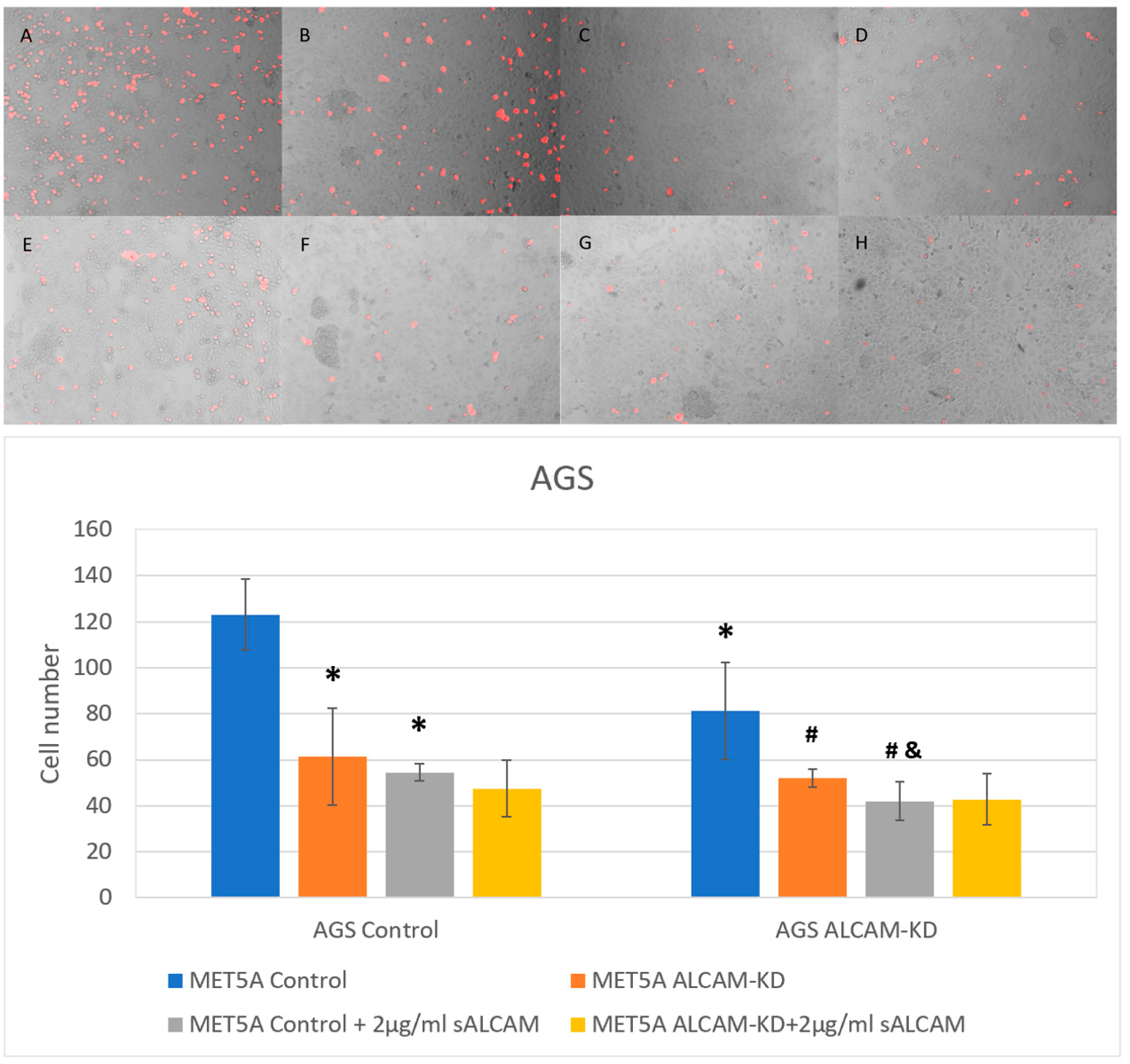
2.4. Dynamic Monitoring of Pancreatic Tumour-Mesothelial Interactions Associated with ALCAM Alterations
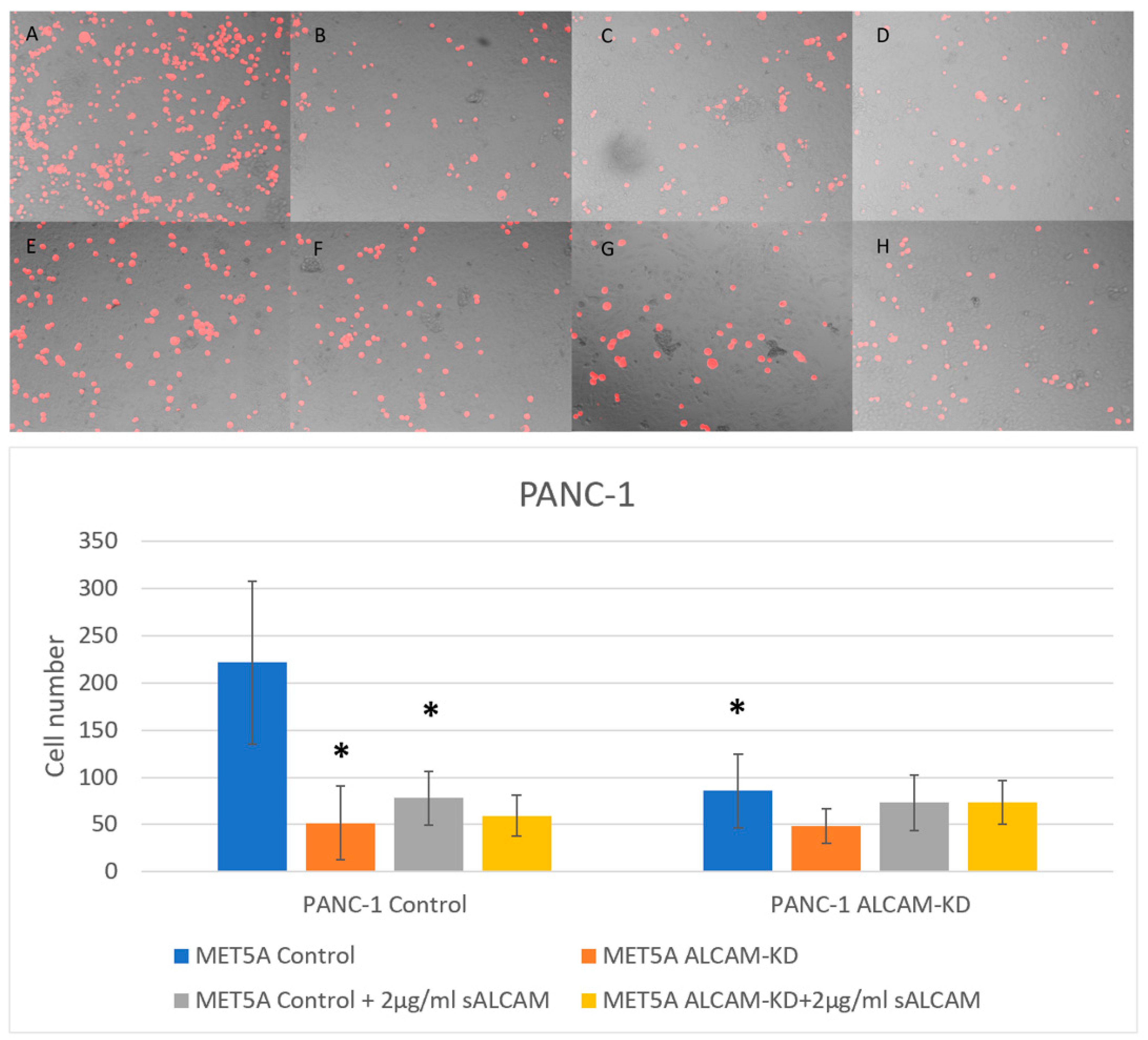
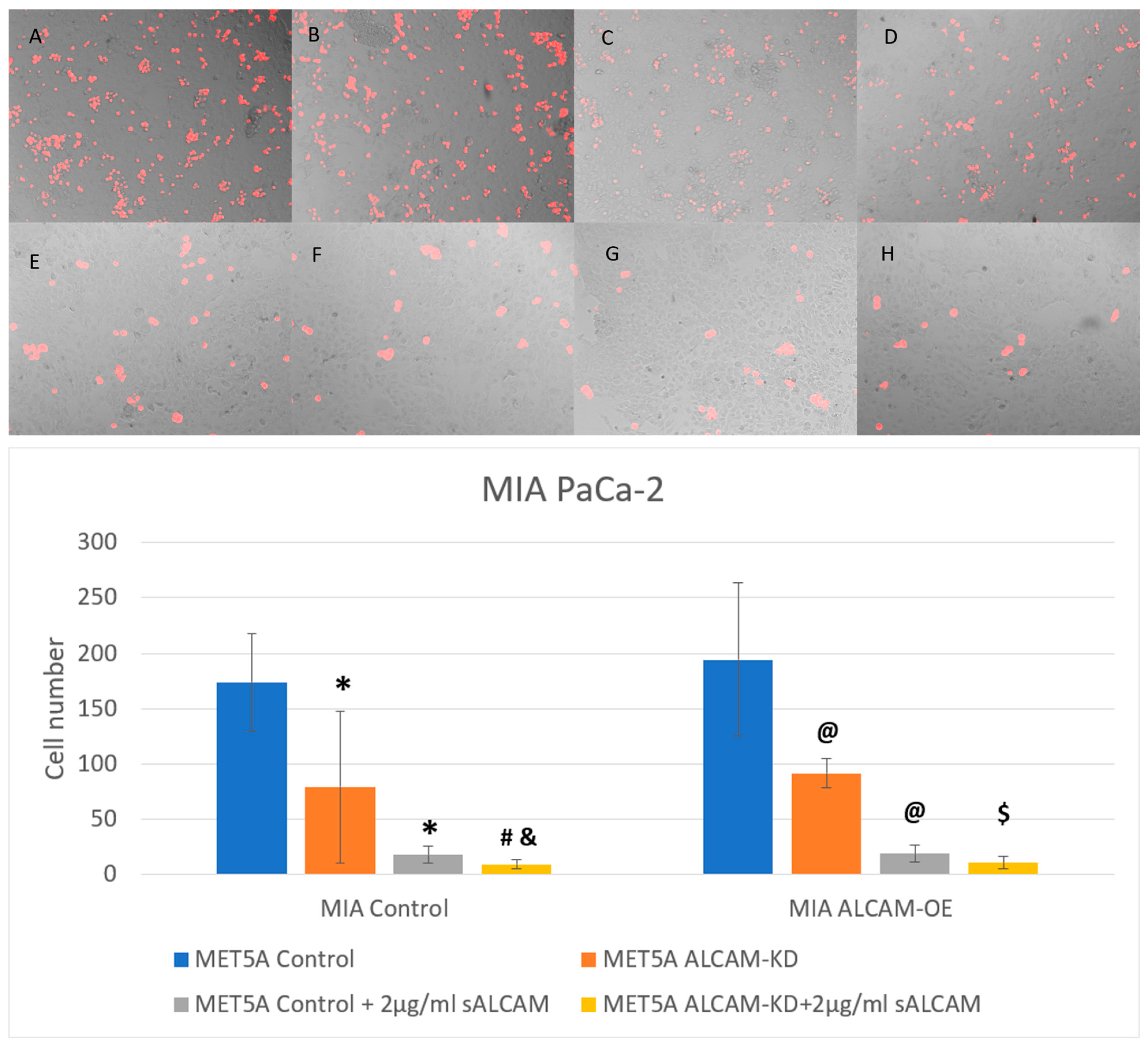
2.5. DiI Based Pancreatic Tumour-Mesothelial Cell Interactions
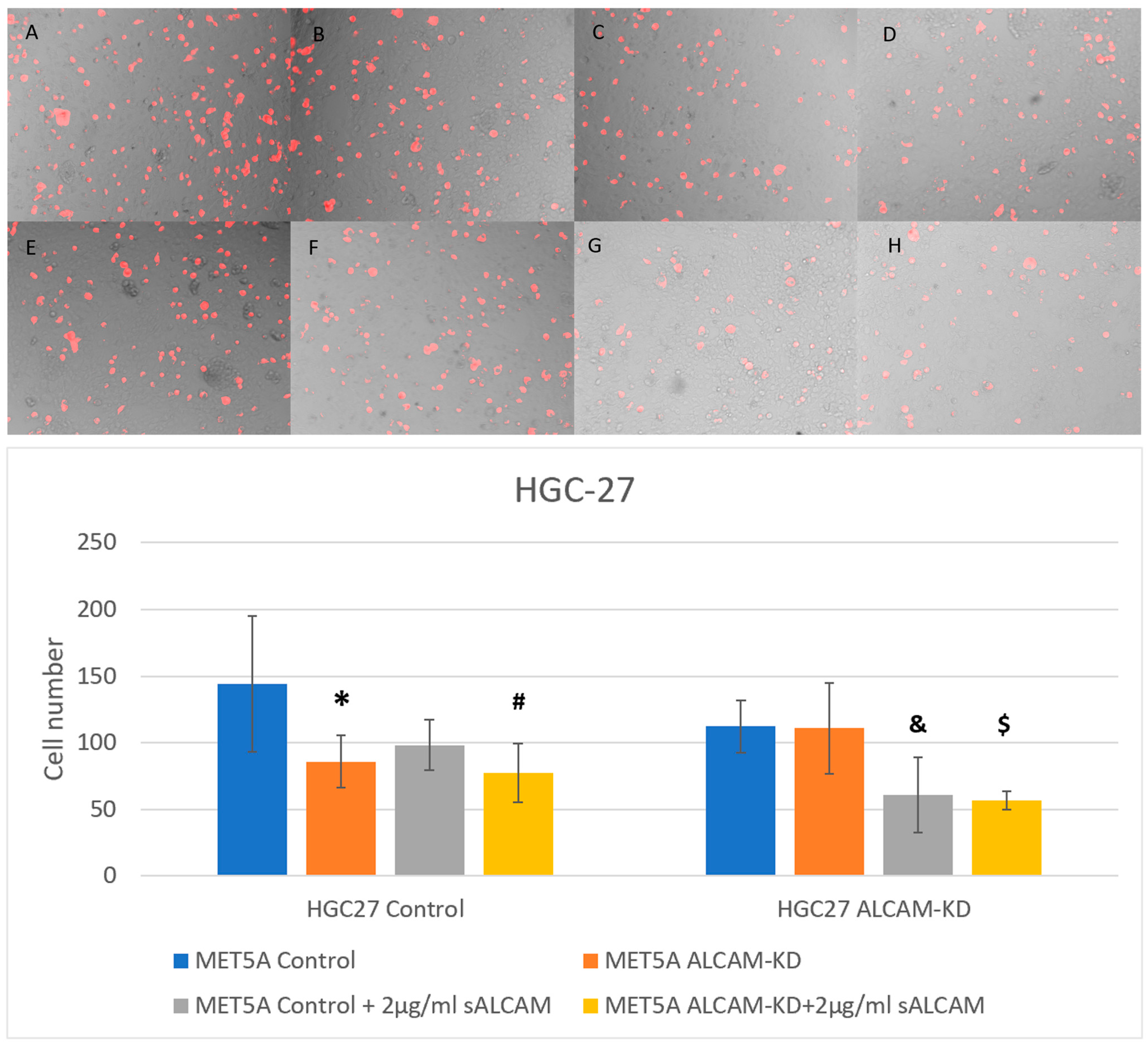
2.6. DiI Based Gastric Tumour-Mesothelial Cell Interactions
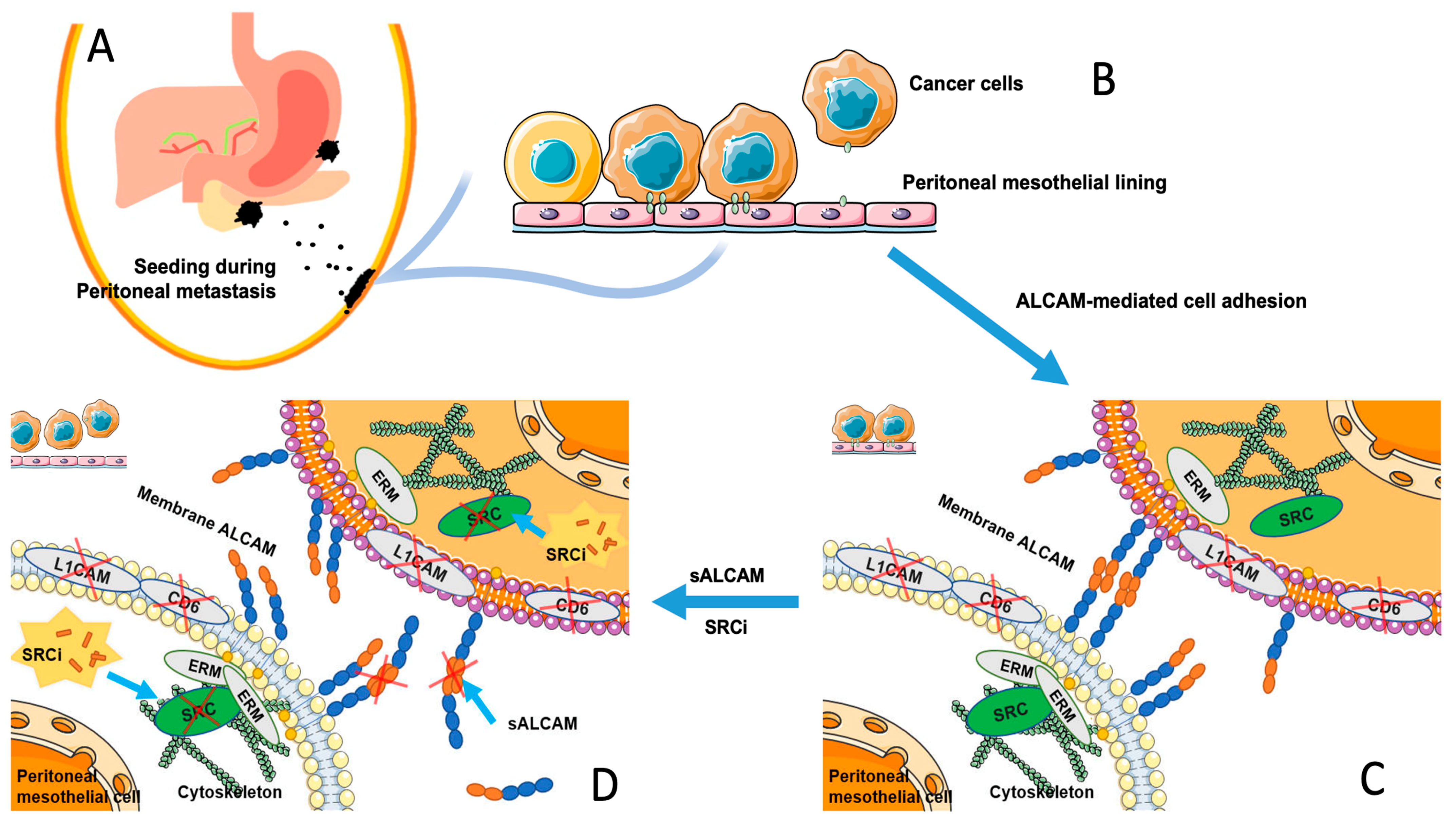
2.7. The Interaction of Tumour-Mesothelial Cells Was Primarily Due to the Action of ALCAM and Mediated by the SRC Pathway
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cell Lines and Cell Cultures
5.2. Key Reagents
5.3. Generation of ALCAM Modified Cells
5.4. Clinical Cohorts
5.5. Determination of ALCAM in Tissues
5.6. Tumour-Mesothelial Interaction Assay
5.7. Dynamic Monitoring of Tumour-Mesothelial Interactions
5.8. Protein Preparation and Protein Electrophesis
5.9. Statistical Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Douglass, H.O., Jr.; Penetrante, R.B. Pancreatic cancer. Why patients die. Int. J. Pancreatol. 1990, 7, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Garrison, R.N.; Kaelin, L.D.; Galloway, R.H.; Heuser, L.S. Malignant ascites. Clinical and experimental observations. Ann. Surg. 1986, 203, 644–651. [Google Scholar] [CrossRef] [PubMed]
- del Castillo, C.F.; Warshaw, L. Peritoneal metastases in pancreatic carcinoma. Hepatogastroenterology 1993, 40, 430–432. [Google Scholar] [PubMed]
- Sugarbaker, P.H.; Ryan, D.P. Cytoreductive surgery plus hyperthermic perioperative chemotherapy to treat peritoneal metastases from colorectal cancer: Standard of care or an experimental approach? Lancet Oncol. 2012, 13, e362–e369. [Google Scholar] [CrossRef]
- Tewari, D.; Java, J.J.; Salani, R.; Armstrong, D.K.; Markman, M.; Herzog, T.; Monk, B.J.; Chan, J.K. Long-term survival advantage and prognostic factors associated with intraperitoneal chemotherapy treatment in advanced ovarian cancer: A gynecologic oncology group study. J. Clin. Oncol. 2015, 33, 1460–1466. [Google Scholar] [CrossRef]
- Leichman, L.; Silberman, H.; Leichman, C.G.; Spears, C.P.; Ray, M.; Muggia, F.M.; Kiyabu, M.; Radin, R.; Laine, L.; Stain, S.; et al. Preoperative systemic chemotherapy followed by adjuvant postoperative intraperitoneal therapy for gastric cancer: A University of Southern California pilot program. J. Clin. Oncol. 1992, 10, 1933–1942. [Google Scholar] [CrossRef]
- Cortes-Guiral, D.; Hubner, M.; Alyami, M.; Bhatt, A.; Ceelen, W.; Glehen, O.; Lordick, F.; Ramsay, R.; Sgarbura, O.; Van Der Speeten, K.; et al. Primary and metastatic peritoneal surface malignancies. Nat. Rev. Dis. Prim. 2021, 7, 91. [Google Scholar] [CrossRef]
