Pathogenesis, Early Diagnosis, and Therapeutic Management of Alcoholic Liver Disease


Abstract
:1. Introduction
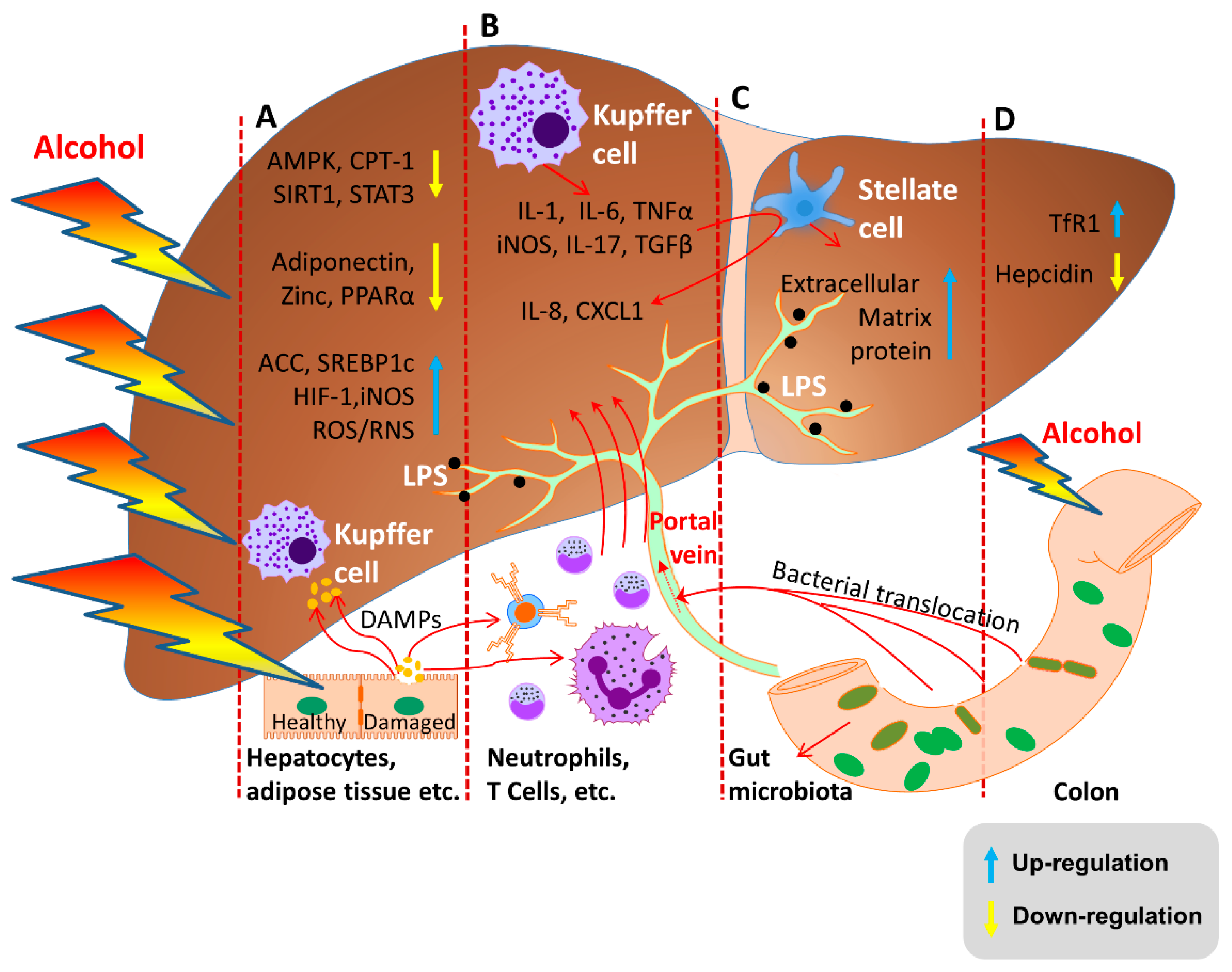
2. Pathogenesis of ALD
2.1. Alcohol Metabolism
2.2. The Spectrum of ALD
2.2.1. Alcoholic Fatty Liver (AFL)/Steatosis
2.2.2. Alcoholic Hepatitis (AH)
2.2.3. Alcoholic Steatohepatitis (ASH)
2.2.4. Alcoholic Liver Fibrosis/Cirrhosis
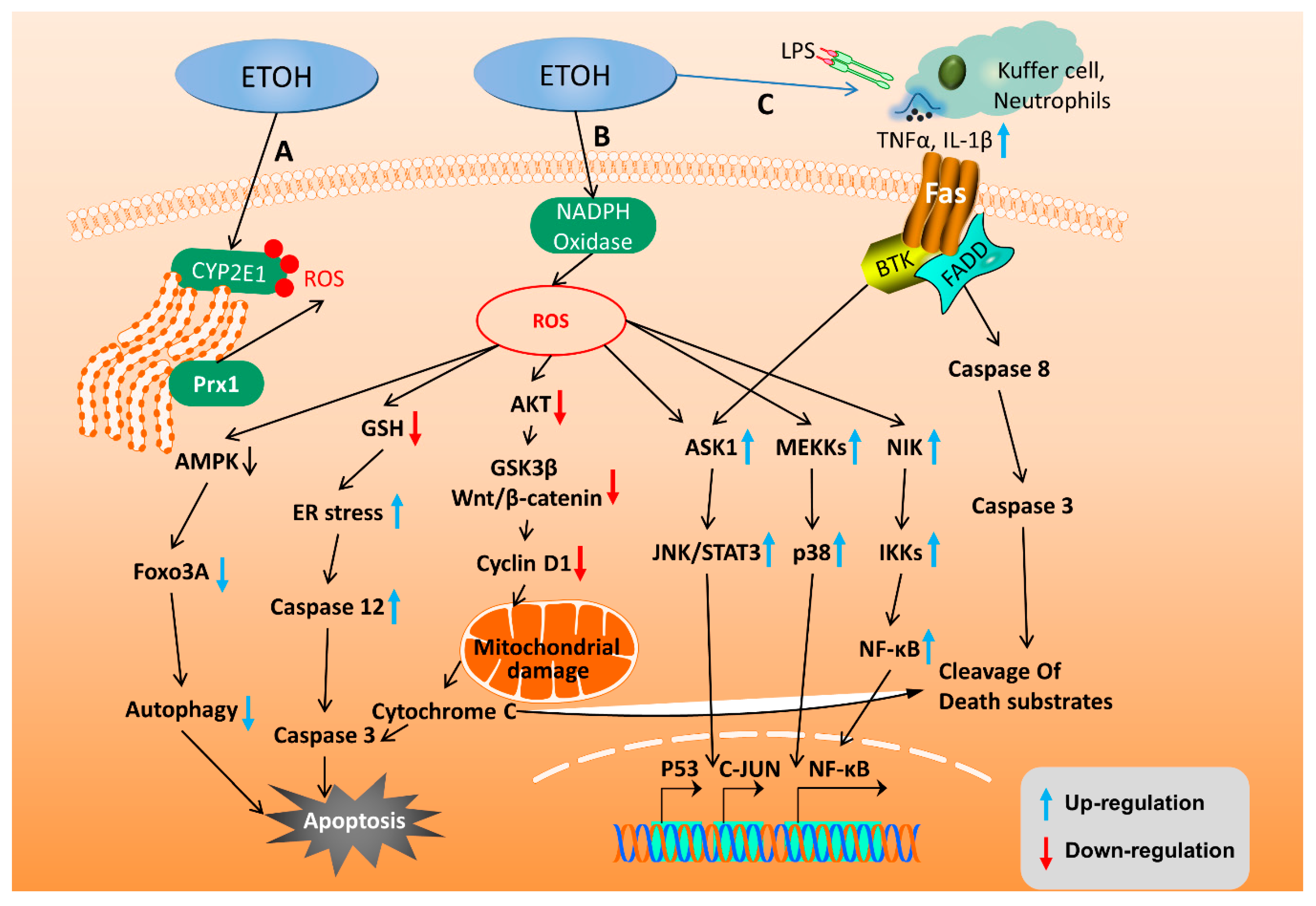
2.3. Molecular Mechanisms of ALD
2.3.1. Alcohol and Its Metabolites Damage the Liver
2.3.2. Oxidative Stress and Lipid Peroxidation
2.3.3. Endotoxin Enteric Leakage
2.3.4. Hepcidin Regulation
2.4. Apoptotic Signaling Pathway and Autophagy in ALD
2.5. Genetic Factors of ALD
3. Early Diagnosis of ALD
3.1. Clinical Symptoms
3.2. Imaging Examination
3.3. Detection of Specific Biomarkers
4. Current Therapies
4.1. Nutritional Therapy
4.2. Alcohol Withdrawal Therapies
4.3. Hormone Related Therapies
4.4. Liver Transplantation
4.5. Targeted Therapies
4.5.1. TNF Receptor Superfamily Target Therapies
4.5.2. Antioxidant Signal Targeting Therapies
4.5.3. Targeting the Inhibition of Hepatocyte Apoptosis
4.5.4. MicroRNAs Targeted Therapies
4.6. Specific Treatment Options for AH
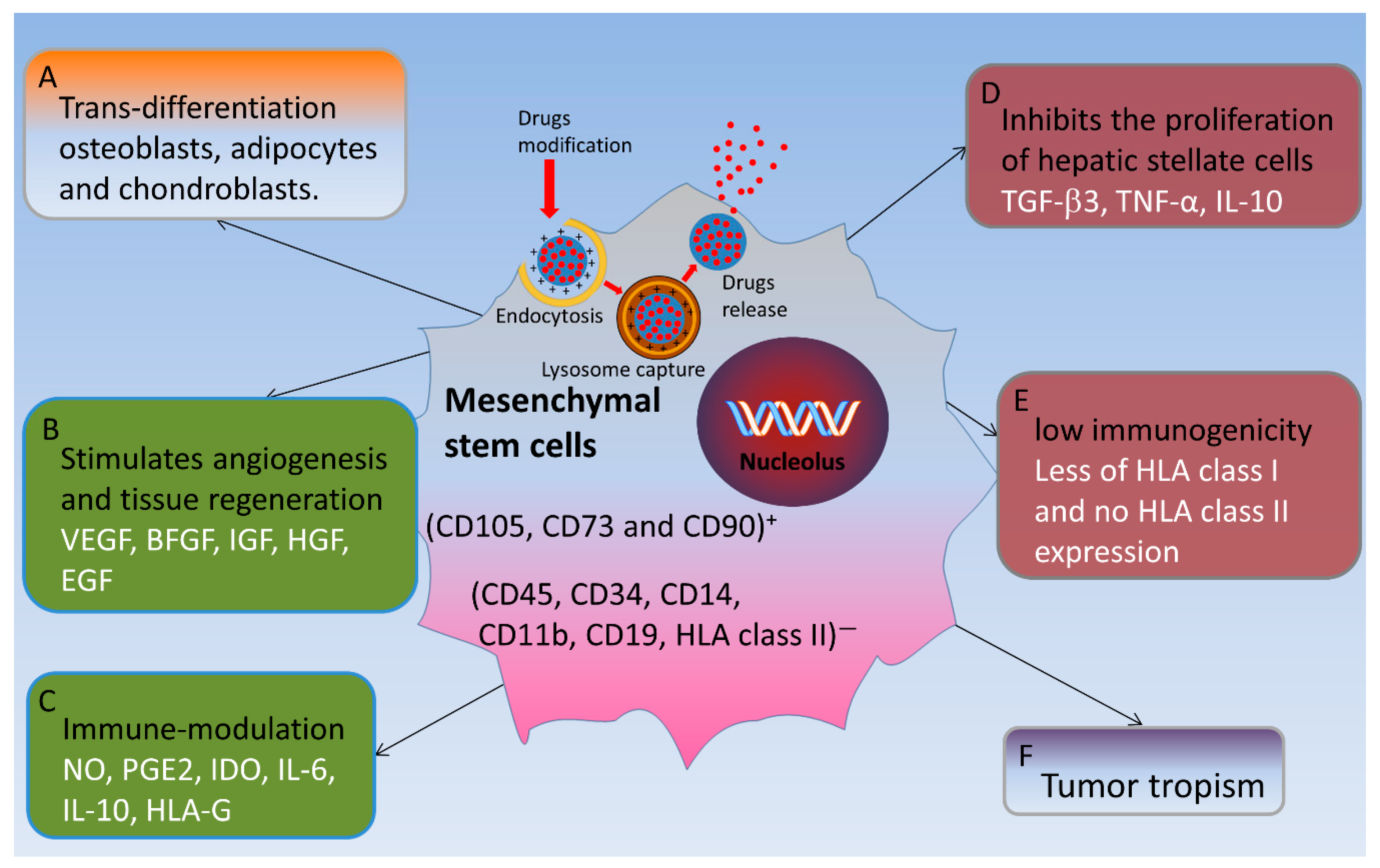
4.7. Pluripotent Mesenchymal Stem Cells for the Treatment of Liver Fibrosis
4.7.1. Functional with Low Immunogenicity and Inhibition of Inflammatory Responses
4.7.2. Tumor Tropism
4.7.3. Promotes the Proliferation and Regeneration of Hepatocytes
4.8. Research Studies on the Effects of Natural Plant Extractions for ALD
5. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACC | Acetyl-CoA Carboxylase |
| ADH | Alcohol Dehydrogenase |
| AFL | Alcoholic Fatty Liver |
| AH | Alcoholic Hepatitis |
| AKT | Serine/threonine-protein kinase B |
| ALD | Alcoholic Liver Disease |
| ALDH | Acetaldehyde Dehydrogenase |
| ALT | Alanine Transaminase |
| AMPK | AMP-activated protein kinase |
| ApoE | Apolipoprotein E |
| ASK | Apoptosis Signal-regulating Kinase 1 |
| AST | Aspartate Transaminase |
| ATP | Adenosine Triphosphate |
| BFGF | Basic Fibroblast Growth Factor |
| BMC | Bone Marrow Cells |
| Btg2 | B-cell translocation gene 2 |
| CAT | Catalase Endoplasmic Reticulum |
| CB | Cannabinoid Receptor |
| CCL20 | Chemokine (C-C motif) Ligand 20 |
| CDT | Carbohydrate-Deficient Transferrin |
| CXCL | Chemokine (C-X-C motif) Ligand 1 |
| CYP2E1 | Cytochrome P450 2E1 |
| CYP450 OR | Cytochrome P450 oxidoreductase |
| DAMPs | Damage-Associated Molecular Patterns |
| EGF | Epidermal Growth Factor |
| Egr-1 | Early growth response protein 1 |
| ER | Endoplasmic Reticulum |
| FABP4 | Fatty Acid Binding Protein |
| FOXO3A | Forkhead box O3 |
| GGT | Gamma-Glutamyltranspeptidase |
| GSK3β | Glycogen Synthase Kinase 3 beta |
| GSR | Glutathione reductase |
| HCV | Hepatitis C Virus |
| HGF | Hepatocyte Growth Factor |
| HIF1α | Hypoxia-Inducible Factor 1-alpha |
| HLA | Human Leukocyte Antigen |
| HMGCoA | 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase |
| HO-1 | Heme Oxygenase |
| HSC | Hepatic Stellate Cell |
| IDO | Indoleamine 2,3-dioxygenase |
| IGF | Insulin-like Growth Factor |
| Igf1R | Insulin-like growth factor 1 receptor |
| IL | Interleukin |
| JNK | C-Jun N-terminal kinases |
| iNOS | Inducible nitric oxide synthase |
| KLF6 | Kruppel-Like Factor 6 |
| LDLR | Low-Density Lipoprotein Receptor |
| LPS | Lipopolysaccharides |
| LXRα | Liver X Receptor alpha |
| MAPK | Mitogen-Activated Protein Kinase |
| MSC | Mesenchymal Stem Cell |
| MTTP | Microsomal Triglyceride Transfer Protein |
| NAD | Nicotinamide Adenine Dinucleotide |
| NF-κB | Nuclear Factor kappa-light-chain-enhancer of activated B |
| Nrf-2 | Nuclear factor erythroid 2-related factor 2 |
| PGC1a | Peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| PGE2 | Prostaglandin E2 |
| PNPLA3 | Patatin like phospholipase domain containing protein 3 |
| PPARα | Peroxisome Proliferator-Activated Receptor alpha |
| P4HA1 | Prolyl 4-hydroxylase subunit alpha 1 |
| ROS | Reactive Oxygen Species |
| RNS | Reactive Nitrogen Species |
| SAMe | S-adenosyl-L-methionine |
| SIRT1 | Sirtuin 1 |
| SNX10 | Sorting Nexin 10 |
| STAT3 | Signal Transducer and Activator of Transcription 3 |
| TGF-β3 | Transforming Growth Factor beta 3 |
| TfR1 | Transferrin Receptor 1 |
| TLR4 | Toll-like Receptor 4 |
| TNF-α | Tumor Necrosis Factor alpha |
| Treg | Regulatory T cells |
| Trx | Thioredoxin |
| TXNIP | Thioredoxin Interacting Protein |
| UCP | Uncoupling Protein |
| VEGF | Vascular Endothelial Growth Factor |
| Yy1 | Yin Yang 1 |
| ZER2 | Zinc finger E-box binding homeobox 2 |
References
- Poznyak, V.; Rekve, D. Management of Substance Abuse Team, Global Status Report on Alcohol and Health 2018; World Health Organization: Geneva, Switzerland, 2018; ISBN 978-92-4-156563-9. [Google Scholar]
- Dunn, W.; Shah, V.H. Pathogenesis of Alcoholic Liver Disease. Clin. Liver Dis. 2016, 20, 445–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galicia-Moreno, M.; Gutierrez-Reyes, G. The role of oxidative stress in the development of alcoholic liver disease. Rev. Gastroenterol. Mex. 2014, 79, 135–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altamirano, J.; Bataller, R. Alcoholic liver disease: Pathogenesis and new targets for therapy. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Bataller, R. Alcoholic liver disease: Pathogenesis and new therapeutic targets. Gastroenterology 2011, 141, 1572–1585. [Google Scholar] [CrossRef] [PubMed]
- Abenavoli, L.; Masarone, M.; Federico, A.; Rosato, V.; Dallio, M.; Loguercio, C.; Persico, M. Alcoholic Hepatitis: Pathogenesis, Diagnosis and Treatment. Rev. Recent Clin. Trials 2016, 11, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Orman, E.S.; Odena, G.; Bataller, R. Alcoholic liver disease: Pathogenesis, management, and novel targets for therapy. J. Gastroenterol. Hepatol. 2013, 28 (Suppl. S1), 77–84. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.S.; Ong, M.; Qu, X. Optimal management for alcoholic liver disease: Conventional medications, natural therapy or combination? World J. Gastroenterol. 2016, 22, 8–23. [Google Scholar] [CrossRef]
- Ezquer, F.; Bruna, F.; Calligaris, S.; Conget, P.; Ezquer, M. Multipotent mesenchymal stromal cells: A promising strategy to manage alcoholic liver disease. World J. Gastroenterol. 2016, 22, 24–36. [Google Scholar] [CrossRef]
- Suk, K.T.; Yoon, J.H.; Kim, M.Y.; Kim, C.W.; Kim, J.K.; Park, H.; Hwang, S.G.; Kim, D.J.; Lee, B.S.; Lee, S.H.; et al. Transplantation with autologous bone marrow-derived mesenchymal stem cells for alcoholic cirrhosis: Phase 2 trial. Hepatology 2016, 64, 2185–2197. [Google Scholar] [CrossRef]
- Marshall, W.J.; Bangert, S.K. Clinical biochemistry: Metabolic and clinical aspects; Churchill Livingstone: Edinburgh, NY, USA, 1995. [Google Scholar]
- Eriksson, C.J.; Fukunaga, T.; Sarkola, T.; Chen, W.J.; Chen, C.C.; Ju, J.M.; Cheng, A.T.; Yamamoto, H.; Kohlenberg-Muller, K.; Kimura, M.; et al. Functional relevance of human adh polymorphism. Alcohol. Clin. Exp. Res. 2001, 25 (Suppl. S5), 157S–163S. [Google Scholar] [CrossRef]
- Bosron, W.F.; Ehrig, T.; Li, T.K. Genetic factors in alcohol metabolism and alcoholism. Semin. Liver Dis. 1993, 13, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Cederbaum, A.I. Alcohol metabolism. Clin. Liver Dis. 2012, 16, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Ceni, E.; Mello, T.; Galli, A. Pathogenesis of alcoholic liver disease: Role of oxidative metabolism. World J. Gastroenterol. 2014, 20, 17756–17772. [Google Scholar] [CrossRef] [PubMed]
