Genomic Instability in Human Lymphocytes from Male Users of Crack Cocaine

Abstract
:1. Introduction
2. Experimental Section
2.1. Population
2.2. Micronucleus Assay
2.3. Comet Assay
2.4. Statistical Analysis
2.5. Ethical Considerations
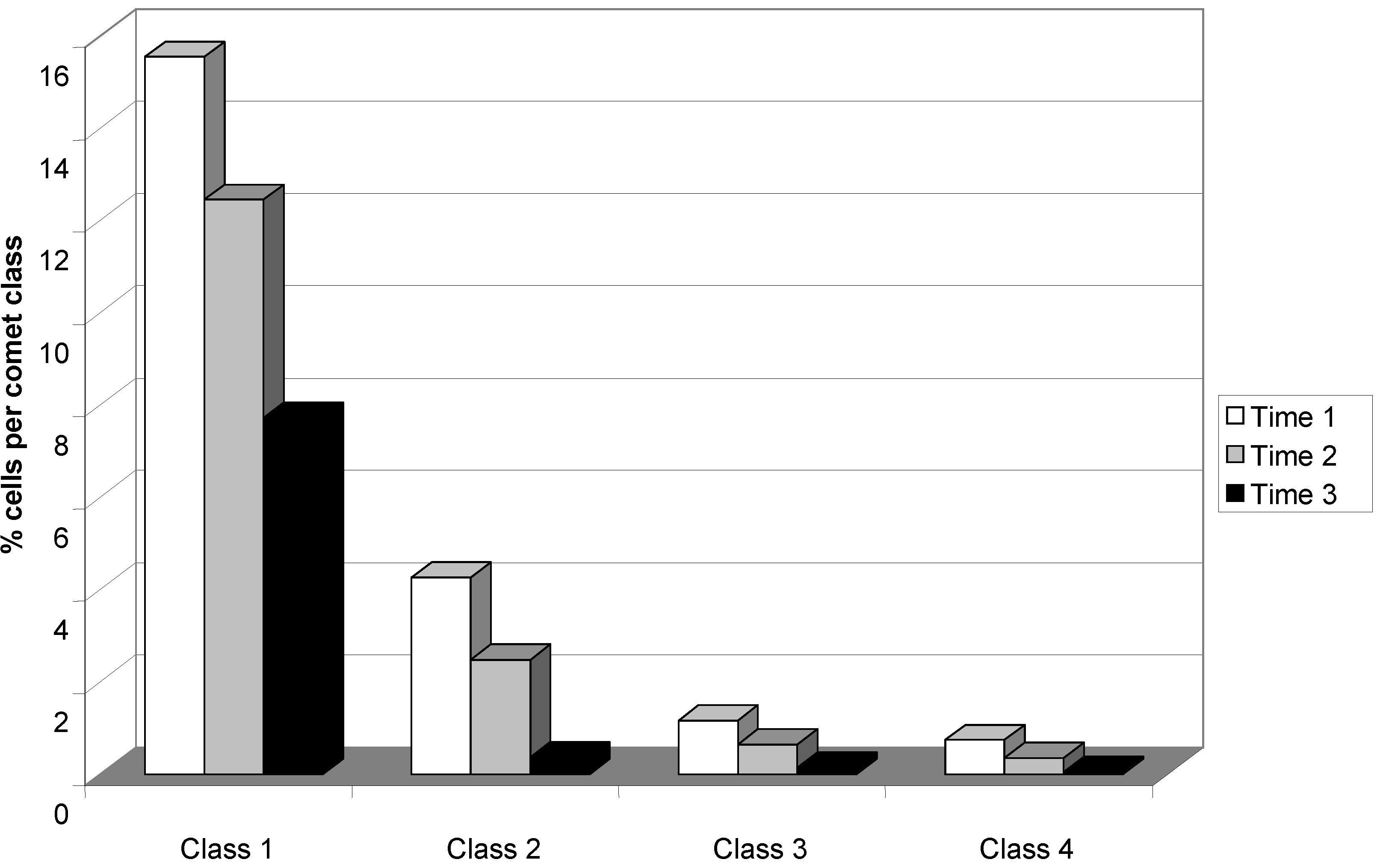
3. Results and Discussion

{kind=link}
{kind=link}
{kind=link}
| Crack Cocaine Users | Controls | |
|---|---|---|
| N * | 31 | 40 |
| Age | 26.13 ± 6.190 | 29.91 ± 9.135 |
| Duration of crack cocaine use (months) | 47.096 ± 30.48 | 0 |
| Doses of crack cocaine/day ** | 12.98 ± 16.69 | 0 |
| Last use before first blood sampling | 66.7 ± 24.63 | 0 |
| Cocaine use | 10 (32.3%) | 0 (0%) |
| Cannabis use | 22 (71%) | 0 (0%) |
| Tobacco use | 21 (67.7%) | 14 (35%) |
| Alcohol intake | 19 (61.3%) | 22 (55%) |
| Ilex paraguariensis infusion | 13 (41.9%) | 31 (77,5%) |
| Controls | Time Point 1 | Time Point 2 | Time Point 3 | |
|---|---|---|---|---|
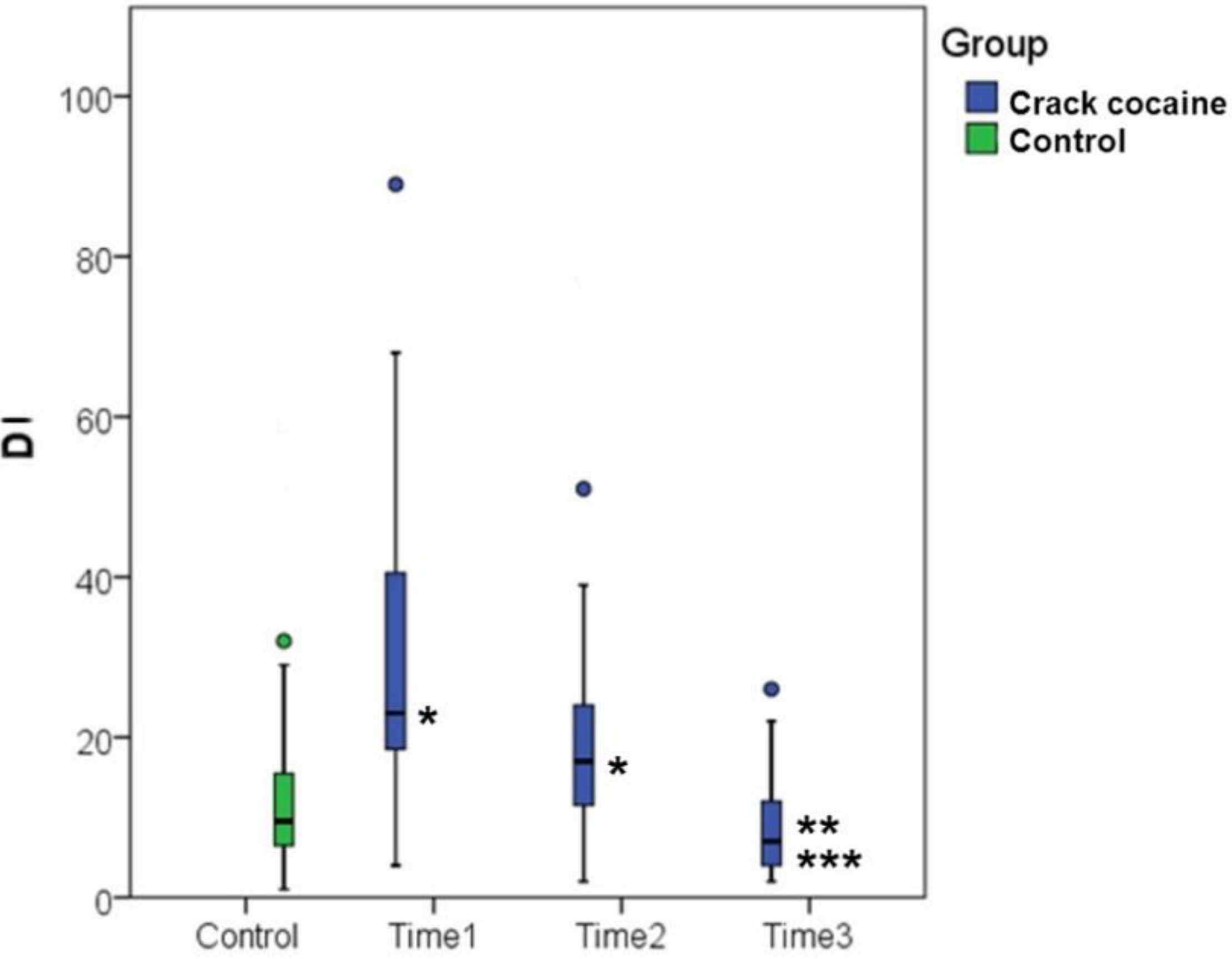
| Comet | 13.05 ± 11.79 (N = 40) | 29.81 ± 19.34 * p < 0.001 (N = 31) | 22.29 ± 19.95 * p = 0.020 (N = 31) | 10.39 ± 8.95 *** p = 0.004 ** p < 0.001 (N = 23) |
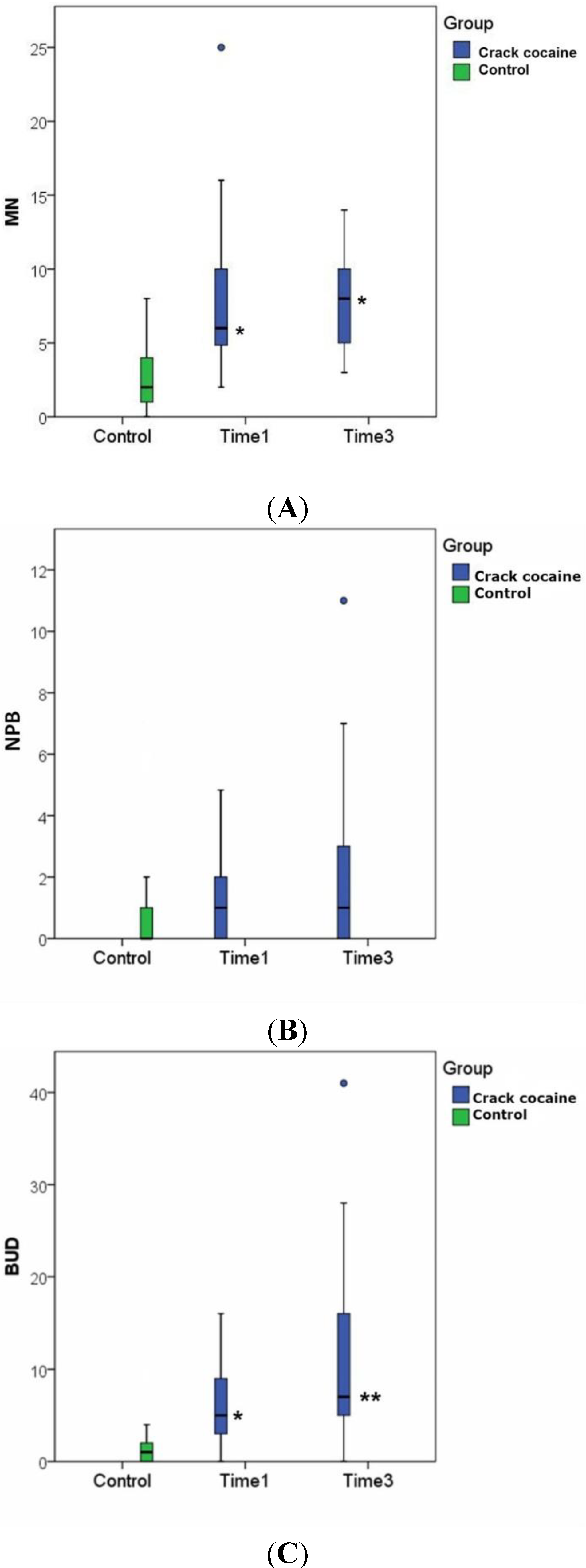
| MN | 2.62 ± 2.26 (N = 29) | 7.67 ± 4.92 * p < 0.001 (N = 29) | - | 7.90 ± 3.32 * p < 0.001 (N = 21) |
| NPBs | 0.76 ± 1.70 (N = 29) | 1.51 ± 1.52 (N = 29) | - | 1.95 ± 2.87 (N = 21) |
| Buds | 1.59 ± 1.88 (N = 29) | 6.27 ± 4.24 * p < 0.001 (N = 29) | - | 16.86 ± 27.56 ** p = 0.009 (N = 21) |



| Correlation | p | N |
|---|---|---|
| Ilex paraguariensis infusion × MN (time point 1) | p = 0.045 | 13 |
| Duration of tobacco use (years) × DI (time point 2) | p = 0.043 | 19 |
| Duration of tobacco use (years) × MN (time point 1) | p = 0.017 | 17 |
| Duration of tobacco use (years) × NPBs (time point 3) | p = 0.023 | 13 |