- Burg, L.; Timmermans, M.; van der Aa, M.; Boll, D.; Rovers, K.; de Hingh, I.; van Altena, A. Incidence and predictors of peritoneal metastases of gynecological origin: A population-based study in the Netherlands. J. Gynecol. Oncol. 2020, 31, e58. [Google Scholar] [CrossRef]
- Thomassen, I.; van Gestel, Y.R.; van Ramshorst, B.; Luyer, M.D.; Bosscha, K.; Nienhuijs, S.W.; Lemmens, V.E.; de Hingh, I.H. Peritoneal carcinomatosis of gastric origin: A population-based study on incidence, survival and risk factors. Int. J. Cancer 2014, 134, 622–628. [Google Scholar] [CrossRef]
- Tanaka, M.; Mihaljevic, A.L.; Probst, P.; Heckler, M.; Klaiber, U.; Heger, U.; Buchler, M.W.; Hackert, T. Meta-analysis of recurrence pattern after resection for pancreatic cancer. Br. J. Surg. 2019, 106, 1590–1601. [Google Scholar] [CrossRef]
- Chu, D.Z.; Lang, N.P.; Thompson, C.; Osteen, P.K.; Westbrook, K.C. Peritoneal carcinomatosis in nongynecologic malignancy. A prospective study of prognostic factors. Cancer 1989, 63, 364–367. [Google Scholar] [CrossRef] [PubMed]
- Segelman, J.; Granath, F.; Holm, T.; Machado, M.; Mahteme, H.; Martling, A. Incidence, prevalence and risk factors for peritoneal carcinomatosis from colorectal cancer. Br. J. Surg. 2012, 99, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, M.; Solon, J.; Chang, K.H.; Deady, S.; Moran, B.; Cahill, R.; Shields, C.; Mulsow, J. Peritoneal metastases from extra-abdominal cancer—A population-based study. Eur. J. Surg. Oncol. 2018, 44, 1811–1817. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, B.; Arvieux, C.; Glehen, O.; Beaujard, A.C.; Rivoire, M.; Baulieux, J.; Fontaumard, E.; Brachet, A.; Caillot, J.L.; Faure, J.L.; et al. Peritoneal carcinomatosis from non-gynecologic malignancies: Results of the EVOCAPE 1 multicentric prospective study. Cancer 2000, 88, 358–363. [Google Scholar] [CrossRef]
- Cao, C.; Yan, T.D.; Black, D.; Morris, D.L. A systematic review and meta-analysis of cytoreductive surgery with perioperative intraperitoneal chemotherapy for peritoneal carcinomatosis of colorectal origin. Ann. Surg. Oncol. 2009, 16, 2152–2165. [Google Scholar] [CrossRef] [PubMed]
