
Abstract
Aberrant regulation of the Wnt/β-catenin signaling pathway is one of the major causes of colorectal cancer (CRC). Loss-of-function mutations in APC are commonly found in CRC, leading to inappropriate activation of canonical Wnt signaling. Conversely, gain-of-function mutations in KRAS and BRAF genes are detected in up to 60% of CRCs. Whereas KRAS/mitogen-activated protein kinase (MAPK) and canonical Wnt/β-catenin pathways are critical for intestinal tumorigenesis, mechanisms integrating these two important signaling pathways during CRC development are unknown. Results herein demonstrate that transformation of normal intestinal epithelial cells (IECs) by oncogenic forms of KRAS, BRAF or MEK1 was associated with a marked increase in β-catenin/TCF4 and c-MYC promoter transcriptional activities and mRNA levels of c-Myc, Axin2 and Lef1. Notably, expression of a dominant-negative mutant of T-Cell Factor 4 (ΔNTCF4) severely attenuated IEC transformation induced by oncogenic MEK1 and markedly reduced their tumorigenic and metastatic potential in immunocompromised mice. Interestingly, the Frizzled co-receptor LRP6 was phosphorylated in a MEK-dependent manner in transformed IECs and in human CRC cell lines. Expression of LRP6 mutant in which serine/threonine residues in each particular ProlineProlineProlineSerine/ThreonineProline motif were mutated to alanines (LRP6-5A) significantly reduced β-catenin/TCF4 transcriptional activity. Accordingly, MEK inhibition in human CRC cells significantly diminished β-catenin/TCF4 transcriptional activity and c-MYC mRNA and protein levels without affecting β-catenin expression or stability. Lastly, LRP6 phosphorylation was also increased in human colorectal tumors, including adenomas, in comparison with healthy adjacent normal tissues. Our data indicate that oncogenic activation of KRAS/BRAF/MEK signaling stimulates the canonical Wnt/β-catenin pathway, which in turn promotes intestinal tumor growth and invasion. Moreover, LRP6 phosphorylation by ERK1/2 may provide a unique point of convergence between KRAS/MAPK and Wnt/β-catenin signalings during oncogenesis.
Similar content being viewed by others
Introduction
Colorectal cancers (CRCs) develop through a series of well-characterized histopathological changes resulting from specific mutations in selected oncogenes and tumor suppressor genes. At least four sequential genetic changes need to occur to ensure CRC evolution.1 One oncogene, KRAS, as well as the tumor suppressor genes adenomatous polyposis coli (APC), SMAD4 and TP53, are the main targets of these genetic changes. Of note, mutations in the APC gene are responsible for familial adenomatous polyposis and also have a rate-limiting role in the initiation of the majority of sporadic CRCs. The major tumor suppressor function of the APC protein is a negative regulator of Wnt signaling, where it forms part of the β-catenin destruction complex, comprising Axin, GSK3β and CK1. Mutations in APC lead to β-catenin stabilization and, consequently, to the deregulation of the Wnt pathway through the activation of TCF/LEF target genes such as c-MYC.2 Most of genetically modified mice carrying different mutations in the Apc gene3 show an intestinal tumor predisposition phenotype and develop few to many adenomas. Remarkably, c-Myc deletion suppresses all the phenotypes of the Apc tumor suppressor loss and halts intestinal regeneration.4, 5
KRAS is another important and frequently mutated gene during colorectal carcinogenesis. KRAS mutations are found in 35–42% of CRCs and advanced adenomas.6, 7 Genetic and biochemical studies have firmly established the central role of KRAS-dependent signaling in regulating colorectal tumor cell proliferation, growth, survival, invasion and metastasis formation.7, 8, 9 The most studied KRAS effector pathways are the RAF-MEK-ERK mitogen-activated protein kinase (MAPK) and the phosphatidylinositol 3-kinase (PI3K)-AKT effector pathways6, 9 with inhibitors of components of both pathways currently under clinical evaluation.10, 11, 12, 13, 14 As KRAS and BRAF mutations are mutually exclusive in colorectal tumors,15, 16 aberrant activation of BRAF signaling is considered critical for KRAS-mediated colorectal oncogenesis.15
