Deletion of Gb3 Synthase in Mice Resulted in the Attenuation of Bone Formation via Decrease in Osteoblasts

 , ,
, ,
Abstract
:1. Introduction
2. Results
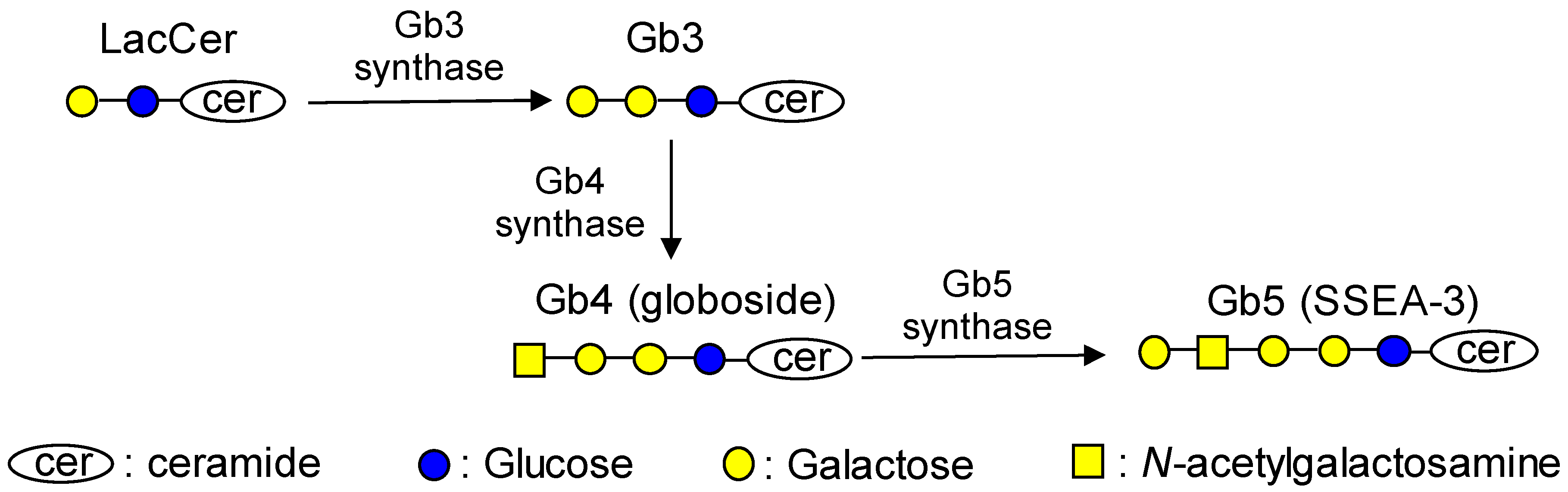
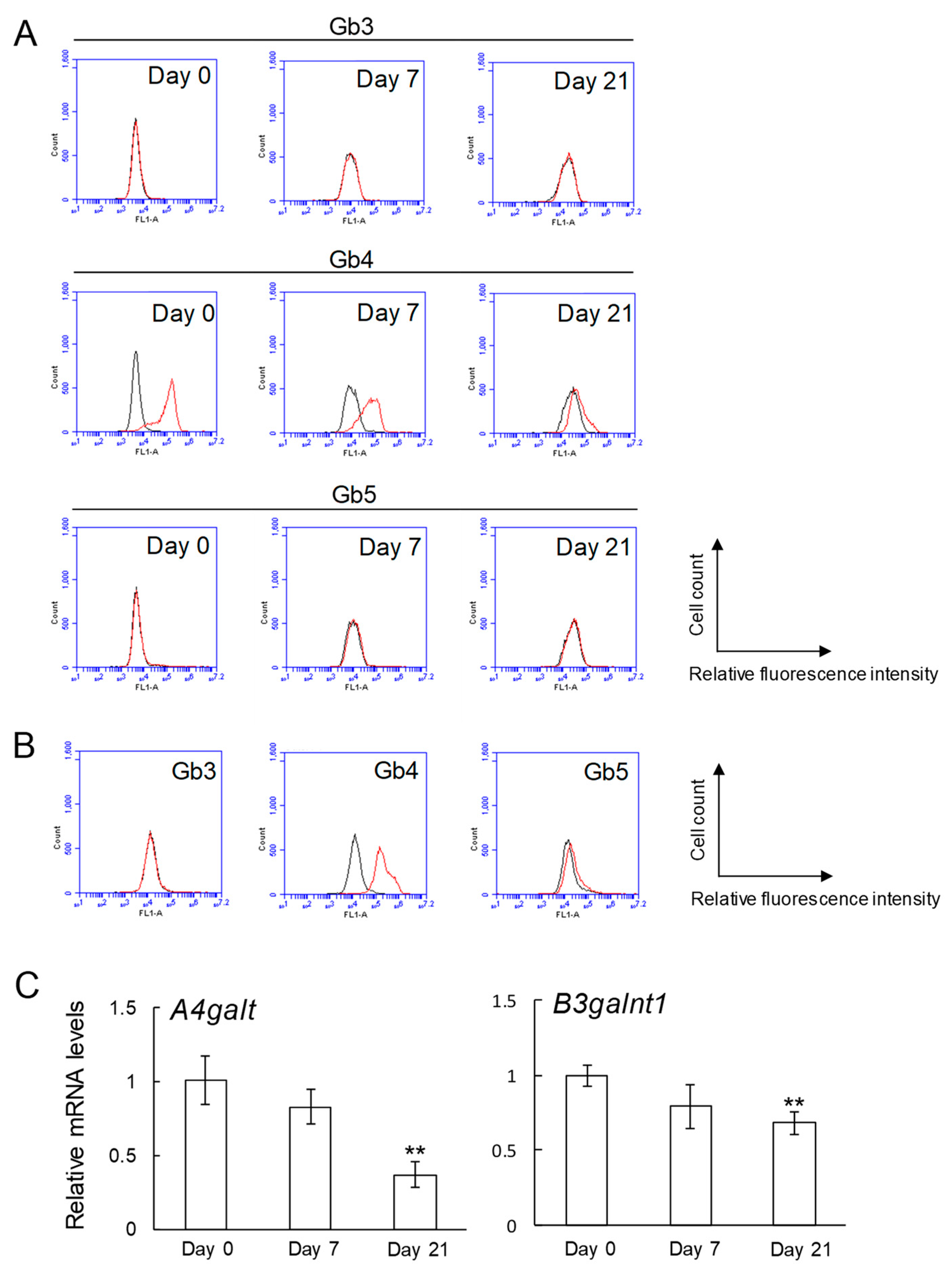
2.1. Expression of Globo-Series Glycosphingolipids (Gb3, Gb4, and Gb5), and Gb3 and Gb4 Synthase Genes in Osteoblasts
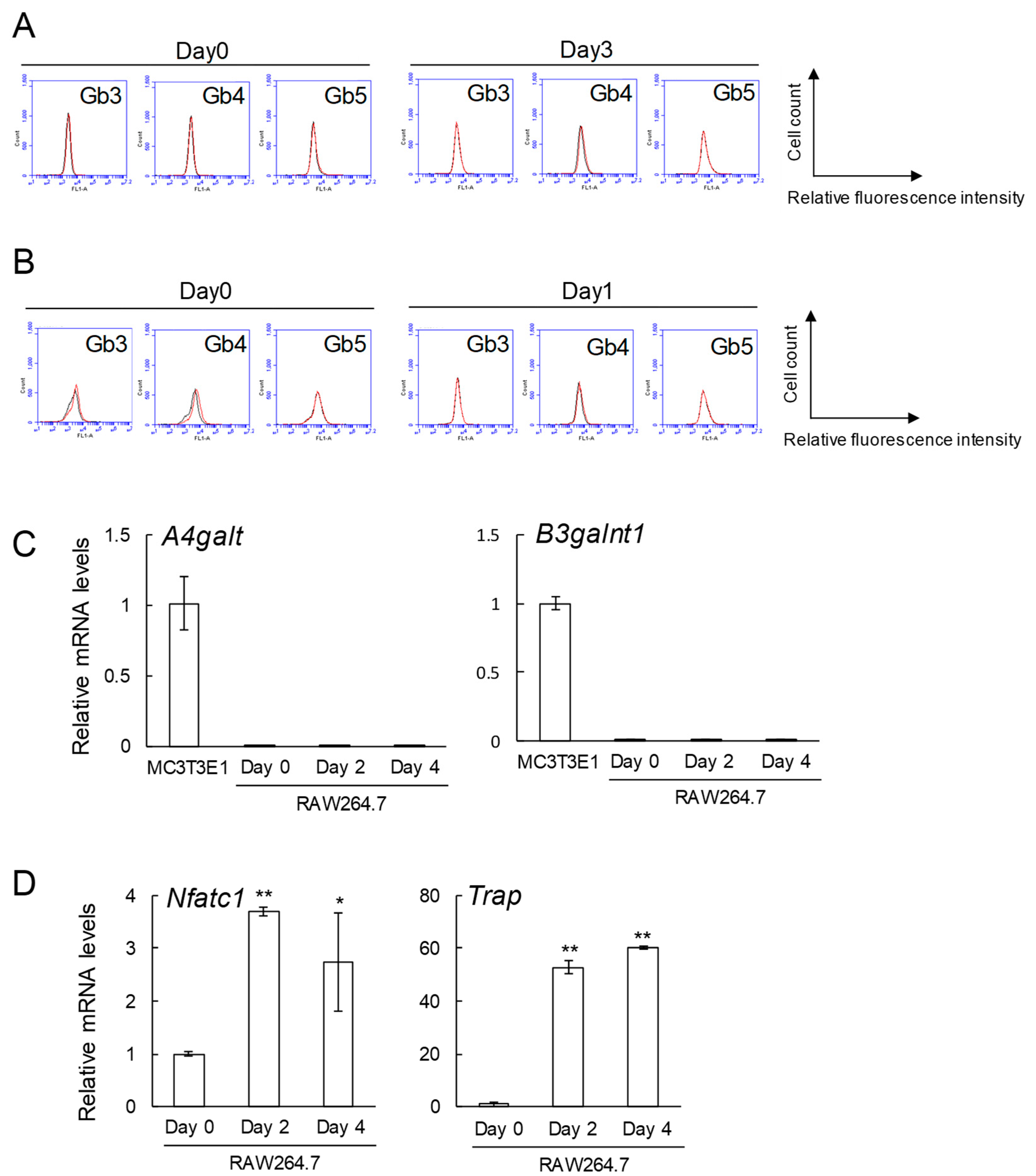
2.2. No Expression of Globo-Series Glycosphingolipids (Gb3, Gb4, and Gb5), and Gb3 and Gb4 Synthase Genes in RAW264.7 Cells and Primary Cultured Pre-Osteoclasts
2.3. Decrease in Femoral Cancellous Bone Mass in Gb3 Synthase-Knockout (Gb3S KO) Mice
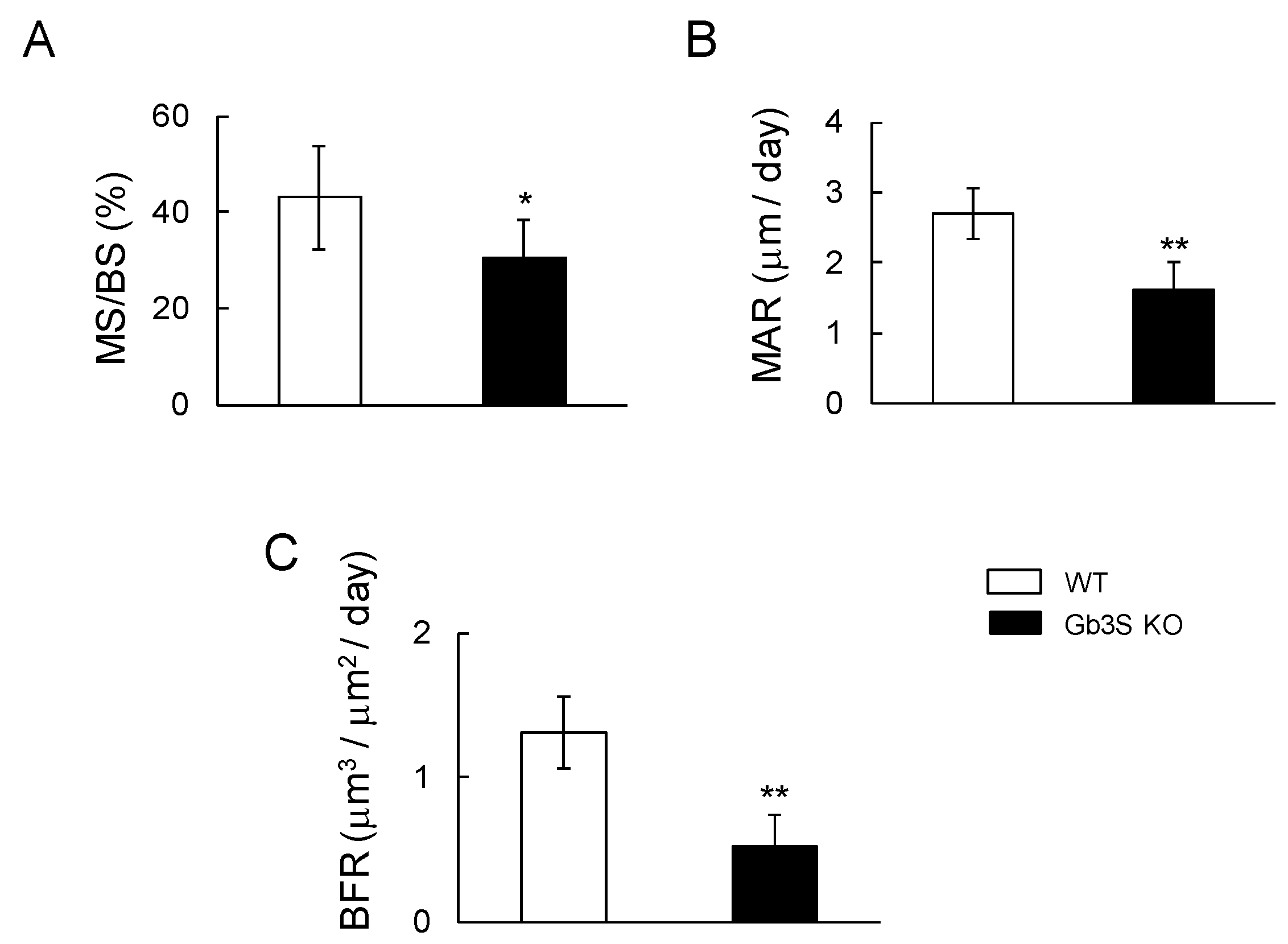
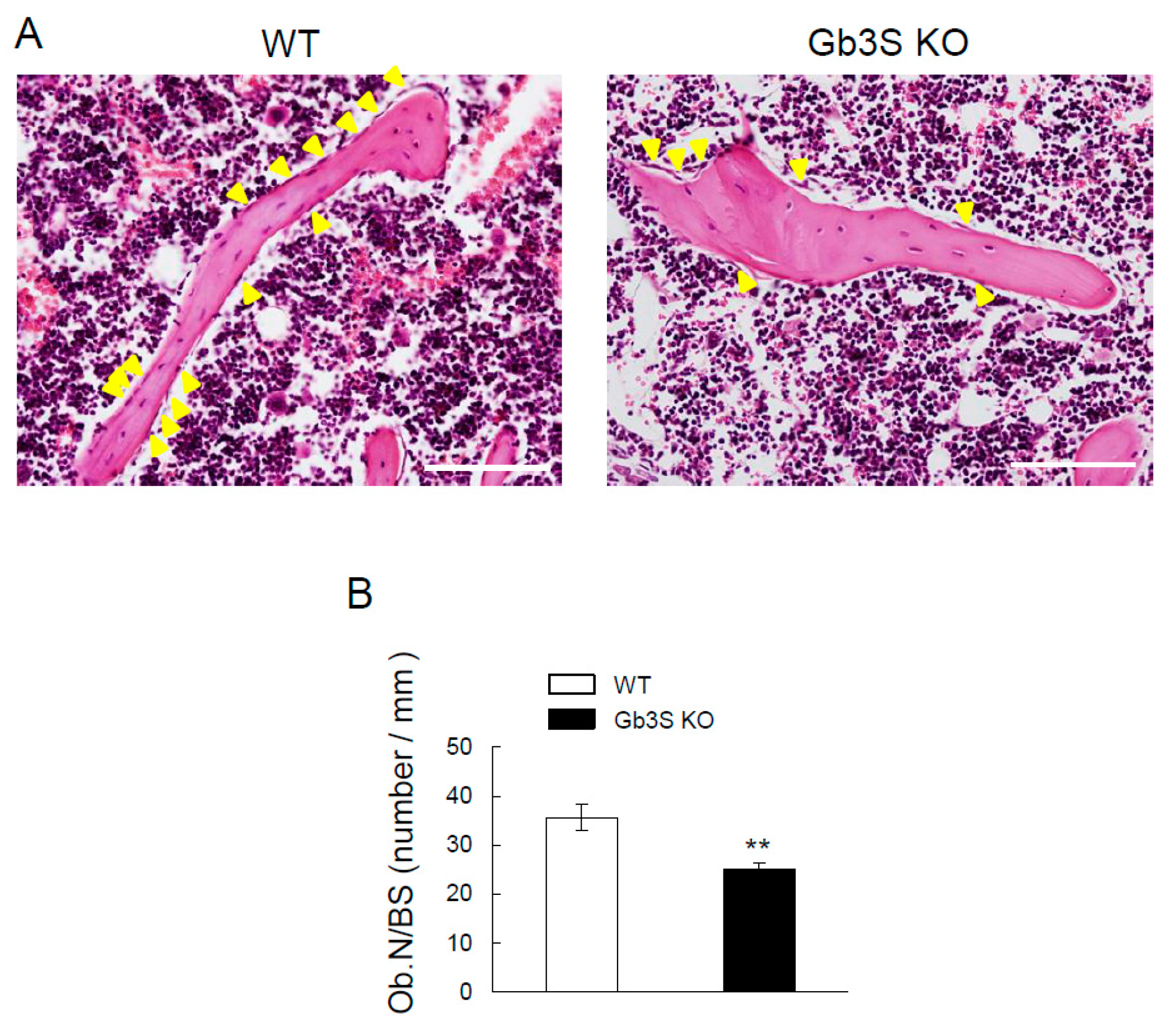
2.4. Decrease in Bone Formation Parameters by Gb3 Synthase Deficiency
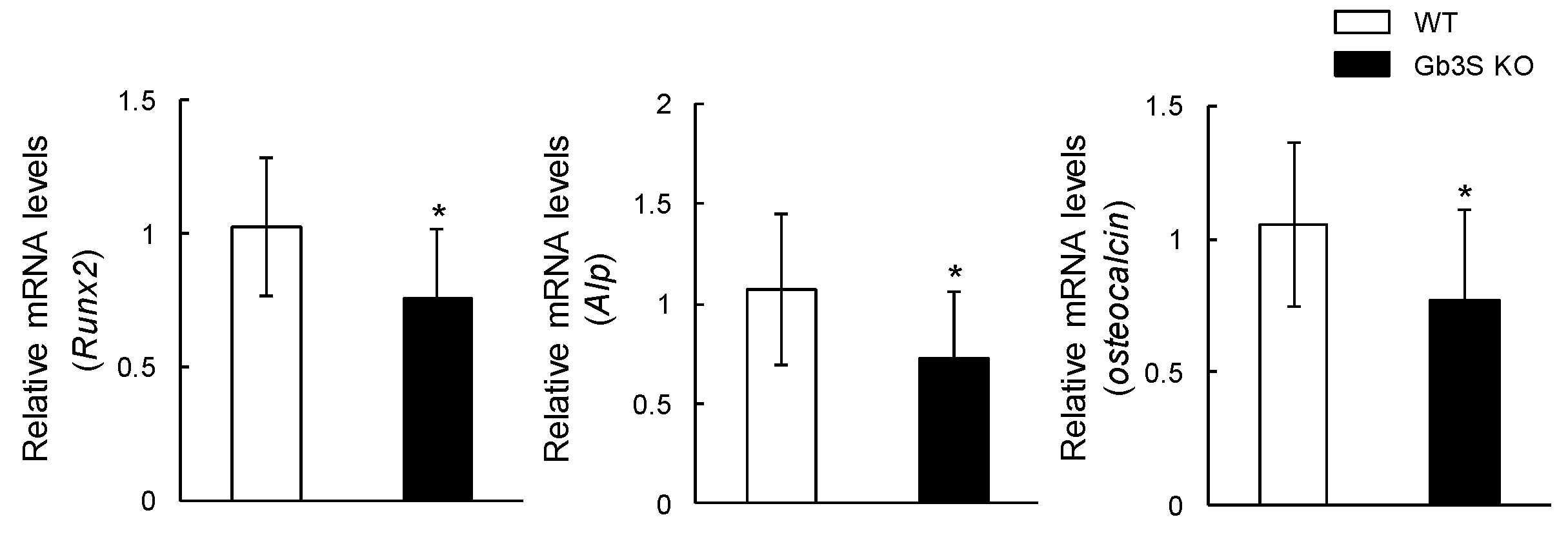
2.5. Decrease in Osteoblast Number in Femoral Cancellous Bone and Osteogenic Differentiation Marker’s Gene Expression (Runt-Related Transcription Factor 2, Alkaline Phosphatase, and Osteocalcin) in Long Bones by Gb3 Synthase Deficiency
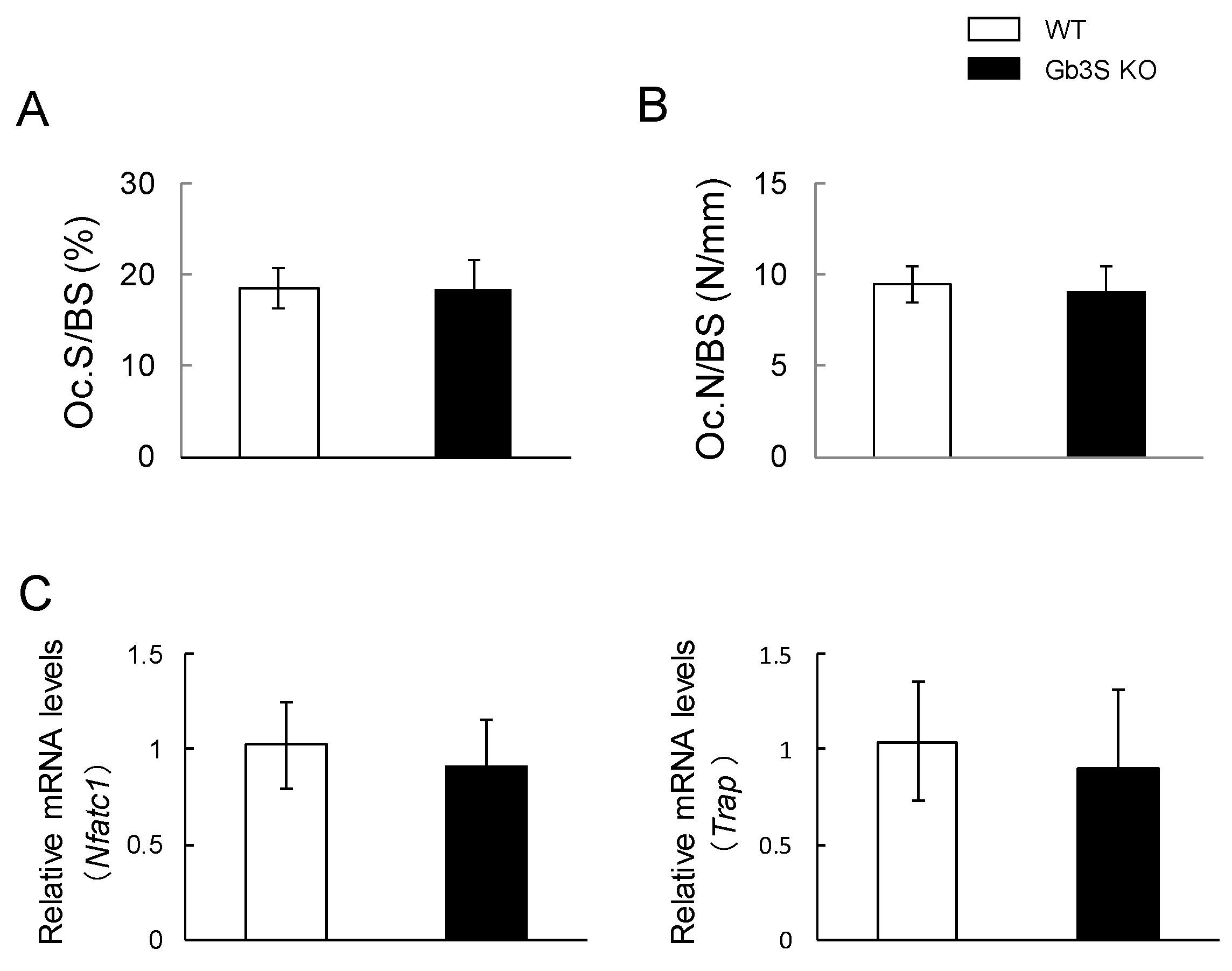
2.6. No Differences in Osteoclast Number and Expression of Osteoclast Differentiation Marker’s Genes (Nuclear Factor of Activated T-Cells, Cytoplasmic 1 and Tartrate-Resistant Acid Phosphatase) between WT and Gb3S KO Mice
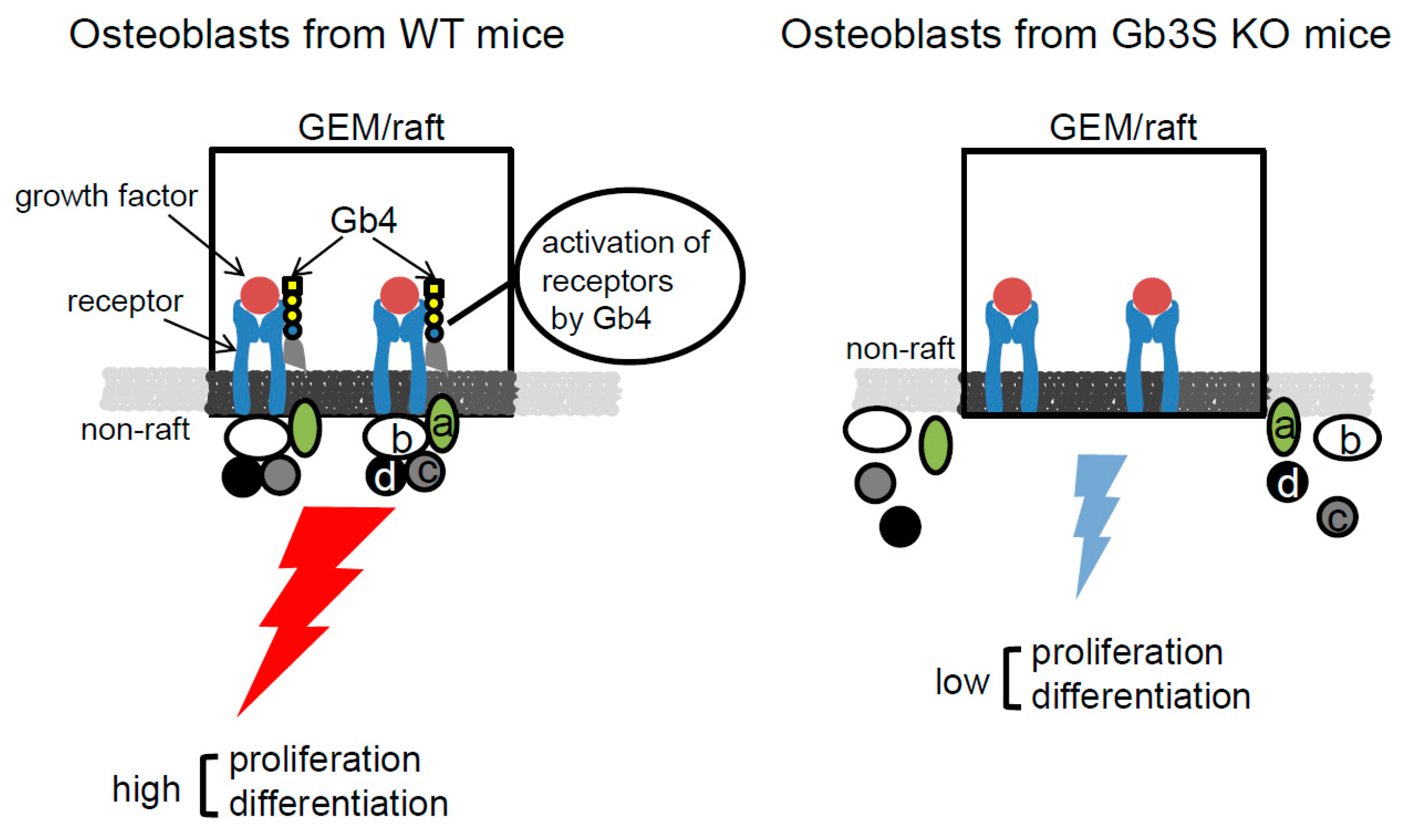
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Cell Culture
4.3. Antibodies
4.4. Induction to Mature Osteoblasts
4.5. In Vitro Osteoclast Induction
4.6. Flow Cytometry
4.7. Quantitative Real-Time PCR (qPCR)
4.8. Three-Dimensional Micro-Computed Tomography (3D-μCT Analysis)
4.9. Measurement of Bone Formation Rate Using Calcein Double Labeling