- Elias, D.; Gilly, F.; Boutitie, F.; Quenet, F.; Bereder, J.M.; Mansvelt, B.; Lorimier, G.; Dube, P.; Glehen, O. Peritoneal colorectal carcinomatosis treated with surgery and perioperative intraperitoneal chemotherapy: Retrospective analysis of 523 patients from a multicentric French study. J. Clin. Oncol. 2010, 28, 63–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glehen, O.; Kwiatkowski, F.; Sugarbaker, P.H.; Elias, D.; Levine, E.A.; De Simone, M.; Barone, R.; Yonemura, Y.; Cavaliere, F.; Quenet, F.; et al. Cytoreductive surgery combined with perioperative intraperitoneal chemotherapy for the management of peritoneal carcinomatosis from colorectal cancer: A multi-institutional study. J. Clin. Oncol. 2004, 22, 3284–3292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glehen, O.; Mohamed, F.; Gilly, F.N. Peritoneal carcinomatosis from digestive tract cancer: New management by cytoreductive surgery and intraperitoneal chemohyperthermia. Lancet Oncol. 2004, 5, 219–228. [Google Scholar] [CrossRef]
- Raspagliesi, F.; Kusamura, S.; Campos Torres, J.C.; de Souza, G.A.; Ditto, A.; Zanaboni, F.; Younan, R.; Baratti, D.; Mariani, L.; Laterza, B.; et al. Cytoreduction combined with intraperitoneal hyperthermic perfusion chemotherapy in advanced/recurrent ovarian cancer patients: The experience of National Cancer Institute of Milan. Eur. J. Surg. Oncol. 2006, 32, 671–675. [Google Scholar] [CrossRef]
- Fujimoto, S.; Takahashi, M.; Mutou, T.; Kobayashi, K.; Toyosawa, T. Successful intraperitoneal hyperthermic chemoperfusion for the prevention of postoperative peritoneal recurrence in patients with advanced gastric carcinoma. Cancer 1999, 85, 529–534. [Google Scholar] [CrossRef]
- Leake, P.A.; Cardoso, R.; Seevaratnam, R.; Lourenco, L.; Helyer, L.; Mahar, A.; Rowsell, C.; Coburn, N.G. A systematic review of the accuracy and utility of peritoneal cytology in patients with gastric cancer. Gastric. Cancer 2012, 15 (Suppl. 1), S27–S37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagotti, A.; Vizzielli, G.; Fanfani, F.; Costantini, B.; Ferrandina, G.; Gallotta, V.; Gueli Alletti, S.; Tortorella, L.; Scambia, G. Introduction of staging laparoscopy in the management of advanced epithelial ovarian, tubal and peritoneal cancer: Impact on prognosis in a single institution experience. Gynecol. Oncol. 2013, 131, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Fagotti, A.; Vizzielli, G.; De Iaco, P.; Surico, D.; Buda, A.; Mandato, V.D.; Petruzzelli, F.; Ghezzi, F.; Garzarelli, S.; Mereu, L.; et al. A multicentric trial (Olympia-MITO 13) on the accuracy of laparoscopy to assess peritoneal spread in ovarian cancer. Am. J. Obstet. Gynecol. 2013, 209, 462.e1–462.e11. [Google Scholar] [CrossRef] [PubMed]
- Havrilesky, L.J.; Cragun, J.M.; Calingaert, B.; Alvarez Secord, A.; Valea, F.A.; Clarke-Pearson, D.L.; Berchuck, A.; Soper, J.T. The prognostic significance of positive peritoneal cytology and adnexal/serosal metastasis in stage IIIA endometrial cancer. Gynecol. Oncol. 2007, 104, 401–405. [Google Scholar] [CrossRef]