- Teschke, R. Alcoholic steatohepatitis (ASH) and alcoholic hepatitis (AH): Cascade of events, clinical aspects, and pharmacotherapy options. Expert Opin. Pharm. 2018, 19, 779–793. [Google Scholar] [CrossRef] [PubMed]
- Chacko, K.R.; Reinus, J. Spectrum of Alcoholic Liver Disease. Clin. Liver Dis. 2016, 20, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Torruellas, C.; French, S.W.; Medici, V. Diagnosis of alcoholic liver disease. World J. Gastroenterol. 2014, 20, 11684–11699. [Google Scholar] [CrossRef] [PubMed]
- Baraona, E.; Lieber, C.S. Effects of ethanol on lipid metabolism. J. Lipid Res. 1979, 20, 289–315. [Google Scholar]
- You, M.; Matsumoto, M.; Pacold, C.M.; Cho, W.K.; Crabb, D.W. The role of AMP-activated protein kinase in the action of ethanol in the liver. Gastroenterology 2004, 127, 1798–1808. [Google Scholar] [CrossRef]
- You, M.; Fischer, M.; Deeg, M.A.; Crabb, D.W. Ethanol induces fatty acid synthesis pathways by activation of sterol regulatory element-binding protein (SREBP). J. Biol. Chem. 2002, 277, 29342–29347. [Google Scholar] [CrossRef]
- Galli, A.; Pinaire, J.; Fischer, M.; Dorris, R.; Crabb, D.W. The transcriptional and DNA binding activity of peroxisome proliferator-activated receptor alpha is inhibited by ethanol metabolism. A novel mechanism for the development of ethanol-induced fatty liver. J. Biol. Chem. 2001, 276, 68–75. [Google Scholar] [CrossRef]
- Aday, A.W.; Mitchell, M.C.; Casey, L.C. Alcoholic hepatitis: Current trends in management. Curr. Opin. Gastroenterol. 2017, 33, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Odena, G.; Chen, J.; Lozano, J.J.; Altamirano, J.; Rodrigo-Torres, D.; Affo, S.; Morales-Ibanez, O.; Matsushita, H.; Zou, J.; Dumitru, R.; et al. LPS-TLR4 Pathway Mediates Ductular Cell Expansion in Alcoholic Hepatitis. Sci. Rep. 2016, 6, 35610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sancho-Bru, P.; Altamirano, J.; Rodrigo-Torres, D.; Coll, M.; Millan, C.; Jose Lozano, J.; Miquel, R.; Arroyo, V.; Caballeria, J.; Gines, P.; et al. Liver progenitor cell markers correlate with liver damage and predict short-term mortality in patients with alcoholic hepatitis. Hepatology 2012, 55, 1931–1941. [Google Scholar] [CrossRef] [PubMed]
- Sakhuja, P. Pathology of alcoholic liver disease, can it be differentiated from nonalcoholic steatohepatitis? World J. Gastroenterol. 2014, 20, 16474–16479. [Google Scholar] [CrossRef] [PubMed]
- Celli, R.; Zhang, X. Pathology of Alcoholic Liver Disease. J. Clin. Transl. Hepatol. 2014, 2, 103–109. [Google Scholar]
- Mandrekar, P.; Bataller, R.; Tsukamoto, H.; Gao, B. Alcoholic hepatitis: Translational approaches to develop targeted therapies. Hepatology 2016, 64, 1343–1355. [Google Scholar] [CrossRef]
- Mihm, S. Danger-Associated Molecular Patterns (DAMPs): Molecular Triggers for Sterile Inflammation in the Liver. Int. J. Mol. Sci. 2018, 19, 3104. [Google Scholar] [CrossRef]
- Dumortier, J.; Dharancy, S.; Cannesson, A.; Lassailly, G.; Rolland, B.; Pruvot, F.R.; Boillot, O.; Faure, S.; Guillaud, O.; Rigole-Donnadieu, H.; et al. Recurrent alcoholic cirrhosis in severe alcoholic relapse after liver transplantation: A frequent and serious complication. Am. J. Gastroenterol. 2015, 110, 1160–1166. [Google Scholar] [CrossRef]
- Gudowska, M.; Wojtowicz, E.; Cylwik, B.; Gruszewska, E.; Chrostek, L. The Distribution of Liver Steatosis, Fibrosis, Steatohepatitis and Inflammation Activity in Alcoholics According to FibroMax Test. Adv. Clin. Exp. Med. 2015, 24, 823–827. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.M.; Horiguchi, N.; Jeong, W.I.; Radaeva, S.; Gao, B. Molecular mechanisms of alcoholic liver disease: Innate immunity and cytokines. Alcohol. Clin. Exp. Res. 2011, 35, 787–793. [Google Scholar] [CrossRef]
- Breitkopf, K.; Nagy, L.E.; Beier, J.I.; Mueller, S.; Weng, H.; Dooley, S. Current experimental perspectives on the clinical progression of alcoholic liver disease. Alcohol. Clin Exp Res. 2009, 33, 1647–1655. [Google Scholar] [CrossRef] [PubMed]
- Rao, R. Endotoxemia and gut barrier dysfunction in alcoholic liver disease. Hepatology 2009, 50, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Seth, D.; Haber, P.S.; Syn, W.K.; Diehl, A.M.; Day, C.P. Pathogenesis of alcohol-induced liver disease: Classical concepts and recent advances. J. Gastroenterol. Hepatol. 2011, 26, 1089–1105. [Google Scholar] [CrossRef] [PubMed]
- Parola, M.; Robino, G. Oxidative stress-related molecules and liver fibrosis. J. Hepatol 2001, 35, 297–306. [Google Scholar] [CrossRef]
- Seitz, H.K.; Bataller, R.; Cortez-Pinto, H.; Gao, B.; Gual, A.; Lackner, C.; Mathurin, P.; Mueller, S.; Szabo, G.; Tsukamoto, H. Alcoholic liver disease. Nat. Rev. Dis. Primers 2018, 4, 16. [Google Scholar] [CrossRef]
- Zhao, N.; Guo, F.F.; Xie, K.Q.; Zeng, T. Targeting Nrf-2 is a promising intervention approach for the prevention of ethanol-induced liver disease. Cell. Mol. Life Sci. 2018, 75, 3143–3157. [Google Scholar] [CrossRef]
- McClain, C.; Vatsalya, V.; Cave, M. Role of Zinc in the Development/Progression of Alcoholic Liver Disease. Curr. Treat. Options. Gastroenterol. 2017, 15, 285–295. [Google Scholar] [CrossRef]
- Han, K.H.; Hashimoto, N.; Fukushima, M. Relationships among alcoholic liver disease, antioxidants, and antioxidant enzymes. World J. Gastroenterol. 2016, 22, 37–49. [Google Scholar] [CrossRef]
- Pang, C.; Sheng, Y.C.; Jiang, P.; Wei, H.; Ji, L.L. Chlorogenic acid prevents acetaminophen-induced liver injury: The involvement of CYP450 metabolic enzymes and some antioxidant signals. J. Zhejiang Univ. Sci. B 2015, 16, 602–610. [Google Scholar] [CrossRef] [Green Version]
- Purohit, V.; Bode, J.C.; Bode, C.; Brenner, D.A.; Choudhry, M.A.; Hamilton, F.; Kang, Y.J.; Keshavarzian, A.; Rao, R.; Sartor, R.B.; et al. Alcohol, intestinal bacterial growth, intestinal permeability to endotoxin, and medical consequences: Summary of a symposium. Alcohol 2008, 42, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Groebner, J.L.; Tuma, P.L. The Altered Hepatic Tubulin Code in Alcoholic Liver Disease. Biomolecules 2015, 5, 2140–2159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, G. Gut-liver axis in alcoholic liver disease. Gastroenterology 2015, 148, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.Y.; Xu, J.; Liu, X.; Zhu, Y.; Gao, B.; Karin, M.; Tsukamoto, H.; Jeste, D.V.; Grant, I.; Roberts, A.J.; et al. The role of IL-17 signaling in regulation of the liver-brain axis and intestinal permeability in Alcoholic Liver Disease. Curr. Pathobiol. Rep. 2016, 4, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Harrison-Findik, D.D. Role of alcohol in the regulation of iron metabolism. World J. Gastroenterol. 2007, 13, 4925–4930. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, G.N.; Dominitz, J.A.; Weiss, N.S.; Heagerty, P.J.; Kowdley, K.V. The effect of alcohol consumption on the prevalence of iron overload, iron deficiency, and iron deficiency anemia. Gastroenterology 2004, 126, 1293–1301. [Google Scholar] [CrossRef] [PubMed]
- Detivaud, L.; Nemeth, E.; Boudjema, K.; Turlin, B.; Troadec, M.B.; Leroyer, P.; Ropert, M.; Jacquelinet, S.; Courselaud, B.; Ganz, T.; et al. Hepcidin levels in humans are correlated with hepatic iron stores, hemoglobin levels, and hepatic function. Blood 2005, 106, 746–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Hepcidin, a key regulator of iron metabolism and mediator of anemia of inflammation. Blood 2003, 102, 783–788. [Google Scholar] [CrossRef] [Green Version]