4.10. TRAP Staining
4.11. Bone Histomorphometric Analyses
4.12. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yu, R.K.; Tsai, Y.T.; Ariga, T. Functional roles of gangliosides in neurodevelopment: An overview of recent advances. Neurochem. Res. 2012, 37, 1230–1244. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Ohmi, Y.; Ohkawa, Y.; Tajima, O.; Furukawa, K. Glycosphingolipids in the regulation of the nerve system. Adv. Neurobiol. 2014, 9, 307–320. [Google Scholar] [PubMed]
- Furukawa, K.; Ohmi, Y.; Ji, S.; Zhang, P.; Bhuiyan, R.H.; Ohkawa, Y.; Tajima, O.; Hashimoto, N.; Furukawa, K. Glycolipids: Essential regulator of neuro-inflammation, metabolism and gliomagenesis. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 2479–2484. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Ohmi, Y.; Kondo, Y.; Bhuiyan, R.H.; Tajima, O.; Zhang, P.; Ohkawa, Y.; Furukawa, K. Elucidation of the enigma of glycosphingolipids in the regulation of inflammation and degeneration. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2019, 95, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Jung, J.U.; Ryu, J.S.; Jin, J.W.; Yang, H.J.; Ko, K.; You, H.K.; Jung, K.Y.; Choo, Y.K. Effects of gangliosides on the differentiation of human mesenchymal stem cells into osteoblasts by modulating epidermal growth factor receptors. Biochem. Biophys. Res. Commun. 2008, 371, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.J.; Jung, K.Y.; Kwak, D.H.; Lee, S.H.; Ryu, J.S.; Kim, J.S.; Chang, K.T.; Lee, J.W.; Choo, Y.K. Inhibition of ganglioside GD1a synthesis suppresses the differentiation of human mesenchymal stem cells into osteoblasts. Dev. Growth Differ. 