- Satoi, S.; Fujii, T.; Yanagimoto, H.; Motoi, F.; Kurata, M.; Takahara, N.; Yamada, S.; Yamamoto, T.; Mizuma, M.; Honda, G.; et al. Multicenter Phase II Study of Intravenous and Intraperitoneal Paclitaxel With S-1 for Pancreatic Ductal Adenocarcinoma Patients With Peritoneal Metastasis. Ann. Surg. 2017, 265, 397–401. [Google Scholar] [CrossRef] [Green Version]
- Zuna, R.E.; Behrens, A. Peritoneal washing cytology in gynecologic cancers: Long-term follow-up of 355 patients. J. Natl. Cancer Inst. 1996, 88, 980–987. [Google Scholar] [CrossRef]
- Mezhir, J.J.; Shah, M.A.; Jacks, L.M.; Brennan, M.F.; Coit, D.G.; Strong, V.E. Positive peritoneal cytology in patients with gastric cancer: Natural history and outcome of 291 patients. Ann. Surg. Oncol. 2010, 17, 3173–3180. [Google Scholar] [CrossRef]
- Ajani, J.A.; Barthel, J.S.; Bekaii-Saab, T.; Bentrem, D.J.; D'Amico, T.A.; Das, P.; Denlinger, C.; Fuchs, C.S.; Gerdes, H.; Hayman, J.A.; et al. Gastric cancer. J. Natl. Compr. Cancer Netw. 2010, 8, 378–409. [Google Scholar] [CrossRef]
- Morgan, R.J., Jr.; Armstrong, D.K.; Alvarez, R.D.; Bakkum-Gamez, J.N.; Behbakht, K.; Chen, L.M.; Copeland, L.; Crispens, M.A.; DeRosa, M.; Dorigo, O.; et al. Ovarian Cancer, Version 1.2016, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2016, 14, 1134–1163. [Google Scholar] [CrossRef]
- Wu, X.; Li, Z.; Li, Z.; Jia, Y.; Shan, F.; Ji, X.; Bu, Z.; Zhang, L.; Wu, A.; Ji, J. Hyperthermic intraperitoneal chemotherapy plus simultaneous versus staged cytoreductive surgery for gastric cancer with occult peritoneal metastasis. J. Surg. Oncol. 2015, 111, 840–847. [Google Scholar] [CrossRef]
- Kuramoto, M.; Shimada, S.; Ikeshima, S.; Matsuo, A.; Yagi, Y.; Matsuda, M.; Yonemura, Y.; Baba, H. Extensive intraoperative peritoneal lavage as a standard prophylactic strategy for peritoneal recurrence in patients with gastric carcinoma. Ann. Surg. 2009, 250, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Paget, S. The distribution of secondary growths in cancer of the breast. 1889. Cancer Metastasis Rev. 1989, 8, 98–101. [Google Scholar] [PubMed]
- Spadea, A.; Rios de la Rosa, J.M.; Tirella, A.; Ashford, M.B.; Williams, K.J.; Stratford, I.J.; Tirelli, N.; Mehibel, M. Evaluating the Efficiency of Hyaluronic Acid for Tumor Targeting via CD44. Mol. Pharm. 2019, 16, 2481–2493. [Google Scholar] [CrossRef] [PubMed]