- Harrison-Findik, D.D.; Schafer, D.; Klein, E.; Timchenko, N.A.; Kulaksiz, H.; Clemens, D.; Fein, E.; Andriopoulos, B.; Pantopoulos, K.; Gollan, J. Alcohol metabolism-mediated oxidative stress down-regulates hepcidin transcription and leads to increased duodenal iron transporter expression. J. Biol. Chem. 2006, 281, 22974–22982. [Google Scholar] [CrossRef]
- Harrison-Findik, D.D.; Klein, E.; Crist, C.; Evans, J.; Timchenko, N.; Gollan, J. Iron-mediated regulation of liver hepcidin expression in rats and mice is abolished by alcohol. Hepatology 2007, 46, 1979–1985. [Google Scholar] [CrossRef]
- Pietrangelo, A. Iron, oxidative stress and liver fibrogenesis. J. Hepatol. 1998, 28 (Suppl. S1), 8–13. [Google Scholar] [CrossRef]
- Valerio, L.G.; Parks, T., Jr.; Petersen, D.R. Alcohol mediates increases in hepatic and serum nonheme iron stores in a rat model for alcohol-induced liver injury. Alcohol. Clin. Exp. Res. 1996, 20, 1352–1361. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, H.; Horne, W.; Kamimura, S.; Niemela, O.; Parkkila, S.; Yla-Herttuala, S.; Brittenham, G.M. Experimental liver cirrhosis induced by alcohol and iron. J. Clin. Invest. 1995, 96, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Silva, I.; Rausch, V.; Seitz, H.K.; Mueller, S. Does Hypoxia Cause Carcinogenic Iron Accumulation in Alcoholic Liver Disease (ALD)? Cancers (Basel) 2017, 9, 145. [Google Scholar] [CrossRef] [PubMed]
- Abdelmegeed, M.A.; Banerjee, A.; Jang, S.; Yoo, S.H.; Yun, J.W.; Gonzalez, F.J.; Keshavarzian, A.; Song, B.J. CYP2E1 potentiates binge alcohol-induced gut leakiness, steatohepatitis, and apoptosis. Free Radic. Biol. Med. 2013, 65, 1238–1245. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.K.; Yu, T.; de la Monte, S.M.; Wands, J.R.; Derdak, Z.; Kim, M. Restoration of Wnt/beta-catenin signaling attenuates alcoholic liver disease progression in a rat model. J. Hepatol. 2015, 63, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Ibusuki, R.; Uto, H.; Oda, K.; Ohshige, A.; Tabu, K.; Mawatari, S.; Kumagai, K.; Kanmura, S.; Tamai, T.; Moriuchi, A.; et al. Human neutrophil peptide-1 promotes alcohol-induced hepatic fibrosis and hepatocyte apoptosis. PLoS ONE 2017, 12, e0174913. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Shukla, S.D. Histone H3 phosphorylation at serine 10 and serine 28 is mediated by p38 MAPK in rat hepatocytes exposed to ethanol and acetaldehyde. Eur. J. Pharm. 2007, 573, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Yang, Y.; Ren, L.; Shao, T.; Li, F.; Zhao, C.; Liu, L.; Zhang, H.; McClain, C.J.; Feng, W. Activation of autophagy attenuates EtOH-LPS-induced hepatic steatosis and injury through MD2 associated TLR4 signaling. Sci. Rep. 2017, 7, 9292. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, J.; Wu, Z.; Wang, X.; Liu, L.; Yao, C. Cyanidin 3-O-beta-Glucoside Ameliorates Ethanol-Induced Acute Liver Injury by Attenuating Oxidative Stress and Apoptosis: The Role of SIRT1/FOXO1 Signaling. Alcohol. Clin. Exp. Res. 2016, 40, 457–466. [Google Scholar] [CrossRef]
- Pani, G.; Fusco, S.; Colavitti, R.; Borrello, S.; Maggiano, N.; Cravero, A.A.; Farre, S.M.; Galeotti, T.; Koch, O.R. Abrogation of hepatocyte apoptosis and early appearance of liver dysplasia in ethanol-fed p53-deficient mice. Biochem. Biophys. Res. Commun. 2004, 325, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Ke, Z.; Xu, M.; Liao, M.; Wang, X.; Qi, Y.; Zhang, T.; Frank, J.A.; Bower, K.A.; Shi, X.; et al. Autophagy is a protective response to ethanol neurotoxicity. Autophagy 2012, 8, 1577–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Cui, R.; Zhao, J.; Mo, R.; Peng, L.; Yan, M. Corosolic acid protects hepatocytes against ethanol-induced damage by modulating mitogen-activated protein kinases and activating autophagy. Eur. J. Pharm. 2016, 791, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Cederbaum, A.I. Autophagy Protects against CYP2E1/Chronic Ethanol-Induced Hepatotoxicity. Biomolecules 2015, 5, 2659–2674. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.M.; Bhakta, A.; Wang, S.; Li, Z.; Manley, S.; Huang, H.; Copple, B.; Ding, W.X. Role of hypoxia inducing factor-1beta in alcohol-induced autophagy, steatosis and liver injury in mice. PLoS ONE 2014, 9, e115849. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Yin, S.; Huo, Y.; Liang, M.; Fan, L.; Ye, M.; Hu, H. Glycycoumarin ameliorates alcohol-induced hepatotoxicity via activation of Nrf2 and autophagy. Free Radic. Biol. Med. 2015, 89, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Denaes, T.; Lodder, J.; Chobert, M.N.; Ruiz, I.; Pawlotsky, J.M.; Lotersztajn, S.; Teixeira-Clerc, F. The Cannabinoid Receptor 2 Protects Against Alcoholic Liver Disease Via a Macrophage Autophagy-Dependent Pathway. Sci. Rep. 2016, 6, 28806. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Xu, X.; Babcock, S.A.; Zhang, Y.; Ren, J. Aldehyde dedydrogenase-2 plays a beneficial role in ameliorating chronic alcohol-induced hepatic steatosis and inflammation through regulation of autophagy. J. Hepatol. 2015, 62, 647–656. [Google Scholar] [CrossRef]
- Nepal, S.; Park, P.H. Activation of autophagy by globular adiponectin attenuates ethanol-induced apoptosis in HepG2 cells: Involvement of AMPK/FoxO3A axis. Biochim. Biophys. Acta. 2013, 1833, 2111–2125. [Google Scholar] [CrossRef] [Green Version]
- You, Y.; Li, W.Z.; Zhang, S.; Hu, B.; Li, Y.X.; Li, H.D.; Tang, H.H.; Li, Q.W.; Guan, Y.Y.; Liu, L.X.; et al. SNX10 mediates alcohol-induced liver injury and steatosis by regulating the activation of chaperone-mediated autophagy. J. Hepatol. 2018, 69, 129–141. [Google Scholar] [CrossRef]
- Stickel, F.; Datz, C.; Hampe, J.; Bataller, R. Pathophysiology and Management of Alcoholic Liver Disease: Update 2016. Gut. Liver 2017, 11, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Kolla, B.P.; Schneekloth, T.D.; Biernacka, J.; Shah, V.; Lazaridis, K.N.; Geske, J.; Karpyak, V. PNPLA3 Association with Alcoholic Liver Disease in a Cohort of Heavy Drinkers. Alcohol Alcohol. 2018, 53, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, T.; Yang, F.; Mao, Y.; Li, L.; Liu, C.; Sun, Q.; Li, Y.; Huang, J. Single-nucleotide rs738409 polymorphisms in the PNPLA3 gene are strongly associated with alcoholic liver disease in Han Chinese males. Hepatol. Int. 2018, 12, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, K.; Wannhoff, A.; Kattner, S.; Brune, M.; Hov, J.R.; Weiss, K.H.; Antoni, C.; Dollinger, M.; Neumann-Haefelin, C.; Seufferlein, T.; et al. PNPLA3 in end-stage liver disease: Alcohol consumption, hepatocellular carcinoma development; transplantation-free survival. J. Gastroenterol. Hepatol. 2014, 29, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.M.; Fan, J.G.; Wang, B.Y.; Lu, L.G.; Shi, J.P.; Niu, J.Q.; Shen, W. Chinese Association for the Study of Liver, D. Guidelines for the diagnosis and management of alcoholic liver disease: Update 2010: (published in Chin. Chin. J. Hepatol. 2010, 18, 167–170). J. Dig. Dis. 2011, 12, 45–50. [Google Scholar]
- Singal, A.K.; Bataller, R.; Ahn, J.; Kamath, P.S.; Shah, V.H. ACG Clinical Guideline: Alcoholic Liver Disease. Am. J. Gastroenterol. 2018, 113, 175–194. [Google Scholar] [CrossRef] [PubMed]