2011, 53, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Bergante, S.; Torretta, E.; Creo, P.; Sessarego, N.; Papini, N.; Piccoli, M.; Fania, C.; Cirillo, F.; Conforti, E.; Ghiroldi, A.; et al. Gangliosides as a potential new class of stem cell markers: The case of GD1a in human bone marrow mesenchymal stem cells. J. Lipid Res. 2014, 55, 549–560. [Google Scholar] [CrossRef]
- Bergante, S.; Creo, P.; Piccoli, M.; Ghiroldi, A.; Menon, A.; Cirillo, F.; Rota, P.; Monasky, M.M.; Ciconte, G.; Pappone, C.; et al. GM1 ganglioside promotes osteogenic differentiation of human tendon stem cells. Stem Cells Int. 2018, 4706943. [Google Scholar] [CrossRef]
- Yo, S.; Hamamura, K.; Mishima, Y.; Hamajima, K.; Mori, H.; Furukawa, K.; Kondo, H.; Tanaka, K.; Sato, T.; Miyazawa, K.; et al. Deficiency of GD3 synthase in mice resulting in the attenuation of bone loss with aging. Int. J. Mol. Sci. 2019, 20, 2825. [Google Scholar] [CrossRef]
- Wiels, J.; Fellous, M.; Tursz, T. Monoclonal antibody against a Burkitt lymphoma-associated antigen. Proc. Natl. Acad. Sci. USA 1981, 78, 6485–6488. [Google Scholar] [CrossRef]
- Kannagi, R.; Cochran, N.A.; Ishigami, F.; Hakomori, S.; Andrews, P.W.; Knowles, B.B.; Solter, D. Stage-specific embryonic antigens (SSEA-3 and -4) are epitopes of a unique globo-series ganglioside isolated from human teratocarcinoma cells. EMBO J. 1983, 2, 2355–2361. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.J.; Andrews, P.W. Surface marker antigens in the characterization of human embryonic stem cells. Stem Cell Res. 2009, 3, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, Y.; Ikeda, K.; Tokuda, N.; Nishitani, C.; Ohto, U.; Akashi-Takamura, S.; Ito, Y.; Uchikawa, M.; Kuroki, Y.; Taguchi, R.; et al. TLR4-MD-2 complex is negatively regulated by an endogenous ligand, globotetraosylceramide. Proc. Natl. Acad. Sci. USA 2013, 110, 4714–4719. [Google Scholar] [CrossRef] [PubMed]