BRAF relays its signals via the MAPK kinases MEK1 and MEK2, which in turn activate ERK1 and ERK2. Activated ERK1/2 then translocate into the nucleus where they phosphorylate and activate many nuclear transcription factors enhancing gene transcription.17 Studies on normal intestinal epithelial cells (IECs) in culture have demonstrated a close correlation between ERK1/2 activation and G1/S phase transition, whereas pharmacological or molecular inhibition of ERK1/2 abrogated cell proliferation.18, 19, 20 Notably, we previously localized activated forms of ERK1/2 in the nucleus of undifferentiated proliferative epithelial cells in the human intestine.18 The involvement of MEK/ERK signaling in intestinal tumorigenesis is supported by a number of observations.20 First, MEK1/2 are phosphorylated and activated in 30–40% of adenomas and 76% of colorectal tumors.21, 22 Second, expression of a constitutively active mutant of MEK1 or MEK2 in rodent normal IECs is sufficient to induce growth in soft agar, epithelial to mesenchymal transition (EMT) and formation of invasive metastatic tumors in nude mice.23, 24, 25, 26 Third, synthetic MEK inhibitors inhibit intestinal polyp growth in ApcMin/+ mice22 and attenuate proliferation of human CRC cells in culture and in mouse xenografts.27 Taken together, these data strongly suggest that MEK/ERK signaling may contribute to colorectal carcinogenesis.20 However, the exact molecular mechanisms by which MEK/ERK signaling achieves such functions in the colon and rectum remain unclear.
Herein, we demonstrate that oncogenic activation of KRAS/BRAF/MEK signaling in IECs activates the canonical Wnt/β-catenin pathway which, in turn, promotes cell migration and invasion as well as tumor growth and metastasis. Moreover, our results indicate that MEK-dependent phosphorylation of the Frizzled co-receptor LRP6 may serve as the link between these two important signaling pathways in CRC.
Results
Oncogenic KRAS and activated MEK1 induce EMT and perturb β-catenin localization
Previous reports have demonstrated that expression of constitutively active mutants of MEK1 (caMEK),24, 25, 26 BRAF28, 29 or KRAS30 in normal IECs such as IEC-6 is sufficient to promote their transformation. As shown in Figure 1, phase-contrast microscopy confirmed that KRASG12V or caMEK-expressing IEC-6 cells had clearly lost their cell–cell contacts and exhibited a fibroblast-like appearance when compared with control cells (Figure 1a). In addition, E-cadherin protein expression was markedly downregulated in KRASG12V and caMEK-transformed cells (Figure 1b) suggesting that these cells have undergone an EMT, as previously detailed.24 As expected, treatment with the MEK inhibitor U0126 for 72 h efficiently rescued the epithelial phenotype of KRASG12V and caMEK-expressing IEC-6 cells (Figure 1a, panels 3 and 6) and marginally increased E-cadherin expression (Figure 1b). As previously reported,23, 24 IEC-6 transformed by either oncogenic KRAS or caMEK did not show marked enhancement of ERK phosphorylation in comprison with control cells. One plausible explanation is that cells permanently stimulated by autoactive KRAS or MEK1 were desensitized via multiple mechanisms including the previously reported ERK-mediated feedback inhibition of MEK and possible increased basal levels of MAPK phosphatases, a phenomenon previously observed in rodent fibroblasts.29, 31, 32, 33

Oncogenic KRAS and activated MEK1 induce EMT and perturb β-catenin localization. (a) Representative phase-contrast microscopy images of IEC-6 cells expressing pBABE (empty vector), KRASG12V, wtMEK or caMEK, and treated or not with 20 μM U0126 during 24 h. (b) Equal amounts of whole-cell lysates were separated by 10% SDS–PAGE and proteins analyzed by western blotting with specific antibodies against E-cadherin, phosphorylated ERK1/2, ERK2 and HA tag. (c) IEC-6 cells stably expressing wtMEK (panels 1–3) or caMEK (panels 4–6), pBABE (panels 7–9) or KRASG12V (panels 10–12) were fixed for immunofluorescence and stained for β-catenin protein (red) and DAPI (blue). Panels 3, 6, 9 and 12. Full overlap of the fluorescence signals (yellow). Representative immunofluorescence images are shown. Bars: 25 μm.