| Duration of cocaine use × MN (time point 3) | p = 0.028 | 9 |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Falck, R.S.; Wang, J.; Carlson, R.G. Among long-term crack smokers, who avoids and who succumbs to cocaine addiction? Drug Alcohol. Depend 2008, 98, 24–29. [Google Scholar] [CrossRef]
- Devlin, R.D.; Henry, J.A. Clinical review: Major consequences of illicit drug consumption. Crit. Care 2008, 12. [Google Scholar] [CrossRef]
- Riezzo, I.; Fiore, C.; de Carlo, C.; Pascale, N.; Neri, M.; Turillazzi, E.; Fineschi, V. Side effects of cocaine abuse: Multiorgan toxicity and pathological consequences current. Med. Chem. 2012, 19, 5624–5646. [Google Scholar]
- Abdalla, R.R.; Madruga, C.S.; Ribeiro, M.; Pinsky, I.; Caetano, R.; Laranjeira, R. Prevalence of cocaine use in Brazil: Data from the II Brazilian National Alcohol and Drugs Survey (BNADS). Addict. Behav. 2014, 39, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Narvaez, J.C.M.; Magalhães, P.V.; Fries, G.R.; Colpo, G.D.; Czepielewski, L.S.; Vianna, P.; Chies, J.A.B.; Rosa, A.R.; Diemen, L.V.; Vieta, E.; et al. Peripheral toxicity in crack cocaine use disorders. Neurosci. Lett. 2013, 544, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Sordi, A.O.; Pechansky, F.; Kessler, F.H.; Kapczinski, F.; Pfaffenseller, B.; Gubert, C.; de Aguiar, B.W.; Narvaez, J.C.M.; von Diemen, L. Oxidative stress and BDNF as possible markers for the severity of crack cocaine use in early withdrawal. Psychopharmacology 2014, 3. [Google Scholar] [CrossRef]
- Moritz, F.; Monteil, C.; Isabelle, M.; Bauer, F.; Renet, S.; Mulder, P.; Richard, V.; Thuillez, C. Role of reactive oxygen species in cocaine-induced cardiac dysfunction. Cardiovasc. Res. 2003, 59, 834–843. [Google Scholar] [CrossRef] [PubMed]
- Bashkatova, V.; Meunier, J.; Maurice, T.; Vanin, A. Memory impairments and oxidative stress in the hippocampus of in-utero cocaine-exposed rats. Neuroreport 2005, 16, 1217–1221. [Google Scholar] [CrossRef] [PubMed]