- Mattheolabakis, G.; Milane, L.; Singh, A.; Amiji, M.M. Hyaluronic acid targeting of CD44 for cancer therapy: From receptor biology to nanomedicine. J. Drug Target. 2015, 23, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Soliman, F.; Ye, L.; Jiang, W.; Hargest, R. Targeting Hyaluronic Acid and Peritoneal Dissemination in Colorectal Cancer. Clin. Color. Cancer 2022, 21, e126–e134. [Google Scholar] [CrossRef]
- Tomonobu, N.; Kinoshita, R.; Sakaguchi, M. S100 Soil Sensor Receptors and Molecular Targeting Therapy Against Them in Cancer Metastasis. Transl. Oncol. 2020, 13, 100753. [Google Scholar] [CrossRef]
- von Lersner, A.; Droesen, L.; Zijlstra, A. Modulation of cell adhesion and migration through regulation of the immunoglobulin superfamily member ALCAM/CD166. Clin. Exp. Metastasis 2019, 36, 87–95. [Google Scholar] [CrossRef]
- Bowen, M.A.; Bajorath, J.; D'Egidio, M.; Whitney, G.S.; Palmer, D.; Kobarg, J.; Starling, G.C.; Siadak, A.W.; Aruffo, A. Characterization of mouse ALCAM (CD166): The CD6-binding domain is conserved in different homologs and mediates cross-species binding. Eur. J. Immunol. 1997, 27, 1469–1478. [Google Scholar] [CrossRef]
- Dippel, V.; Milde-Langosch, K.; Wicklein, D.; Schumacher, U.; Altevogt, P.; Oliveira-Ferrer, L.; Janicke, F.; Schroder, C. Influence of L1-CAM expression of breast cancer cells on adhesion to endothelial cells. J. Cancer Res. Clin. Oncol. 2013, 139, 107–121. [Google Scholar] [CrossRef]
- van Kempen, L.C.; Nelissen, J.M.; Degen, W.G.; Torensma, R.; Weidle, U.H.; Bloemers, H.P.; Figdor, C.G.; Swart, G.W. Molecular basis for the homophilic activated leukocyte cell adhesion molecule (ALCAM)-ALCAM interaction. J. Biol. Chem. 2001, 276, 25783–25790. [Google Scholar] [CrossRef]
- Yang, Y.; Sanders, A.J.; Ruge, F.; Dong, X.; Cui, Y.; Dou, Q.P.; Jia, S.; Hao, C.; Ji, J.; Jiang, W.G. Activated leukocyte cell adhesion molecule (ALCAM)/CD166 in pancreatic cancer, a pivotal link to clinical outcome and vascular embolism. Am. J. Cancer Res. 2021, 11, 5917–5932. [Google Scholar] [PubMed]
- Munsterberg, J.; Loreth, D.; Brylka, L.; Werner, S.; Karbanova, J.; Gandrass, M.; Schneegans, S.; Besler, K.; Hamester, F.; Robador, J.R.; et al. ALCAM contributes to brain metastasis formation in non-small-cell lung cancer through interaction with the vascular endothelium. Neuro Oncol. 2020, 22, 955–966. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.M.; Ferragut, F.; Cardenas Delgado, V.M.; Bracalente, C.; Bravo, A.I.; Cagnoni, A.J.; Nunez, M.; Morosi, L.G.; Quinta, H.R.; Espelt, M.V.; et al. Glycosylation-dependent binding of galectin-8 to activated leukocyte cell adhesion molecule (ALCAM/CD166) promotes its surface segregation on breast cancer cells. Biochim. Biophys. Acta 2016, 1860, 2255–2268. [Google Scholar] [CrossRef] [PubMed]