- Sueyoshi, S.; Sawai, S.; Satoh, M.; Seimiya, M.; Sogawa, K.; Fukumura, A.; Tsutsumi, M.; Nomura, F. Fractionation of gamma-glutamyltransferase in patients with nonalcoholic fatty liver disease and alcoholic liver disease. World J. Hepatol. 2016, 8, 1610–1616. [Google Scholar] [CrossRef] [PubMed]
- Palmentieri, B.; de Sio, I.; La Mura, V.; Masarone, M.; Vecchione, R.; Bruno, S.; Torella, R.; Persico, M. The role of bright liver echo pattern on ultrasound B-mode examination in the diagnosis of liver steatosis. Dig. Liver Dis. 2006, 38, 485–489. [Google Scholar] [CrossRef]
- Grandison, G.A.; Angulo, P. Can NASH be diagnosed, graded, and staged noninvasively? Clin. Liver Dis. 2012, 16, 567–585. [Google Scholar] [CrossRef]
- Kollmann, D.; Rasoul-Rockenschaub, S.; Steiner, I.; Freundorfer, E.; Gyori, G.P.; Silberhumer, G.; Soliman, T.; Berlakovich, G.A. Good outcome after liver transplantation for ALD without a 6 months abstinence rule prior to transplantation including post-transplant CDT monitoring for alcohol relapse assessment-a retrospective study. Transpl. Int. 2016, 29, 559–567. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver. EASL clinical practical guidelines: Management of alcoholic liver disease. J. Hepatol. 2012, 57, 399–420. [Google Scholar] [CrossRef]
- Vasiliou, V. Biological basis of alcohol-induced cancer; Springer Science+Business Media: New York, NY, USA, 2014. [Google Scholar]
- Suk, K.T.; Kim, M.Y.; Baik, S.K. Alcoholic liver disease: Treatment. World J. Gastroenterol. 2014, 20, 12934–12944. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, S.S.; Goyal, O.; Kishore, H.; Sidhu, S. New paradigms in management of alcoholic hepatitis: A review. Hepatol. Int. 2017, 11, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Morley, K.C.; Leung, S.; Baillie, A.; Haber, P.S. The efficacy and biobehavioural basis of baclofen in the treatment of alcoholic liver disease (BacALD): Study protocol for a randomised controlled trial. Contemp. Clin. Trials 2013, 36, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Salama, Z.A.; Sadek, A.; Abdelhady, A.M.; Darweesh, S.K.; Morsy, S.A.; Esmat, G. Losartan may inhibit the progression of liver fibrosis in chronic HCV patients. Hepatobiliary Surg. Nutr. 2016, 5, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sookoian, S.; Fernandez, M.A.; Castano, G. Effects of six months losartan administration on liver fibrosis in chronic hepatitis C patients: A pilot study. World J. Gastroenterol. 2005, 11, 7560–7563. [Google Scholar] [CrossRef] [PubMed]
- Rambaldi, A.; Iaquinto, G.; Gluud, C. Anabolic-androgenic steroids for alcoholic liver disease: A Cochrane review. Am. J. Gastroenterol. 2002, 97, 1674–1681. [Google Scholar] [CrossRef]
- Marroni, C.A.; Fleck, A.M.; Fernandes, S.A., Jr.; Galant, L.H.; Mucenic, M.; de Mattos Meine, M.H.; Mariante-Neto, G.; Brandao, A.B.M. Liver transplantation and alcoholic liver disease: History, controversies, and considerations. World J. Gastroenterol. 2018, 24, 2785–2805. [Google Scholar] [CrossRef]
- Iruzubieta, P.; Crespo, J.; Fabrega, E. Long-term survival after liver transplantation for alcoholic liver disease. World J. Gastroenterol. 2013, 19, 9198–9208. [Google Scholar] [CrossRef]
- Rice, J.P.; Eickhoff, J.; Agni, R.; Ghufran, A.; Brahmbhatt, R.; Lucey, M.R. Abusive drinking after liver transplantation is associated with allograft loss and advanced allograft fibrosis. Liver Transpl. 2013, 19, 1377–1386. [Google Scholar] [CrossRef]
- Testino, G.; Leone, S.; Ansaldi, F.; Borro, P. Silymarin and S-adenosyl-L-methionine (SAMe): Two promising pharmacological agents in case of chronic alcoholic hepathopathy. A review and a point of view. Minerva. Gastroenterol. Dietol. 2013, 59, 341–356. [Google Scholar]
- Kessova, I.G.; Cederbaum, A.I. Mitochondrial alterations in livers of Sod1-/- mice fed alcohol. Free Radic. Biol. Med. 2007, 42, 1470–1480. [Google Scholar] [CrossRef]
- Assiri, M.A.; Roy, S.R.; Harris, P.S.; Ali, H.; Liang, Y.; Shearn, C.T.; Orlicky, D.J.; Roede, J.R.; Hirschey, M.D.; Backos, D.S.; et al. Chronic Ethanol Metabolism Inhibits Hepatic Mitochondrial Superoxide Dismutase via Lysine Acetylation. Alcohol. Clin. Exp. Res. 2017, 41, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, A.; Pinkaew, D.; Doan, H.Q.; Jacob, R.B.; Verma, S.K.; Friedman, H.; Peterson, A.C.; Kuyumcu-Martinez, M.N.; McDougal, O.M.; Fujise, K. Fortilin potentiates the peroxidase activity of Peroxiredoxin-1 and protects against alcohol-induced liver damage in mice. Sci. Rep. 2016, 6, 18701. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.H.; Sung, S.H.; Cho, E.J.; Lee, S.K.; Lee, H.E.; Woo, H.A.; Yu, D.Y.; Kil, I.S.; Rhee, S.G. Concerted action of sulfiredoxin and peroxiredoxin I protects against alcohol-induced oxidative injury in mouse liver. Hepatology 2011, 53, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Singh, S.; Matsumoto, A.; Manna, S.K.; Abdelmegeed, M.A.; Golla, S.; Murphy, R.C.; Dong, H.; Song, B.J.; Gonzalez, F.J.; et al. Chronic Glutathione Depletion Confers Protection against Alcohol-induced Steatosis: Implication for Redox Activation of AMP-activated Protein Kinase Pathway. Sci. Rep. 2016, 6, 29743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakhautdin, B.; Das, D.; Mandal, P.; Roychowdhury, S.; Danner, J.; Bush, K.; Pollard, K.; Kaspar, J.W.; Li, W.; Salomon, R.G.; et al. Protective role of HO-1 and carbon monoxide in ethanol-induced hepatocyte cell death and liver injury in mice. J. Hepatol. 2014, 61, 1029–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Cederbaum, A.I. Acute ethanol pretreatment increases FAS-mediated liver injury in mice: Role of oxidative stress and CYP2E1-dependent and -independent pathways. Free Radic. Biol. Med. 2007, 42, 971–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koteish, A.; Yang, S.; Lin, H.; Huang, X.; Diehl, A.M. Chronic ethanol exposure potentiates lipopolysaccharide liver injury despite inhibiting Jun N-terminal kinase and caspase 3 activation. J. Biol. Chem. 2002, 277, 13037–13044. [Google Scholar] [CrossRef]
- Miller, A.M.; Wang, H.; Park, O.; Horiguchi, N.; Lafdil, F.; Mukhopadhyay, P.; Moh, A.; Fu, X.Y.; Kunos, G.; Pacher, P.; et al. Anti-inflammatory and anti-apoptotic roles of endothelial cell STAT3 in alcoholic liver injury. Alcohol. Clin. Exp. Res. 2010, 34, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Hoek, J.B.; Pastorino, J.G. Cellular signaling mechanisms in alcohol-induced liver damage. Semin. Liver Dis. 2004, 24, 257–272. [Google Scholar] [CrossRef]
- Lee, Y.J.; Shukla, S.D. Pro- and anti-apoptotic roles of c-Jun N-terminal kinase (JNK) in ethanol and acetaldehyde exposed rat hepatocytes. Eur. J. Pharm. 2005, 508, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Yin, S.; Jiang, X.; Huang, D.; Shen, G. Melatonin protects against alcoholic liver injury by attenuating oxidative stress, inflammatory response, and apoptosis. Eur. J. Pharm. 2009, 616, 287–292. [Google Scholar] [CrossRef]
- Zhao, H.; Zhao, C.; Dong, Y.; Zhang, M.; Wang, Y.; Li, F.; Li, X.; McClain, C.; Yang, S.; Feng, W. Inhibition of miR122a by Lactobacillus rhamnosus GG culture supernatant increases intestinal occludin expression and protects mice from alcoholic liver disease. Toxicol. Lett. 2015, 234, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.Y.; Liu, J.D.; Lu, X.Y.; Yan, X.; Huang, C.; Meng, X.M.; Li, J. miR-203 Inhibits Alcohol-Induced Hepatic Steatosis by Targeting Lipin1. Front. Pharm. 2018, 9, 275. [Google Scholar] [CrossRef] [PubMed]