- Stefanidou, M.; Chatziioannou, A.; Livaditou, A.; Rellaki, A.; Alevisopoulos, G.; Spiliopoulou, H.; Koutselinis, A. DNA toxicity of cocaine hydrochloride and cocaine freebase by means of DNA image analysis on Tetrahymena pyriformis. Biol. Pharm. Bull. 2001, 25, 332–334. [Google Scholar] [CrossRef]
- Stefanidou, M.E.; Hatzi, V.I.; Terzoudi, G.I.; Loutsidou, A.C.; Maravelias, C.P. Effect of cocaine and crack on the ploidy status of Tetrahymena pyriformis: A study using DNA image analysis. Cytotechnology 2011, 63, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Stefanidou, M.; Loutsidou, A.C.; Chasapis, C.T.; Spiliopoulou, C.A. Immunotoxicity of cocaine and crack. Curr. Drug Abuse Rev. 2011, 4, 95–97. [Google Scholar] [CrossRef] [PubMed]
- De Souza, M.F.; Gonçales, T.A.; Steinmetz, A.; Moura, D.J.; Saffi, J.; Gomez, R.; Barros, H.M.T. Cocaine induces DNA damage in distinct brain areas of female rats under different hormonal conditions. Clin. Exp. Pharmacol. Physiol. 2014, 41, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.G.A.; Santos, J.N.; Cury, P.R.; Da Silva, V.H.P.; Oliveira, N.R.C.; Padovani, R.C.; Tucci, A.M.; Ribeiro, D.A. Cytogenetic biomonitoring of oral mucosa cells of crack cocaine users. Environ. Sci. Pollut. Res. 2014, 1. [Google Scholar] [CrossRef]
- Fenech, M.; Morley, A.A. Measurement of micronuclei in lymphocytes. Mutat. Res. 1985, 147, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M.; Crott, J.W. Micronuclei, nucleoplasmic bridges and nuclear buds induced in folic acid deficient human lymphocytes—Evidence for brakage—Fusion—Bridge cycles in the cytokinesis block micronucleus assay. Mutat. Res. 2002, 504, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Maluf, S.; Pra, D.; Friedrisch, J.; Bittar, C.; da Silva, M.; Henriques, J.; Silla, L. Length of treatment and dose as determinants of mutagenicity in sickle cell disease patients treated with hydroxyurea. Environ. Toxicol. Pharmacol. 