- Marcus, D.M.; Kundu, S.K.; Suzuki, A. The P blood group system: Recent progress in immunochemistry and genetics. Semin. Hematol. 1981, 18, 63–71. [Google Scholar] [PubMed]
- Jacewicz, M.; Clausen, H.; Nudelman, E.; Donohue-Rolfe, A.; Keusch, G.T. Pathogenesis of shigella diarrhea. XI. Isolation of a shigella toxin-binding glycolipid from rabbit jejunum and Hela cells and its identification as globotriaosylceramide. J. Exp. Med. 1986, 163, 1391–1404. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, T.; Iida, T. Immunochemical study on the red blood cells. I. Globoside, as the agglutinogen of the ABO system on erythrocytes. Jpn. J. Exp. Med. 1953, 23, 327–331. [Google Scholar] [PubMed]
- Kijimoto-Ochiai, S.; Naiki, M.; Makita, A. Defects of glycosyltransferase activities in human fibroblasts of Pk and p blood group phenotypes. Proc. Natl. Acad. Sci. USA 1977, 74, 5407–5410. [Google Scholar] [CrossRef]
- Park, S.Y.; Kwak, C.Y.; Shayman, J.A.; Kim, J.H. Globoside promotes activation of ERK by interaction with the epidermal growth factor receptor. Biochim. Biophys. Acta Gen. Subj. 2012, 1820, 1141–1148. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.K.; Ariga, T.; Yoshino, H.; Katoh-Semba, R.; Ren, S. Differential effects of glycosphingolipids n protein kinase C activity in PC12D pheochromocytoma cells. J. Biomed. Sci. 1994, 1, 229–236. [Google Scholar] [CrossRef]
- Song, Y.; Withers, D.A.; Hakomoro, S.I. Globoside-dependent adhesion of human embryonal carcinoma cells, based on carbohydrate-carbohydrate interaction, initiates signal transduction and induces enhanced activity of transcription factor AP1 and CREB. J. Biol. Chem. 1998, 273, 2517–2525. [Google Scholar] [CrossRef]