- Bala, S.; Petrasek, J.; Mundkur, S.; Catalano, D.; Levin, I.; Ward, J.; Alao, H.; Kodys, K.; Szabo, G. Circulating microRNAs in exosomes indicate hepatocyte injury and inflammation in alcoholic, drug-induced, and inflammatory liver diseases. Hepatology 2012, 56, 1946–1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satishchandran, A.; Ambade, A.; Rao, S.; Hsueh, Y.C.; Iracheta-Vellve, A.; Tornai, D.; Lowe, P.; Gyongyosi, B.; Li, J.; Catalano, D.; et al. MicroRNA 122, Regulated by GRLH2, Protects Livers of Mice and Patients From Ethanol-Induced Liver Disease. Gastroenterology 2018, 154, 238–252.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, G.; Satishchandran, A. MicroRNAs in alcoholic liver disease. Semin. Liver Dis. 2015, 35, 36–42. [Google Scholar]
- Jin, X.; Yu, M.S.; Huang, Y.; Xiang, Z.; Chen, Y.P. MiR-30e-UCP2 pathway regulates alcoholic hepatitis progress by influencing ATP and hydrogen peroxide expression. Oncotarget 2017, 8, 64294–64302. [Google Scholar] [CrossRef] [Green Version]
- Brandon-Warner, E.; Feilen, N.A.; Culberson, C.R.; Field, C.O.; deLemos, A.S.; Russo, M.W.; Schrum, L.W. Processing of miR17-92 Cluster in Hepatic Stellate Cells Promotes Hepatic Fibrogenesis During Alcohol-Induced Injury. Alcohol Clin. Exp. Res. 2016, 40, 1430–1442. [Google Scholar] [CrossRef]
- Kim, Y.D.; Hwang, S.L.; Lee, E.J.; Kim, H.M.; Chung, M.J.; Elfadl, A.K.; Lee, S.E.; Nedumaran, B.; Harris, R.A.; Jeong, K.S. Melatonin ameliorates alcohol-induced bile acid synthesis by enhancing miR-497 expression. J. Pineal. Res. 2017, 62, e12386. [Google Scholar] [CrossRef]
- Heo, M.J.; Kim, T.H.; You, J.S.; Blaya, D.; Sancho-Bru, P.; Kim, S.G. Alcohol dysregulates miR-148a in hepatocytes through FoxO1, facilitating pyroptosis via TXNIP overexpression. Gut 2018, 68, 708–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.; Liang, X.; Jogasuria, A.; Davidson, N.O.; You, M. miR-217 regulates ethanol-induced hepatic inflammation by disrupting sirtuin 1-lipin-1 signaling. Am. J. Pathol. 2015, 185, 1286–1296. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Liu, H.; Chen, F.; Li, D.; Zhao, Y. MiR-214 promotes the alcohol-induced oxidative stress via down-regulation of glutathione reductase and cytochrome P450 oxidoreductase in liver cells. Alcohol Clin. Exp. Res. 2014, 38, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Bala, S.; Szabo, G. MicroRNA Signature in Alcoholic Liver Disease. Int. J. Hepatol. 2012, 2012, 498232. [Google Scholar] [CrossRef] [PubMed]
- Bala, S.; Csak, T.; Saha, B.; Zatsiorsky, J.; Kodys, K.; Catalano, D.; Satishchandran, A.; Szabo, G. The pro-inflammatory effects of miR-155 promote liver fibrosis and alcohol-induced steatohepatitis. J. Hepatol. 2016, 64, 1378–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eguchi, A.; Lazaro, R.G.; Wang, J.; Kim, J.; Povero, D.; Willliams, B.; Ho, S.B.; Starkel, P.; Schnabl, B.; Ohno-Machado, L.; et al. Extracellular vesicles released by hepatocytes from gastric infusion model of alcoholic liver disease contain a MicroRNA barcode that can be detected in blood. Hepatology 2017, 65, 475–490. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.X.; Sun, Y.Y.; Huang, A.L.; Li, X.F.; Huang, C.; Ma, T.T.; Li, J. MicroRNA-200a induces apoptosis by targeting ZEB2 in alcoholic liver disease. Cell Cycle 2018, 17, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Iwagami, Y.; Zou, J.; Zhang, H.; Cao, K.; Ji, C.; Kim, M.; Huang, C.K. Alcohol-mediated miR-34a modulates hepatocyte growth and apoptosis. J. Cell. Mol. Med. 2018, 22, 3897–3995. [Google Scholar] [CrossRef]
- El-Assal, O.; Hong, F.; Kim, W.H.; Radaeva, S.; Gao, B. IL-6-deficient mice are susceptible to ethanol-induced hepatic steatosis: IL-6 protects against ethanol-induced oxidative stress and mitochondrial permeability transition in the liver. Cell. Mol. Immunol. 2004, 1, 205–211. [Google Scholar]
- Tilg, H.; Moschen, A.R.; Szabo, G. Interleukin-1 and inflammasomes in alcoholic liver disease/acute alcoholic hepatitis and nonalcoholic fatty liver disease/nonalcoholic steatohepatitis. Hepatology 2016, 64, 955–965. [Google Scholar] [CrossRef]
- Bertola, A.; Park, O.; Gao, B. Chronic plus binge ethanol feeding synergistically induces neutrophil infiltration and liver injury in mice: A critical role for E-selectin. Hepatology 2013, 58, 1814–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ki, S.H.; Park, O.; Zheng, M.; Morales-Ibanez, O.; Kolls, J.K.; Bataller, R.; Gao, B. Interleukin-22 treatment ameliorates alcoholic liver injury in a murine model of chronic-binge ethanol feeding: Role of signal transducer and activator of transcription 3. Hepatology 2010, 52, 1291–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago-Lomeli, M.; Gomez-Quiroz, L.E.; Ortiz-Ortega, V.M.; Kershenobich, D.; Gutierrez-Ruiz, M.C. Differential effect of interleukin-10 on hepatocyte apoptosis. Life Sci. 2005, 76, 2569–2579. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.H.; Huo, L.J.; Li, T.T. Antioxidant axis Nrf2-keap1-ARE in inhibition of alcoholic liver fibrosis by IL-22. World J. Gastroenterol. 2017, 23, 2002–2011. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, M.; Miquel, R.; Colmenero, J.; Moreno, M.; Garcia-Pagan, J.C.; Bosch, J.; Arroyo, V.; Gines, P.; Caballeria, J.; Bataller, R. Hepatic expression of CXC chemokines predicts portal hypertension and survival in patients with alcoholic hepatitis. Gastroenterology 2009, 136, 1639–1650. [Google Scholar] [CrossRef] [PubMed]
- Affo, S.; Morales-Ibanez, O.; Rodrigo-Torres, D.; Altamirano, J.; Blaya, D.; Dapito, D.H.; Millan, C.; Coll, M.; Caviglia, J.M.; Arroyo, V.; et al. CCL20 mediates lipopolysaccharide induced liver injury and is a potential driver of inflammation and fibrosis in alcoholic hepatitis. Gut 2014, 63, 1782–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degre, D.; Lemmers, A.; Gustot, T.; Ouziel, R.; Trepo, E.; Demetter, P.; Verset, L.; Quertinmont, E.; Vercruysse, V.; Le Moine, O.; et al. Hepatic expression of CCL2 in alcoholic liver disease is associated with disease severity and neutrophil infiltrates. Clin. Exp. Immunol. 2012, 169, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Roh, Y.S.; Zhang, B.; Loomba, R.; Seki, E. TLR2 and TLR9 contribute to alcohol-mediated liver injury through induction of CXCL1 and neutrophil infiltration. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G30–G41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambade, A.; Lowe, P.; Kodys, K.; Catalano, D.; Gyongyosi, B.; Cho, Y.; Iracheta Vellve, A.; Adejumo, A.; Saha, B.; Calenda, C.; et al. Pharmacological inhibition of CCR2/5 signaling prevents and reverses alcohol-induced liver damage, steatosis and inflammation in mice. Hepatology 2018. [Google Scholar] [CrossRef] [PubMed]
- Karim, S.; Liaskou, E.; Hadley, S.; Youster, J.; Faint, J.; Adams, D.H.; Lalor, P.F. An in vitro model of human acute ethanol exposure that incorporates CXCR3- and CXCR4-dependent recruitment of immune cells. Toxicol. Sci. 2013, 132, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Cassard, A.M.; Ciocan, D. Microbiota, a key player in alcoholic liver disease. Clin. Mol. Hepatol. 2018, 24, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, P.; Chen, W.C.; Schnabl, B. The intestinal microbiome and the leaky gut as therapeutic targets in alcoholic liver disease. Front. Physiol. 2012, 3, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, M.; Han, D.H.; Hong, J.; Kim, D.J.; Suk, K.T. Are Probiotics Effective in Targeting Alcoholic Liver Diseases? Probiotics Antimicrob. Proteins 2018. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.C.; Xu, L.M.; Du, S.J.; Huang, S.S.; Wu, H.; Dong, J.J.; Huang, J.R.; Wang, X.D.; Feng, W.K.; Chen, Y.P. Lactobacillus rhamnosus GG supernatant promotes intestinal barrier function, balances Treg and TH17 cells and ameliorates hepatic injury in a mouse model of chronic-binge alcohol feeding. Toxicol. Lett. 2016, 241, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Kim, S.W.; Han, S.H.; Kim, D.J.; Suk, K.T.; Kim, Y.S.; Kim, M.J.; Kim, M.Y.; Baik, S.K.; Ham, Y.L. Probiotics (Lactobacillus rhamnosus R0011 and acidophilus R0052) reduce the expression of toll-like receptor 4 in mice with alcoholic liver disease. PLoS ONE 2015, 10, e0117451. [Google Scholar] [CrossRef] [PubMed]