β-Catenin links E-cadherin and α-catenin to the cytoskeleton to form a complex that maintains normal epithelial polarity and intercellular adhesion.34 As E-cadherin is downregulated during EMT, we speculated that β-catenin accumulates in the cytoplasm and translocates into the nucleus in IECs transformed by oncogenic MEK (Figure 1c, panels 4–6) or KRAS (Figure 1c, panels 10–12). As shown in Figure 1c, the control cells (wtMEK, panels 1–3 or pBabe, panels 7–9) showed typical honeycomb cell surface β-catenin staining as observed in the parental IEC-624). By contrast, after expression of activated MEK (Figure 1c, panels 4–6) or oncogenic KRAS (Figure 1c, panels 10–12), cells have changed their morphology and β-catenin decreased from cell–cell contacts, increasing in both the cytoplasm and in the nucleus (Figure 1c, panels 6 and 12).
Induction of β-catenin/TCF complex transcriptional activity in IECs transformed by oncogenic KRAS or MEK1
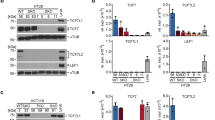
As nuclear β-catenin staining was observed in IECs transformed by oncogenic KRAS or activated MEK1, possible modulation of β-catenin/TCF transcriptional activity was investigated. Cells were thereby transfected with the TOPFLASH reporter, which directly assays β-catenin/TCF activity. As shown in Figure 2a, β-catenin/TCF4 transcriptional activity was significantly enhanced in cells transformed by oncogenic KRAS or MEK1 in comparison with their respective controls. Transcriptional activation of c-Myc is often used as a Wnt readout, as its promoter contains β-catenin/TCF response elements.35 Indeed, similarly to TOPFLASH activity, the c-Myc promoter activity was significantly increased in KRASG12V and caMEK-transformed IECs (Figure 2b). Quantitative PCR (QPCR) and western blot analyses confirmed the significant MEK-dependent induction of c-Myc mRNA and protein levels in cells transformed by activated MEK1 or KRASG12V (Figures 2c, d). Moreover, QPCR analyses demonstrated that oncogenic KRAS or MEK1 increased the expression of Axin2 and Lef1 genes (Figures 2e and f), two other transcriptional targets of the β-catenin/TCF complex.36 These inductions were again dependent on MEK activity, as they were abrogated by U0126. These data indicate that the nuclear β-catenin/TCF complex activity is activated upon oncogenic stimulation of KRAS/MAPK signaling in IECs.

Induction of β-catenin/TCF complex transcriptional activity in IECs transformed by oncogenic KRAS or MEK1. (a, b) IEC-6 cells stably expressing pBABE, KRASG12V, wtMEK or caMEK were transfected with 0.3 μg of TOPFLASH/FOPFLASH reporter genes (a) or c-myc/c-mut (4 × TBE2-wt/4 × TBE2-mut) luciferase reporters (b). Thirty-six hours after transfection, cells were lysed and luciferase activity was measured. The increase in luciferase activity was calculated relative to the level observed in pBABE-expressing cells, which was set at 1. Values were also normalized with Renilla-luciferase vector. Results are the mean±s.e. of at least three separate experiments. Significantly different from respective control at *P<0.05; **P<0.01; or ***P<0.001 (Student’s t-test). (c, d) Cells expressing pBABE, KRASG12V, wtMEK or caMEK were treated or not with 20 μM U0126 during 24 h. Thereafter, cells were lyzed and mRNA were analyzed with quantitative real-time PCR for expression of c-Myc (c) and proteins were analyzed by western blotting for the expression of c-Myc, phosphorylated ERK1/2 and total ERK2 (d). (e, f) Cells expressing pBABE, KRASG12V, wtMEK or caMEK were treated or not with 20 μM U0126 during 24 h. Thereafter, cells were lyzed and mRNA analyzed with quantitative real-time PCR for expression of Axin2 and Lef1.