- Nakamura, T.; Chiba, Y.; Naruse, M.; Saito, K.; Harada, H.; Fukumoto, S. Globoside accelerates the differentiation of dental epithelial cells into ameloblasts. Int. J. Oral Sci. 2016, 8, 205–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.Y.; Gupta, V.; Patwardhan, G.A.; Bhinge, K.; Zhao, Y.; Bao, J.; Mehendale, H.; Cabot, M.C.; Li, Y.T.; Jazwinski, S.M. Glucosylceramide synthase upregulates MDR1 expression in the regulation of cancer drug resistance through cSrc and β-catenin signaling. Mol. Cancer 2010, 9, 145. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.F.; Chaudhary, L.; Fausto, A.; Halstead, L.R.; Ory, D.S.; Avioli, L.V.; Cheng, S.L. Erk is essential for growth, differentiation, integrin expression, and cell function in human osteoblastic cells. J. Biol. Chem. 2001, 276, 14443–14450. [Google Scholar] [CrossRef] [PubMed]
- Nakura, A.; Higuchi, C.; Yoshida, K.; Yoshikawa, H. PKCα suppresses osteoblastic differentiation. Bone 2011, 48, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Westendorf, J.J.; Kahler, R.A.; Schroeder, T.M. Wnt signaling in osteoblasts and bone diseases. Gene 2004, 341, 19–39. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Choi, Y.; Yoon, D.S.; Lee, K.M.; Kim, D.; Lee, J.W. Zinc promotes osteoblast differentiation in human mesenchymal stem cells via activation of the cAMP-PKA-CREB signaling pathway. Stem Cells Dev. 2018, 27, 1125–1135. [Google Scholar] [CrossRef]
- Hakomori, S. Glycosynapses: Microdomains controlling carbohydrate-dependent cell adhesion and signaling. Acad. Bras. Cienc. 2004, 76, 553–572. [Google Scholar] [CrossRef]