- Inaguma, S.; Lasota, J.; Wang, Z.; Czapiewski, P.; Langfort, R.; Rys, J.; Szpor, J.; Waloszczyk, P.; Okon, K.; Biernat, W.; et al. Expression of ALCAM (CD166) and PD-L1 (CD274) independently predicts shorter survival in malignant pleural mesothelioma. Hum. Pathol. 2018, 71, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, F.; Murakami, H.; Mizuno, T.; Fujii, M.; Kondo, Y.; Usami, N.; Yokoi, K.; Osada, H.; Sekido, Y. Activated leukocyte cell-adhesion molecule (ALCAM) promotes malignant phenotypes of malignant mesothelioma. J. Thorac. Oncol. 2012, 7, 890–899. [Google Scholar] [CrossRef] [Green Version]
- Cardenes, B.; Clares, I.; Bezos, T.; Toribio, V.; Lopez-Martin, S.; Rocha, A.; Peinado, H.; Yanez-Mo, M.; Cabanas, C. ALCAM/CD166 Is Involved in the Binding and Uptake of Cancer-Derived Extracellular Vesicles. Int. J. Mol. Sci. 2022, 23, 5753. [Google Scholar] [CrossRef]
- Sodek, K.L.; Murphy, K.J.; Brown, T.J.; Ringuette, M.J. Cell-cell and cell-matrix dynamics in intraperitoneal cancer metastasis. Cancer Metastasis Rev. 2012, 31, 397–414. [Google Scholar] [CrossRef] [Green Version]
- Mikula-Pietrasik, J.; Uruski, P.; Tykarski, A.; Ksiazek, K. The peritoneal “soil” for a cancerous “seed”: A comprehensive review of the pathogenesis of intraperitoneal cancer metastases. Cell Mol. Life Sci. 2018, 75, 509–525. [Google Scholar] [CrossRef] [Green Version]
- Ng, D.; Ali, A.; Lee, K.; Eymael, D.; Abe, K.; Luu, S.; Kazazian, K.; Lu, Y.Q.; Brar, S.; Conner, J.; et al. Investigating the mechanisms of peritoneal metastasis in gastric adenocarcinoma using a novel ex vivo peritoneal explant model. Sci. Rep. 2022, 12, 11499. [Google Scholar] [CrossRef]
- Avula, L.R.; Hagerty, B.; Alewine, C. Molecular mediators of peritoneal metastasis in pancreatic cancer. Cancer Metastasis Rev. 2020, 39, 1223–1243. [Google Scholar] [CrossRef]
- Ishigami, S.; Ueno, S.; Arigami, T.; Arima, H.; Uchikado, Y.; Kita, Y.; Sasaki, K.; Nishizono, Y.; Omoto, I.; Kurahara, H.; et al. Clinical implication of CD166 expression in gastric cancer. J. Surg. Oncol. 2011, 103, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Qian, C.; Jing, L.; Ren, J.; Guan, Y. Meta-analysis indicating that high ALCAM expression predicts poor prognosis in colorectal cancer. Oncotarget 2017, 8, 48272–48281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, L.; Russo, L.; Ulugoel, S.; Freire Dos Santos, R.; Breuer, E.; Gupta, A.; Lehmann, K. Peritoneal Metastasis: Current Status and Treatment Options. Cancers 2021, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Wiiger, M.T.; Gehrken, H.B.; Fodstad, O.; Maelandsmo, G.M.; Andersson, Y. A novel human recombinant single-chain antibody targeting CD166/ALCAM inhibits cancer cell invasion in vitro and in vivo tumour growth. Cancer Immunol. Immunother. 