Attenuation of caMEK-driven morphological transformation of IECs occurs upon interference with the β-catenin/TCF4 complex
To test the hypothesis that the activation of the β-catenin/TCF4 complex is required for transformation induced by oncogenic KRAS/MEK signaling, a dominant-negative TCF4 mutant (ΔNTCF4) was overexpressed in cells transformed by activated MEK1. This NH2 terminally deleted mutant lacks the β-catenin-interaction domain and has been shown to interfere with the activity of the endogenous, constitutively active, β-catenin/TCF4 complex present in CRC cells.37, 38 Accordingly, caMEK-expressing cells transfected with the ΔNTCF4 construct exhibited significant attenuation of the transcriptional activity of the β-catenin/TCF complex and c-Myc promoter (Figure 3a). Furthermore, western blot analyses confirmed the reduction in c-Myc and Fra-1 transcription factors following the expression of the ΔNTCF4 mutant in caMEK cells (Figure 3b). Of note, expression of E-cadherin protein was also partially restored upon expression of the TCF4 mutant (Figure 3b). In addition, when ΔNTCF4 was expressed, the caMEK-transformed cells showed a partial reversion to an epithelial morphology (Figure 3c). Similar results were obtained after treatment of caMEK-transformed cells with ICG-001 (Figure 3d), a small-molecule antagonist of β-catenin/TCF-mediated transcription.39

Attenuation of caMEK-driven morphological transformation of IECs occurs upon interference with the β-catenin/TCF4 complex. (a) Subconfluent IEC-6 wtMEK or caMEK cells stably expressing a dominant-negative form of TCF4 (ΔNTCF4) or the empty vector (EV) were transfected with 0.3 μg of the TOPFLASH/FOPFLASH reporter genes and c-myc/c-mut (4 × TBE2-wt/4 × TBE2-mut) luciferase reporters. Thirty-six hours after transfection, cells were lysed and luciferase activity was measured. The luciferase activity was calculated relative to the level observed in EV-expressing cells, which was set at 100%. Values were also normalized with Renilla-luciferase vector. Results are the mean±s.e. of at least three separate experiments. Significantly different from respective control at *P<0.05 or **P<0.01 (Student’s t-test). (b) Equal amounts of lysates from IEC-6 wtMEK or caMEK cells stably expressing ΔNTCF4 or E.V. were separated by SDS–PAGE, and proteins analyzed by western blotting with specific antibodies against Tcf4, c-Myc, Fra-1, E-cadherin and total ERK2. (c) Representative phase-contrast microscopy images of IEC-6 caMEK expressing ΔNTCF4 or E.V. (as control). Bars: 50 μm. (d) Representative phase contrast microscopy images of IEC-6 caMEK that were treated or not with 7.5 μM ICG-001 during 36 h. Bars: 25 μm.
Expression of ΔNTCF4 inhibits proliferative, tumoral and invasive properties of cells transformed by activated MEK1
To determine the importance of the β-catenin/TCF complex in the oncogenicity of activated MEK1 in IECs, we first assessed the proliferation rate of caMEK cells expressing or not the ΔNTCF4 mutant. As shown in Figure 4a, expression of the TCF4 mutant significantly slowed the proliferation of caMEK-transformed cells without affecting proliferation of control wtMEK-expressing cells. Furthermore, ΔNTCF4 expression strongly diminished the ability of caMEK cells to form colonies in soft agarose (Figure 4b). The effect of ΔNTCF4 expression was also determined on migration and invasion of caMEK-transformed cells in the presence of 20 μM hydroxyurea, known to arrest the cell cycle in the G1/S phase. As shown in Figures 4c and d, ΔNTCF4 expression in caMEK cells significantly reduced their capacity to migrate and to invade Matrigel. The tumorigenicity of these cell populations in vivo was subsequently assessed after subcutaneous injection into the flanks of nude mice. As shown in Figure 4e, caMEK-expressing cells induced palpable tumors with a short latency period of 1 week after injection. Interestingly, co-expression of ΔNTCF4 clearly impaired their capacity to grow as tumors in nude mice. Lastly, we investigated whether ΔNTCF4 expression alters the capacity of caMEK cells to form metastases in an experimental metastasis assay. Nude mice injected with caMEK cells into the tail vein showed extensive lung metastasis within 28 days, whereas caMEK cells expressing ΔNTCF4 exhibited attenuated lung colonization (Figure 4f).