- Furukawa, K.; Ohkawa, Y.; Yamauchi, Y.; Hamamura, K.; Ohmi, Y.; Furukawa, K. Fine tuning of cell signals by glycosylation. J. Biochem. 2012, 151, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Ikeda, K.; Hamamura, K.; Zhang, Q.; Kondo, Y.; Matsumoto, Y.; Ohmi, Y.; Yamauchi, Y.; Furukawa, K.; Taguchi, R.; et al. GM1/GD1b/GA1 synthase expression results in the reduced cancer phenotypes with modulation of composition and raft-localization of gangliosides in a melanoma cell line. Cancer Sci. 2010, 101, 2039–2047. [Google Scholar] [CrossRef]
- Hamamura, K.; Tsuji, M.; Hotta, H.; Ohkawa, Y.; Takahashi, M.; Shibuya, H.; Nakashima, H.; Yamauchi, Y.; Hashimoto, N.; Hattori, H.; et al. Functional activation of Src family kinase Yes protein is essential for the enhanced malignant properties of human melanoma cells expressing ganglioside GD3. J. Biol. Chem. 2011, 286, 18526–18537. [Google Scholar] [CrossRef]
- Furukawa, K.; Ohmi, Y.; Ohkawa, Y.; Bhuiyan, R.H.; Zhang, P.; Tajima, O.; Hashimoto, N.; Hamamura, K.; Furukawa, K. New era of research on cancer-associated glycosphingolipids. Cancer Sci. 2019, 110, 1544–1551. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Tokuda, N.; Numata, S.; Ito, M.; Ohta, M.; Kawamura, K.; Wiels, J.; Urano, T.; Tajima, O.; Furukawa, K.; et al. Targeted disruption of Gb3/CD77 synthase gene resulted in the complete deletion of globo-series glycosphingolipids and loss of sensitivity to verotoxins. J. Biol. Chem. 2006, 281, 10230–10235. [Google Scholar] [CrossRef] [PubMed]