2009, 27, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.P.; Mccoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Tice, R.R.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, J.C.; Sasaki, Y.F. Single cell gel/comet assay: Guidelines for in vitro and in vivo genetic toxicology testing. Environ. Mol. Mutag. 2000, 35, 206–221. [Google Scholar] [CrossRef]
- Hartmann, A.; Agurell, E.; Beevers, C.; Brendler-Schwaab, S.; Burlinson, B.; Clay, P.; Collins, A.; Smith, A.; Speit, G.; Thybaud, V.; Tice, R.R. Recommendations for conducting the in vivo alkaline Comet assay. Mutagenesis 2003, 18, 45–51. [Google Scholar]
- Nadin, S.B.; Vargas-Roig, L.M.; Ciocca, D.R.A. The silver staining method for single-cell gel assay. J. Histochem. Cytochem. 2001, 49, 1183–1186. [Google Scholar] [CrossRef] [PubMed]
- Maluf, S.W.; Mergener, M.; Dalcanale, L.; Costa, C.C.; Polo, T.; Kayser, M.; Silva, L.B.; Prá, D.; Teixeira, P. DNA damage in peripheral blood of patients with Chronic Obstructive Pulmonary Disease (COPD). Mutat. Res. 2007, 626, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Bagatini, P.; Palazzo, R.; Rodrigues, M.; Costa, C.; Maluf, S.W. Induction and removal of DNA damage in blood leukocytes of patients with type 2 diabetes mellitus undergoing hemodialysis. Mutat. Res. 2008, 657, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Kirsch-Volders, M.; Fenech, M. Inclusion of micronuclei in non-divided mononuclear lymphocytes and necrosis/apoptosis may provide a more comprehensive cytokinesis block micronucleus assay for biomonitoring purposes. Mutagenesis 2001, 16, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M. The in vitro micronucleus technique. Mutat. Res. 2000, 455, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Umegaki, K.; Fenech, M. Nucleoplasmic bridges are a