- Chiu, W.C.; Huang, Y.L.; Chen, Y.L.; Peng, H.C.; Liao, W.H.; Chuang, H.L.; Chen, J.R.; Yang, S.C. Synbiotics reduce ethanol-induced hepatic steatosis and inflammation by improving intestinal permeability and microbiota in rats. Food Funct. 2015, 6, 1692–1700. [Google Scholar] [CrossRef] [PubMed]
- Grander, C.; Adolph, T.E.; Wieser, V.; Lowe, P.; Wrzosek, L.; Gyongyosi, B.; Ward, D.V.; Grabherr, F.; Gerner, R.R.; Pfister, A.; et al. Recovery of ethanol-induced Akkermansia muciniphila depletion ameliorates alcoholic liver disease. Gut 2018, 67, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Barone, R.; Rappa, F.; Macaluso, F.; Caruso Bavisotto, C.; Sangiorgi, C.; Di Paola, G.; Tomasello, G.; Di Felice, V.; Marciano, V.; Farina, F.; et al. Alcoholic Liver Disease: A Mouse Model Reveals Protection by Lactobacillus fermentum. Clin. Transl. Gastroenterol. 2016, 7, e138. [Google Scholar] [CrossRef]
- Apte, U.M.; Banerjee, A.; McRee, R.; Wellberg, E.; Ramaiah, S.K. Role of osteopontin in hepatic neutrophil infiltration during alcoholic steatohepatitis. Toxicol. Appl. Pharm. 2005, 207, 25–38. [Google Scholar] [CrossRef]
- Seth, D.; Duly, A.; Kuo, P.C.; McCaughan, G.W.; Haber, P.S. Osteopontin is an important mediator of alcoholic liver disease via hepatic stellate cell activation. World J. Gastroenterol. 2014, 20, 13088–13104. [Google Scholar] [CrossRef]
- Ge, X.; Leung, T.M.; Arriazu, E.; Lu, Y.; Urtasun, R.; Christensen, B.; Fiel, M.I.; Mochida, S.; Sorensen, E.S.; Nieto, N. Osteopontin binding to lipopolysaccharide lowers tumor necrosis factor-alpha and prevents early alcohol-induced liver injury in mice. Hepatology 2014, 59, 1600–1616. [Google Scholar] [CrossRef] [PubMed]
- Lazaro, R.; Wu, R.; Lee, S.; Zhu, N.L.; Chen, C.L.; French, S.W.; Xu, J.; Machida, K.; Tsukamoto, H. Osteopontin deficiency does not prevent but promotes alcoholic neutrophilic hepatitis in mice. Hepatology 2015, 61, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Magdaleno, F.; Ge, X.; Fey, H.; Lu, Y.; Gaskell, H.; Blajszczak, C.C.; Aloman, C.; Fiel, M.I.; Nieto, N. Osteopontin deletion drives hematopoietic stem cell mobilization to the liver and increases hepatic iron contributing to alcoholic liver disease. Hepatol. Commun. 2018, 2, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Basu, P.P.; Aloysius, M.M.; Shah, N.J.; Brown, R.S., Jr. Review article: The endocannabinoid system in liver disease, a potential therapeutic target. Aliment. Pharm. 2014, 39, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Trebicka, J.; Racz, I.; Siegmund, S.V.; Cara, E.; Granzow, M.; Schierwagen, R.; Klein, S.; Wojtalla, A.; Hennenberg, M.; Huss, S.; et al. Role of cannabinoid receptors in alcoholic hepatic injury: Steatosis and fibrogenesis are increased in CB2 receptor-deficient mice and decreased in CB1 receptor knockouts. Liver Int. 2011, 31, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Y.; Lin, H.C. Cross-regulation of cannabinoid CB(1) and CB(2) receptors governs hepatic steatosis. Liver Int. 2010, 30, 1397–1399. [Google Scholar] [CrossRef] [PubMed]
- Amato, G.S.; Manke, A.; Harris, D.L.; Wiethe, R.W.; Vasukuttan, V.; Snyder, R.W.; Lefever, T.W.; Cortes, R.; Zhang, Y.; Wang, S.; et al. Blocking Alcoholic Steatosis in Mice with a Peripherally Restricted Purine Antagonist of the Type 1 Cannabinoid Receptor. J. Med. Chem. 2018, 61, 4370–4385. [Google Scholar] [CrossRef] [PubMed]
- De Gottardi, A.; Spahr, L.; Ravier-Dall’Antonia, F.; Hadengue, A. Cannabinoid receptor 1 and 2 agonists increase lipid accumulation in hepatocytes. Liver Int. 2010, 30, 1482–1489. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Kim, Y.H.; Jang, H.H.; Park, J.; Kim, J.R.; Koh, M.; Jeong, W.I.; Koo, S.H.; Park, T.S.; Yun, C.H.; et al. Estrogen-related receptor gamma controls hepatic CB1 receptor-mediated CYP2E1 expression and oxidative liver injury by alcohol. Gut 2013, 62, 1044–1054. [Google Scholar] [CrossRef]
- Wang, S.; Pacher, P.; De Lisle, R.C.; Huang, H.; Ding, W.X. A Mechanistic Review of Cell Death in Alcohol-Induced Liver Injury. Alcohol Clin. Exp. Res. 2016, 40, 1215–1223. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, T.; Xu, F.; Li, Y.; Wu, M.; Zhu, D.; Cong, X.; Liu, Y. How important is differentiation in the therapeutic effect of mesenchymal stromal cells in liver disease? Cytotherapy 2014, 16, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.; Zhang, N.; You, N.; Li, Q.; Liu, W.; Jiang, N.; Liu, J.; Zhang, H.; Wang, D.; Tao, K.; et al. The differentiation of MSCs into functional hepatocyte-like cells in a liver biomatrix scaffold and their transplantation into liver-fibrotic mice. Biomaterials 2012, 33, 8995–9008. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Ringden, O. Mesenchymal stem cells: Properties and role in clinical bone marrow transplantation. Curr. Opin. Immunol. 2006, 18, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Mohamadnejad, M.; Namiri, M.; Bagheri, M.; Hashemi, S.M.; Ghanaati, H.; Zare Mehrjardi, N.; Kazemi Ashtiani, S.; Malekzadeh, R.; Baharvand, H. Phase 1 human trial of autologous bone marrow-hematopoietic stem cell transplantation in patients with decompensated cirrhosis. World J. Gastroenterol. 2007, 13, 3359–3363. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.O.; Kim, Y.J.; Baik, S.K.; Kim, M.Y.; Eom, Y.W.; Cho, M.Y.; Park, H.J.; Park, S.Y.; Kim, B.R.; Kim, J.W.; et al. Histological improvement following administration of autologous bone marrow-derived mesenchymal stem cells for alcoholic cirrhosis: A pilot study. Liver Int. 2014, 34, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Puglisi, M.A.; Tesori, V.; Lattanzi, W.; Piscaglia, A.C.; Gasbarrini, G.B.; D’Ugo, D.M.; Gasbarrini, A. Therapeutic implications of mesenchymal stem cells in liver injury. J. Biomed. Biotechnol. 2011, 2011, 860578. [Google Scholar] [CrossRef]
- Meier, R.P.; Muller, Y.D.; Morel, P.; Gonelle-Gispert, C.; Buhler, L.H. Transplantation of mesenchymal stem cells for the treatment of liver diseases, is there enough evidence? Stem Cell Res. 2013, 11, 1348–1364. [Google Scholar] [CrossRef]
- Seitz, H.K.; Mueller, S. Established therapies and new therapeutic strategies in alcoholic liver disease. Z Gastroenterol 2017, 55, 291–303. [Google Scholar]
- Parekkadan, B.; van Poll, D.; Megeed, Z.; Kobayashi, N.; Tilles, A.W.; Berthiaume, F.; Yarmush, M.L. Immunomodulation of activated hepatic stellate cells by mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2007, 363, 247–252. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Jiang, H.; Su, S.; Wang, D.; Liang, Z.; Zhang, J.; Yang, W. Inhibition of hepatic stellate cell proliferation by bone marrow mesenchymal stem cells via regulation of the cell cycle in rat. Exp. Med. 2012, 4, 375–380. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Chen, B.; Chen, L.J.; Zhang, C.F.; Xiang, C. Current status and future prospects of mesenchymal stem cell therapy for liver fibrosis. J. Zhejiang Univ. Sci. B 2016, 17, 831–841. [Google Scholar] [CrossRef] [PubMed]