Expression of ΔNTCF4 inhibits proliferative, tumoral and invasive properties of cells transformed by activated MEK1. (a) IEC-6 caMEK cells stably expressing ΔNTCF4 or E.V. (as control) were seeded and the number of cells counted during 7 days. (b) IEC-6 caMEK cells stably expressing ΔNTCF4 or E.V. were cultured in soft agarose for 3 weeks before 3-(4,5-Dimethylthiazol-2-Yl)-2,5-Diphenyltetrazolium Bromide (MTT) staining. The number of colonies was calculated using the Image J software. (c) Invasion capacity of IEC-6 caMEK cells stably expressing ΔNTCF4 or E.V. through Matrigel was studied using Matrigel-coated Transwells during 48 h. Thereafter, cells were fixed and stained with 0.5% crystal violet solution. (d) Migration of IEC-6 caMEK cells stably expressing ΔNTCF4 or E.V. to the undersurface of the polycarbonate membrane of Boyden chambers was evaluated 24 h after seeding, in presence of 20 μM hydroxyurea. The number of cells in c, d was determined in 10 fields, the experiments performed in duplicate and the number of E.V.-expressing cells, which had migrated was set at 100%. Significantly different from respective control at ***P<0.001 (Student’s t-test). (e) Tumor growth over time was measured after subcutaneous injection of 2 × 106 of IEC-6 caMEK cells stably expressing ΔNTCF4 or E.V. The results represent the mean tumor volume obtained from at least six mice injected for each cell line. Independent experiments were performed twice. (f) Representative digital images of mouse lungs 21 days after tail vein injection of 106 IEC-6 caMEK cells expressing E.V. or ΔNTCF4. Similar results were obtained in two independent experiments.
Inhibition of MEK activity in human CRC cell lines significantly reduces β-catenin/TCF complex activity
To evaluate whether KRAS/MAPK signaling regulates the β-catenin/TCF complex in human cell models, we analyzed the impact of MEK inhibitor on the activity of TOPFLASH reporter activity and c-myc expression in two human CRC cell lines, namely HT-29 and DLD-1, which have an activating mutation in the BRAF and KRAS genes, respectively. As shown in Figure 5a, treatment of these CRC cell lines with U0126 significantly reduced TOPFLASH activity as well as c-MYC mRNA levels (Figure 5b), indicating that expression of the β-catenin/TCF complex activity was likely dependent on MEK activity in these cells. Western blot analyses demonstrated that MEK inhibition also reduced c-MYC protein levels in both cell lines without, however, affecting E-cadherin and β-catenin expressions (Figure 5c).

Inhibition of MEK activity in human CRC cell lines significantly reduces β-catenin/TCF complex activity. (a) Subconfluent DLD-1 and HT-29 cells were transfected with 0.3 μg of the TOPFLASH/FOPFLASH luciferase reporter vectors. Twelve hours after transfection, cells were treated or not with 20 μM U0126 during 24 h after which luciferase activity was measured. The luciferase activity was calculated relative to the level observed in dimethylsulphoxide-treated cells, which was set at 1. The luciferase activity was also normalized with Renilla-luciferase vector. Results are the mean±s.e. of at least three separate experiments. Significantly different from untreated cells at *P<0.05; **P<0.01 or ***P<0.0001 (Student’s t-test). (b, c) HT-29 and DLD-1 cells were treated or not with 20 μM U0126 during 16 h after which c-myc mRNA levels were evaluated using quantitative real-time PCR, whereas proteins were analyzed by western blotting with specific antibodies against c-MYC, β-catenin, E-cadherin, phosphorylated ERK1/2 and total ERK2. (d) HT-29 cells were treated during 16 h with 20 μM U0126. Thereafter, cells were fixed for immunofluorescence and stained for β-catenin protein (red) and DAPI (blue). (e, f) Cells were treated during 16 h with 20 μM U0126. Thereafter, 800 μg of cell lysates were immunoprecipitated with nontarget IgG (negative control), anti-TCF4 (e) or anti-E-cadherin (f) antibodies. Proteins from immunoprecipitates were solubilized in Laemmli’s buffer, separated by 7.5% SDS–PAGE and analyzed by western blotting to determine β-catenin association. IP: immunoprecipitation.