- Koshihara, Y.; Kawamura, M.; Endo, S.; Tsutsumi, C.; Kodama, H.; Oda, H.; Higaki, S. Establishment of human osteoblastic cells derived from periosteum in culture. In Vitro Cell. Dev. Biol. 1989, 25, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Hamamura, K.; Chen, A.; Tanjung, N.; Takigawa, S.; Sudo, A.; Yokota, H. In vitro and in silico analysis of an inhibitory mechanism of osteoclastogenesis by salubirnal and guanabenz. Cell. Signal. 2015, 27, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y.; Tokuda, N.; Furukawa, K.; Ando, R.; Uchikawa, M.; Zhang, Q.; Xiaoyan, F.; Furukawa, K. Efficient generation of useful monoclonal antibodies reactive with globotriaosylceramide using knockout mice lacking Gb3/CD77 synthase. Glycoconj. J. 2011, 28, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Iwamura, K.; Furukawa, K.; Uchikawa, M.; Sojka, B.N.; Kojima, Y.; Wiels, J.; Shiku, H.; Urano, T.; Furukawa, K. The blood group P1 synthase gene is identical to the Gb3/CD77 synthase gene. J. Biol. Chem. 2003, 278, 44429–44438. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Kondo, H.; Hamamura, K.; Togari, A. Systemic administration of low-dose naltrexone increases bone mass due to blockade of opioid growth factor receptor signaling in mice osteoblasts. Life Sci. 2019, 224, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Mori, H.; Hamamura, K.; Yo, S.; Hamajima, K.; Ootani, K.; Honda, M.; Ishizuka, K.; Kondo, H.; Tanaka, K.; Kodama, D.; et al. Conditioned medium from rat dental pulp reduces the number of osteoclasts via attenuation of adhesiveness in osteoclast precursors. J. Oral Sci. 2018, 60, 352–359. [Google Scholar] [CrossRef]
- Tanaka, K.; Hirai, T.; Kodama, D.; Kondo, H.; Hamamura, K.; Togari, A. α1B-Adrenoceptor signaling regulates bone formation through the up-regulation of CCAAT/enhancer-binding protein δ expression in osteoblasts. Br. J. Pharmacol. 2016, 173, 1058–1069. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Forward Primer | Backward Primer |