- An, L.; Lang, Q.; Shen, W.; Shi, Q.; Feng, F. Dynamic metabolic profiling of urine biomarkers in rats with alcoholinduced liver damage following treatment with ZhiZiDaHuang decoction. Mol. Med. Rep. 2016, 14, 2093–2100. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Liu, T.; Zhang, L.; Xing, L.J.; Zheng, P.Y.; Ji, G. Chinese medicinal formula, Qinggan Huoxue Recipe protects rats from alcoholic liver disease via the lipopolysaccharide-Kupffer cell signal conduction pathway. Exp. Med. 2014, 8, 363–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Luo, Y.; Deng, H.; Qin, S.; Tang, W.; Zeng, L.; Zhou, B. Hugan Qingzhi medication ameliorates hepatic steatosis by activating AMPK and PPARalpha pathways in L02 cells and HepG2 cells. J. Ethnopharmacol. 2014, 154, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.M.; Shaw, L.H.; Chang, P.J.; Tung, S.Y.; Chang, T.S.; Shen, C.H.; Hsieh, Y.Y.; Wei, K.L. Hepatoprotective effect of resveratrol against ethanol-induced oxidative stress through induction of superoxide dismutase in vivo and in vitro. Exp. Med. 2016, 11, 1231–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.Q.; Fan, Z.Q.; Tang, Y.F.; Ke, Z.J. The resveratrol attenuates ethanol-induced hepatocyte apoptosis via inhibiting ER-related caspase-12 activation and PDE activity in vitro. Alcohol. Clin. Exp. Res. 2014, 38, 683–693. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, Y.; Li, Q.; Xu, M.; Bai, J.; Wu, S. Resveratrol improves alcoholic fatty liver disease by downregulating HIF-1alpha expression and mitochondrial ROS production. PLoS ONE 2017, 12, e0183426. [Google Scholar] [CrossRef]
- Chen, L.Y.; Chen, Q.; Cheng, Y.F.; Jin, H.H.; Kong, D.S.; Zhang, F.; Wu, L.; Shao, J.J.; Zheng, S.Z. Diallyl trisulfide attenuates ethanol-induced hepatic steatosis by inhibiting oxidative stress and apoptosis. Biomed. Pharm. 2016, 79, 35–43. [Google Scholar] [CrossRef]
- Chen, L.Y.; Chen, Q.; Zhu, X.J.; Kong, D.S.; Wu, L.; Shao, J.J.; Zheng, S.Z. Diallyl trisulfide protects against ethanol-induced oxidative stress and apoptosis via a hydrogen sulfide-mediated mechanism. Int. Immunopharmacol. 2016, 36, 23–30. [Google Scholar] [CrossRef]
- Guan, M.J.; Zhao, N.; Xie, K.Q.; Zeng, T. Hepatoprotective effects of garlic against ethanol-induced liver injury: A mini-review. Food Chem. Toxicol. 2018, 111, 467–473. [Google Scholar] [CrossRef]
- Zeng, T.; Zhang, C.L.; Song, F.Y.; Zhao, X.L.; Yu, L.H.; Zhu, Z.P.; Xie, K.Q. The activation of HO-1/Nrf-2 contributes to the protective effects of diallyl disulfide (DADS) against ethanol-induced oxidative stress. Biochim. Biophys. Acta. 2013, 1830, 4848–4859. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.J.; Truong, V.L.; Ko, S.Y.; Nguyen, X.N.; Ingkasupart, P.; Jun, M.; Shin, J.Y.; Jeong, W.S. Antioxidant and Hepatoprotective Effects of Procyanidins from Wild Grape (Vitis amurensis) Seeds in Ethanol-Induced Cells and Rats. Int. J. Mol. Sci. 2016, 17, 758. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Chen, C.; Wang, J.; Xie, W.; Wang, M.; Li, X.; Zhang, X. Purple potato (Solanum tuberosum L.) anthocyanins attenuate alcohol-induced hepatic injury by enhancing antioxidant defense. J. Nat. Med. 2016, 70, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Chen, C.; Xie, W.; Wang, M.; Wang, J.; Zhang, X. Anthocyanins attenuate alcohol-induced hepatic injury by inhibiting pro-inflammation signalling. Nat. Prod. Res. 2016, 30, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Su, B.; Fan, S.; Fei, H.; Zhao, W. Protective effect of oligomeric proanthocyanidins against alcohol-induced liver steatosis and injury in mice. Biochem. Biophys. Res. Commun. 2015, 458, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Stice, C.P.; Liu, C.; Aizawa, K.; Greenberg, A.S.; Ausman, L.M.; Wang, X.D. Dietary tomato powder inhibits alcohol-induced hepatic injury by suppressing cytochrome p450 2E1 induction in rodent models. Arch. Biochem. Biophys. 2015, 572, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Zhang, W.; Zhong, W.; Sun, X.; Zhou, Z. Dietary Fisetin Supplementation Protects Against Alcohol-Induced Liver Injury in Mice. Alcohol Clin. Exp. Res. 2016, 40, 2076–2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, X.; Deng, Z.B.; Mu, J.; Zhang, L.; Yan, J.; Miller, D.; Feng, W.; McClain, C.J.; Zhang, H.G. Ginger-derived nanoparticles protect against alcohol-induced liver damage. J. Extracell. Vesicles 2015, 4, 28713. [Google Scholar] [CrossRef] [PubMed]
- Varga, Z.V.; Matyas, C.; Erdelyi, K.; Cinar, R.; Nieri, D.; Chicca, A.; Nemeth, B.T.; Paloczi, J.; Lajtos, T.; Corey, L.; et al. beta-Caryophyllene protects against alcoholic steatohepatitis by attenuating inflammation and metabolic dysregulation in mice. Br. J. Pharm. 2018, 175, 320–334. [Google Scholar] [CrossRef]
- Xiao, J.; Zhang, R.; Huang, F.; Liu, L.; Deng, Y.; Ma, Y.; Wei, Z.; Tang, X.; Zhang, Y.; Zhang, M. Lychee (Litchi chinensis Sonn.) Pulp Phenolic Extract Confers a Protective Activity against Alcoholic Liver Disease in Mice by Alleviating Mitochondrial Dysfunction. J. Agric. Food Chem. 2017, 65, 5000–5009. [Google Scholar] [CrossRef]
- Han, J.Y.; Lee, S.; Yang, J.H.; Kim, S.; Sim, J.; Kim, M.G.; Jeong, T.C.; Ku, S.K.; Cho, I.J.; Ki, S.H. Korean Red Ginseng attenuates ethanol-induced steatosis and oxidative stress via AMPK/Sirt1 activation. J. Ginseng. Res. 2015, 39, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.F.; Sun, L.P.; Yang, Q.H.; Lu, D.X.; Luo, S. Ginsenosides from stems and leaves of ginseng prevent ethanol-induced lipid accumulation in human L02 hepatocytes. Chin. J. Integr. Med. 2017, 23, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.G.; Ji, D.F.; Zhong, S.; Shi, L.G.; Hu, G.Y.; Chen, S. Saponins from Panax japonicus protect against alcohol-induced hepatic injury in mice by up-regulating the expression of GPX3, SOD1 and SOD3. Alcohol Alcohol. 2010, 45, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Wo, L.; Du, Z.; Tang, L.; Song, Z.; Dou, X. Danshen protects against early-stage alcoholic liver disease in mice via inducing PPARalpha activation and subsequent 4-HNE degradation. PLoS ONE 2017, 12, e0186357. [Google Scholar] [CrossRef] [PubMed]
- Yue, S.; Hu, B.; Wang, Z.; Yue, Z.; Wang, F.; Zhao, Y.; Yang, Z.; Shen, M. Salvia miltiorrhiza compounds protect the liver from acute injury by regulation of p38 and NFkappaB signaling in Kupffer cells. Pharm. Biol. 2014, 52, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Hu, Y.; Ding, C.; Zeng, W.; Shan, W.; Fan, H.; Zhao, Y.; Shi, X.; Gao, L.; Xu, T.; et al. Salvianolic acid B protects against chronic alcoholic liver injury via SIRT1-mediated inhibition of CRP and ChREBP in rats. Toxicol. Lett. 2017, 267, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kang, O.H.; Kim, S.B.; Mun, S.H.; Seo, Y.S.; Hwang, H.C.; Lee, Y.M.; Lee, H.S.; Kang, D.G.; Kwon, D.Y. Puerarin ameliorates hepatic steatosis by activating the PPARalpha and AMPK signaling pathways in hepatocytes. Int. J. Mol. Med. 2015, 35, 803–809. [Google Scholar] [CrossRef]
- Xiong, Y.; Yang, Y.; Yang, J.; Chai, H.; Li, Y.; Yang, J.; Jia, Z.; Wang, Z. Tectoridin, an isoflavone glycoside from the flower of Pueraria lobata, prevents acute ethanol-induced liver steatosis in mice. Toxicology 2010, 276, 64–72. [Google Scholar] [CrossRef]