2010, 59, 1665–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, R.; Sato, H.; Yamauchi, A.; Takahashi, Y.; Inoue, Y.; Sumardika, I.W.; Chen, Y.; Tomonobu, N.; Araki, K.; Shien, K.; et al. exSSSRs (extracellular S100 soil sensor receptors)-Fc fusion proteins work as prominent decoys to S100A8/A9-induced lung tropic cancer metastasis. Int. J. Cancer 2019, 144, 3138–3145. [Google Scholar] [CrossRef] [PubMed]
- Sanders, A.J.; Owen, S.; Morgan, L.D.; Ruge, F.; Collins, R.J.; Ye, L.; Mason, M.D.; Jiang, W.G. Importance of activated leukocyte cell adhesion molecule (ALCAM) in prostate cancer progression and metastatic dissemination. Oncotarget 2019, 10, 6362–6377. [Google Scholar] [CrossRef] [Green Version]
- Bartolomé, R.A.; Pintado-Berninches, L.; Jaén, M.; de Los Ríos, V.; Imbaud, J.I.; Casal, J.I. SOSTDC1 promotes invasion and liver metastasis in colorectal cancer via interaction with ALCAM/CD166. Oncogene 2020, 39, 6085–6098. [Google Scholar] [CrossRef]
- Antoine-Bertrand, J.; Ghogha, A.; Luangrath, V.; Bedford, F.K.; Lamarche-Vane, N. The activation of ezrin-radixin-moesin proteins is regulated by netrin-1 through Src kinase and RhoA/Rho kinase activities and mediates netrin-1-induced axon outgrowth. Mol. Biol. Cell 2011, 22, 3734–3746. [Google Scholar] [CrossRef]
- Tudor, C.; te Riet, J.; Eich, C.; Harkes, R.; Smisdom, N.; Bouhuijzen, W.J.; Ameloot, M.; Holt, M.; Kanger, J.S.; Figdor, C.G.; et al. Syntenin-1 and ezrin proteins link activated leukocyte cell adhesionmolecule to the actin cytoskeleton. J. Biol. Chem. 2014, 289, 13445–13460. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Ye, L.; Ji, K.; Zhang, L.; Hargest, R.; Ji, J.; Jiang, W.G. Death-associated protein-3, DAP-3, correlates with preoperative chemotherapy effectiveness and prognosis of gastric cancer patients following perioperative chemotherapy and radical gastrectomy. Br. J. Cancer 2014, 110, 421–429. [Google Scholar] [CrossRef]