sensitive measure of chromosome rearrangements in the cytokinesis-block micronucleus assay. Mutagenesis 2003, 18, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R. The comet assay for dna damage and repair, principles, applications, and limitations. Mol. Biotechnol. Mar. 2004, 26, 249–261. [Google Scholar] [CrossRef]
- Jiang, W.; Lu, Y.; Chen, Z.; Chen, S.; Zhang, M.; Jin, L.; Luo, J.; He, J. Studying the genotoxicity of vincristine on human lymphocytes using comet assay, micronucleus assay and TCR gene mutation test in vitro. Toxicology 2008, 252, 113–117. [Google Scholar]
- Fenech, M.; Kirsch-Volders, M.; Natarajan, A.T.; Surralles, J.; Crott, J.W.; Parry, J.; Norppa, H.; Eastmond, D.A.; Tucker, J.D.; Thomas, P. Molecular mechanisms of micronucleus, nucleoplasmic bridge and nuclear bud formation in mammalian and human cells. Mutagenesis 2011, 26, 125–132. [Google Scholar] [CrossRef] [PubMed]
- El-Zein, R.A.; Fenech, M.; Lopez, M.S.; Spitz, M.R.; Etzel, C.J. Cancer epidemiology. Biomarkers Prev. May 2008, 17, 1111–1119. [Google Scholar] [CrossRef]
- Fenech, M. Biomarkers of genetic damage for cancer epidemiology. Toxicology 2002, 181–182, 411–416. [Google Scholar]
- Alves, R.J.V.; Jotz, G.P.; Amaral, V.S.; Montes, T.M.H.; Sampaio, H.; Andrade, H.H.R. The evaluation of mate’ (Ilex paraguariensis) genetic toxicity in human lymphocytes by the cytokinesis-block in the micronucleus assay. Toxicol. Vitro 2008, 22, 695–698. [Google Scholar] [CrossRef]
- Patankar, N.; Vaidya, V.G. Evaluation of genetic toxicity of insecticide phosphamidon using in vitro & in vivo mammalian test systems. Indian J. Exp. Biol. 1980, 18, 1145–1147. [Google Scholar] [PubMed]
- Ahmed, T.; Pathak, R.; Mustafa, M.D.; Kar, R.; Tripathi, A.K.; Ahmed, R.S.; Banerjee, B.D. Ameliorating effect of N-acetylcysteine and curcumin on pesticide-induced oxidative DNA damage in human peripheral blood mononuclear cells. Environ. Monit. Assess. Aug. 2011, 179, 293–299. [Google Scholar] [CrossRef]