- Zhao, Y.Z.; Zhang, L.; Gupta, P.K.; Tian, F.R.; Mao, K.L.; Qiu, K.Y.; Yang, W.; Lv, C.Z.; Lu, C.T. Using PG-Liposome-Based System to Enhance Puerarin Liver-Targeted Therapy for Alcohol-Induced Liver Disease. Aaps. Pharmscitech. 2016, 17, 1376–1382. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Lian, N.; Bian, M.; Zhang, C.; Chen, X.; Shao, J.; Wu, L.; Chen, A.; Guo, Q.; Zhang, F.; et al. Oroxylin A prevents alcohol-induced hepatic steatosis through inhibition of hypoxia inducible factor 1alpha. Chem. Biol. Interact. 2018, 285, 14–20. [Google Scholar] [CrossRef]
- Li, H.D.; Chen, X.; Yang, Y.; Huang, H.M.; Zhang, L.; Zhang, X.; Zhang, L.; Huang, C.; Meng, X.M.; Li, J. Wogonin attenuates inflammation by activating PPAR-gamma in alcoholic liver disease. Int. Immunopharmacol. 2017, 50, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Y.; Bai, R.; Wang, M.; Du, S. Baicalin Attenuates Alcoholic Liver Injury through Modulation of Hepatic Oxidative Stress, Inflammation and Sonic Hedgehog Pathway in Rats. Cell. Physiol. Biochem. 2016, 39, 1129–1140. [Google Scholar] [CrossRef] [PubMed]
- Ghoneim, A.I. Effects of curcumin on ethanol-induced hepatocyte necrosis and apoptosis: Implication of lipid peroxidation and cytochrome c. Naunyn. Schmiedebergs. Arch. Pharm. 2009, 379, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Zhang, F.; Xu, W.; Wu, X.; Lian, N.; Jin, H.; Chen, Q.; Chen, L.; Shao, J.; Wu, L.; et al. Curcumin attenuates ethanol-induced hepatic steatosis through modulating Nrf2/FXR signaling in hepatocytes. Iubmb. Life 2015, 67, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Nanji, A.A.; Jokelainen, K.; Tipoe, G.L.; Rahemtulla, A.; Thomas, P.; Dannenberg, A.J. Curcumin prevents alcohol-induced liver disease in rats by inhibiting the expression of NF-kappa B-dependent genes. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284, G321–G327. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Z.; Wang, X.; Qi, D.; Qu, A.; Wang, G. Amelioration of Ethanol-Induced Hepatitis by Magnesium Isoglycyrrhizinate through Inhibition of Neutrophil Cell Infiltration and Oxidative Damage. Mediat. Inflamm. 2017, 2017, 3526903. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Xu, W.; Shao, J.; Zhang, F.; Chen, A.; Zheng, S. Blockade of hedgehog pathway is required for the protective effects of magnesium isoglycyrrhizinate against ethanol-induced hepatocyte steatosis and apoptosis. Iubmb. Life 2017, 69, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.C.; Lee, Y.H.; Kim, S.H.; Kim, K.J.; Kim, K.M.; Oh, S.; Jung, Y.S. Hepatoprotective effect of licorice, the root of Glycyrrhiza uralensis Fischer, in alcohol-induced fatty liver disease. BMC Complement. Altern. Med. 2016, 16, 19. [Google Scholar] [CrossRef]
- Zhang, P.; Ma, D.; Wang, Y.; Zhang, M.; Qiang, X.; Liao, M.; Liu, X.; Wu, H.; Zhang, Y. Berberine protects liver from ethanol-induced oxidative stress and steatosis in mice. Food Chem. Toxicol. 2014, 74, 225–232. [Google Scholar] [CrossRef]
- Lu, C.; Xu, W.; Shao, J.; Zhang, F.; Chen, A.; Zheng, S. Nrf2 Activation Is Required for Ligustrazine to Inhibit Hepatic Steatosis in Alcohol-Preferring Mice and Hepatocytes. Toxicol. Sci. 2017, 155, 432–443. [Google Scholar] [CrossRef]
- Patsenker, E.; Chicca, A.; Petrucci, V.; Moghadamrad, S.; de Gottardi, A.; Hampe, J.; Gertsch, J.; Semmo, N.; Stickel, F. 4-O’-methylhonokiol protects from alcohol/carbon tetrachloride-induced liver injury in mice. J. Mol. Med. (Berl) 2017, 95, 1077–1089. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jung, J.Y.; Jang, E.J.; Jegal, K.H.; Moon, S.Y.; Ku, S.K.; Kang, S.H.; Cho, I.J.; Park, S.J.; Lee, J.R.; et al. Combination of honokiol and magnolol inhibits hepatic steatosis through AMPK-SREBP-1 c pathway. Exp. Biol. Med. (Maywood) 2015, 240, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Lucey, M.R. Liver transplantation for alcoholic liver disease. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 300–307. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| MicroRNAs | Target | Dysregulation | References |
|---|---|---|---|
| miR-203 | Lipin1 | Decreased | [108] |
| miR-122 | Lipid metabolism: HMGCoA, ApoE, MTTP, PGC1a, HIF1A; Fibrosis: P4HA1; Hepatocellular cancer: Igf1R, ADAM10, cyclin G1, KLF6 | Decreased/Increased | [109,110,111] |
| miR-30e | UCP, ATP | Decreased | [112] |
| miR-19b | miR-17-92 | Decreased | [113] |
| miR-497 | Btg2, Yy1 | Decreased | [114] |
| miR-148a | FOXO1, TXNIP | Decreased | [115] |
| miR-217 | SIRT1/Lipin1 | Increased | [116] |
| miR-214 | GSR, CYP450 OR | Increased | [117] |
| miR-132 | Neuroimmune microRNAs | Increased | [111,118] |
| miR-155 | FABP4, LXRα, ACC1, LDLR | Increased | [109,111,118,119] |
| miR-29a; Let-7f; miR-340 | ASH | Increased | [120] |
| miR-17-92 | Fibrosis | Increased | [113] |
| miR-200a | ZER2 | Increased | [121] |
| miR-34a | SIRT1 | Increased | [122] |
| Names | The Sources | Target | References |
|---|---|---|---|
| Resveratrol | Grapes, red wine, peanuts and berries. | HIF-1α, Oxidative stress | [169,170,171] |
| Diallyl trisulfide | Garlic | Nrf-2/HO-1, Steatosis | [172,173,174,175] |
| Anthocyanin | Purple potato, wild grape | Anti-inflammatory, Oxidative stress, Steatosis | [62,176,177,178,179] |
| Tomato powder | Tomato | CYP2E1 | [180] |
| Fisetin | Strawberry, apple, persimmon, lotus root, and onion | NOX4, Adiponectin, AMPK | [181] |
| Ginger-derived nanoparticles | Ginger | Nrf-2 | [182] |
| β-caryophyllene | Plant-derived food additive | Cannabinoid 2 receptors | [183] |
| lychee pulp phenolic | Lychee | Fatty acid β-oxidation, Hepatocyte apoptosis | [184] |
| Names | The Sources | Target | References |
|---|---|---|---|
| Ginsenosides | Ginseng | Steatosis, oxidative stress | [185,186,187] |
| Salvianolic acid | Salvia miltiorrhiza | SIRT1, P38, NF-κB | [188,189,190] |
| Puerarin | Pueraria lobata | PPARα, AMPK | [191,192,193] |
| Baicalin | Scutellaria baicalensis | Hedgehog Pathway, Oxidative stress, Inflammation | [194,195,196] |
| Curcumin | Curcuma longa | NF-κB, Nrf-2, Cytochrome c, Lipid peroxidation | [197,198,199] |
| Glycyrrhizic acid | Glycyrrhiza uralensis | Glutathione, TNF-α | [200,201,202] |
| Berberine | Coptis chinensis | PPARα, HNF4α | [203] |
| Ligustrazine | Ligusticum chuanxiong | Nrf-2, Hepatic steatosis | [204] |
| Honokiol and magnolol | Magnolia officinalis | AMPK/SREBP-1c, CB2 | [205,206] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, L.-Z.; Chandimali, N.; Han, Y.-H.; Lee, D.-H.; Kim, J.-S.; Kim, S.-U.; Kim, T.-D.; Jeong, D.K.; Sun, H.-N.; Lee, D.S.; et al. Pathogenesis, Early Diagnosis, and Therapeutic Management of Alcoholic Liver Disease. Int. J. Mol. Sci. 2019, 20, 2712. https://doi.org/10.3390/ijms20112712
Kong L-Z, Chandimali N, Han Y-H, Lee D-H, Kim J-S, Kim S-U, Kim T-D, Jeong DK, Sun H-N, Lee DS, et al. Pathogenesis, Early Diagnosis, and Therapeutic Management of Alcoholic Liver Disease. International Journal of Molecular Sciences. 2019; 20(11):2712. https://doi.org/10.3390/ijms20112712
Chicago/Turabian StyleKong, Ling-Zu, Nisansala Chandimali, Ying-Hao Han, Dong-Ho Lee, Ji-Su Kim, Sun-Uk Kim, Tae-Don Kim, Dong Kee Jeong, Hu-Nan Sun, Dong Sun Lee, and et al. 2019. "Pathogenesis, Early Diagnosis, and Therapeutic Management of Alcoholic Liver Disease" International Journal of Molecular Sciences 20, no. 11: 2712. https://doi.org/10.3390/ijms20112712