|---|---|---|
| A4galt | 5′-CCTGTTCCCATCTGGAGGAG-3′ | 5′-CCCTTTCATCAGCACAACCA-3′ |
| B3galnt1 | 5′-TCTTGACTGCCCTTCCCAAT-3′ | 5′-GGAGCGTGAAGCGAAAGTCT-3′ |
| Runx2 | 5′-CCCAGCCACCTTTACCTACA-3′ | 5′-TATGGAGTGCTGCTGGTCTG-3′ |
| Alp | 5′-AACCCAGACACAAGCATTCC-3′ | 5′-GCCTTTGAGGTTTTTGGTCA-3′ |
| osteocalcin | 5′-CCGGGAGCAGTGTGAGCTTA-3′ | 5′-AGGCGGTCTTCAAGCCATACT-3′ |
| Nfatc1 | 5′-GGTGCTGTCTGGCCATAACT-3′ | 5′-GCGGAAAGGTGGTATCTCAA-3′ |
| Trap | 5′-TCCTGGCTCAAAAAGCAGTT-3′ | 5′-ACATAGCCCACACCGTTCTC-3′ |
| Gapdh | 5′-TGCACCACCAACTGCTTAG-3′ | 5′-GGATGCAGGGATGATGTTC-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamamura, K.; Hamajima, K.; Yo, S.; Mishima, Y.; Furukawa, K.; Uchikawa, M.; Kondo, Y.; Mori, H.; Kondo, H.; Tanaka, K.; et al. Deletion of Gb3 Synthase in Mice Resulted in the Attenuation of Bone Formation via Decrease in Osteoblasts. Int. J. Mol. Sci. 2019, 20, 4619. https://doi.org/10.3390/ijms20184619
Hamamura K, Hamajima K, Yo S, Mishima Y, Furukawa K, Uchikawa M, Kondo Y, Mori H, Kondo H, Tanaka K, et al. Deletion of Gb3 Synthase in Mice Resulted in the Attenuation of Bone Formation via Decrease in Osteoblasts. International Journal of Molecular Sciences. 2019; 20(18):4619. https://doi.org/10.3390/ijms20184619
Chicago/Turabian StyleHamamura, Kazunori, Kosuke Hamajima, Shoyoku Yo, Yoshitaka Mishima, Koichi Furukawa, Makoto Uchikawa, Yuji Kondo, Hironori Mori, Hisataka Kondo, Kenjiro Tanaka, and et al. 2019. "Deletion of Gb3 Synthase in Mice Resulted in the Attenuation of Bone Formation via Decrease in Osteoblasts" International Journal of Molecular Sciences 20, no. 18: 4619. https://doi.org/10.3390/ijms20184619