- Stefani, E.; Moore, M.; Aune, D.; Deneo-Pellegrini, H.; Ronco, A.L.; Boffetta, P.; Correa, P.; Acosta, G.; Mendilaharsu, M.; Luaces, M.E.; et al. Maté consumption and risk of cancer: A multi-site case-control study in Uruguay. Asian. Pac. J. Cancer Prev. 2001, 12, 1089–1093. [Google Scholar]
- Szymanska, K.; Matos, E.; Hung, R.J.; Wunsch-Filho, V.; Eluf-Neto, J.; Menezes, A.; Daudt, A.W.; Brennan, P.; Boffetta, P. Drinking of mate’ and the risk of cancers of the upper aerodigestive tract in Latin America: A case–control study. Cancer Causes Control 2010, 21, 1799–1806. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freitas, T.A.B.d.; Palazzo, R.P.; De Andrade, F.M.; Reichert, C.L.; Pechansky, F.; Kessler, F.; De Farias, C.B.; De Andrade, G.G.; Leistner-Segal, S.; Maluf, S.W. Genomic Instability in Human Lymphocytes from Male Users of Crack Cocaine. Int. J. Environ. Res. Public Health 2014, 11, 10003-10015. https://doi.org/10.3390/ijerph111010003
Freitas TABd, Palazzo RP, De Andrade FM, Reichert CL, Pechansky F, Kessler F, De Farias CB, De Andrade GG, Leistner-Segal S, Maluf SW. Genomic Instability in Human Lymphocytes from Male Users of Crack Cocaine. International Journal of Environmental Research and Public Health. 2014; 11(10):10003-10015. https://doi.org/10.3390/ijerph111010003
Chicago/Turabian StyleFreitas, Thiago Aley Brites de, Roberta Passos Palazzo, Fabiana Michelsen De Andrade, César Luis Reichert, Flávio Pechansky, Félix Kessler, Caroline Brunetto De Farias, Gisele Gomes De Andrade, Sandra Leistner-Segal, and Sharbel Weidner Maluf. 2014. "Genomic Instability in Human Lymphocytes from Male Users of Crack Cocaine" International Journal of Environmental Research and Public Health 11, no. 10: 10003-10015. https://doi.org/10.3390/ijerph111010003