- Ji, J.; Jia, S.; Jia, Y.; Ji, K.; Hargest, R.; Jiang, W.G. WISP-2 in human gastric cancer and its potential metastatic suppressor role in gastric cancer cells mediated by JNK and PLC-gamma pathways. Br. J. Cancer 2015, 113, 921–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiscox, S.; Jiang, W.G. Quantification of tumour cell-endothelial cell attachment by 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine (DiI). Cancer Lett. 1997, 112, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Keese, C.R.; Bhawe, K.; Wegener, J.; Giaever, I. Real-time impedance assay to follow the invasive activities of metastatic cells in culture. Biotechniques 2002, 33, 842–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.G.; Ablin, R.J.; Kynaston, H.G.; Mason, M.D. The prostate transglutaminase (TGase-4, TGaseP) regulates the interaction of prostate cancer and vascular endothelial cells, a potential role for the ROCK pathway. Microvasc. Res. 2009, 77, 150–157. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HR | P * | Survival Time | |
|---|---|---|---|
| Gastric cancer | 8.79 | 0.003 | 92.8 ± 2.3 vs. 80.2 ± 4.5 Months |
| Pancreatic cancer | 2.84 | 0.208 | 93.1 ± 11.3 vs. 66.5 ± 15.3 Months |
| Target | Forward Primer | Reverse Primer * |
|---|---|---|
| ALCAM | ttatcataccttgccgatt | gggtggaagtcatggtatag |
| ALCAM | caggaggttgaaggactaaa | actgaacctgaccgtacagggatcagttttctttgtca |
| CD6 | ctactgcggccacaaag | actgaacctgaccgtacactcggaagtgtacctcca |
| L1CAM | ccacttgtttaaggagagga | actgaacctgaccgtacagatgatggcactcacaaag |
| SRC | tgtggccctctatgactatg | aaactccccttgctcatgta |
| Ezrin | tggagagagagaaagagcag | ttcttctctgcctcagtgat |
| Moesin | taagaaggctcagcaagaac | cttcttggactcatctctgg |
| GAPDH | ggctgcttttaactctggta | gactgtggtcatgagtcctt |
| GAPDH | aaggtcatccatgacaactt | actgaacctgaccgtacagccatccacagtcttctg |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.M.; Ye, L.; Ruge, F.; Fang, Z.; Ji, K.; Sanders, A.J.; Jia, S.; Hao, C.; Dou, Q.P.; Ji, J.; et al. Activated Leukocyte Cell Adhesion Molecule (ALCAM), a Potential ‘Seed’ and ‘Soil’ Receptor in the Peritoneal Metastasis of Gastrointestinal Cancers. Int. J. Mol. Sci. 2023, 24, 876. https://doi.org/10.3390/ijms24010876
Yang YM, Ye L, Ruge F, Fang Z, Ji K, Sanders AJ, Jia S, Hao C, Dou QP, Ji J, et al. Activated Leukocyte Cell Adhesion Molecule (ALCAM), a Potential ‘Seed’ and ‘Soil’ Receptor in the Peritoneal Metastasis of Gastrointestinal Cancers. International Journal of Molecular Sciences. 2023; 24(1):876. https://doi.org/10.3390/ijms24010876
Chicago/Turabian StyleYang, Yi Ming, Lin Ye, Fiona Ruge, Ziqian Fang, Ke Ji, Andrew J. Sanders, Shuqin Jia, Chunyi Hao, Q. Ping Dou, Jiafu Ji, and et al. 2023. "Activated Leukocyte Cell Adhesion Molecule (ALCAM), a Potential ‘Seed’ and ‘Soil’ Receptor in the Peritoneal Metastasis of Gastrointestinal Cancers" International Journal of Molecular Sciences 24, no. 1: 876. https://doi.org/10.3390/ijms24010876