To elucidate the molecular mechanisms by which MEK inhibition reduced β-catenin/TCF transcriptional activity in human CRC cells, β-catenin subcellular distribution was verified. As the majority of CRC cells exhibit APC mutations,40 it was therefore expected that these cells would exhibit some nuclear β-catenin staining.41 Indeed, control HT-29 cells showed β-catenin staining predominantly in the cytoplasm and also in the nucleus. However, the nuclear staining of β-catenin was markedly reduced upon MEK inhibition, whereas the membranous signal seemingly accumulated in areas of intercellular contacts (Figure 5d). Similar results were obtained in DLD-1 cells (data not shown). Co-immunoprecipitation assays further demonstrated that the β-catenin/TCF4 association was indeed clearly reduced following MEK inhibition in these cells (Figure 5e). By contrast, we could not detect modified interaction of β-catenin with E-cadherin in U0126-treated cells (Figure 5f).
LRP6 is phosphorylated in a MEK-dependent manner in human CRC cells and in IECs expressing oncogenic KRAS, BRAF or MEK1
As nuclear localization of β-catenin was reduced following MEK inhibition in CRC cells, we first speculated that this could potentially be associated with changes in β-catenin phosphorylation. Indeed, CK1α phosphorylates β-catenin on serine-45, providing a recognition site for GSK3β, allowing GSK3β to phosphorylate threonine-41, serine-37 and serine-3341) leading to β-catenin degradation.42 In addition, phosphorylation of β-catenin at tyrosine-142 has been shown to act as a switch from the transcriptional to the adhesive role of β-catenin.43, 44 Src kinases can also phosphorylate tyrosines-86 and 654 on β-catenin; however, only tyrosine-654 phosphorylation regulates its binding to E-cadherin.45 Finally, both Akt and PKA can phosphorylate β-catenin at serine-552, which is associated with its nuclear transcriptional activity.46, 47, 48 However, by using the phospho-specific antibodies, we found that U0126 treatment did not alter β-catenin phosphorylation on these various phosphorylation sites (Figure 6a).

LRP6 is phosphorylated in a MEK-dependent manner in human CRC cells and in IEC-6 expressing oncogenic KRAS, BRAF or MEK1. (a, b) DLD-1 and HT-29 cells were treated or not (DMSO) with 20 μM U0126 during 16 h and equal amounts of cell lysates were separated by SDS–PAGE. In a, proteins were analyzed by western blotting for expression of β-catenin phosphorylated on serine-552, tyrosine-86, tyrosine-654 and tyrosine-142 with phospho-specific antibodies. In addition, β-catenin unphosphorylated on serine-37 and threonine-41 was also analyzed by a specific antibody as well as phosphorylated ERK1/2 and total ERK2. In b, proteins were analyzed by western blotting for expression of total LRP6 and LRP6 phosphorylated on serine-1490 and threonine-1572 as well as phosphorylated ERK1/2 and total ERK2. (c) Equal amounts of lysates from IEC-6 pBABE, KRASG12V, wtMEK and caMEK expressing cells treated or not with 20 μM U0126 during 24 h were analyzed by western blotting for the expression of total ERK2, phosphorylated ERK1/2, total Lrp6 and Lrp6 phosphorylated on serine-1490 and threonine-1572. (d) IEC-6 BRAFV600EER cells were stimulated or not with 250 nM 4-OH tamoxifen in presence or absence of MEK inhibitors (20 μM U0126; 2 μM PD184352) at the indicated times. Proteins were analyzed by western blotting for the expression of total ERK2, phosphorylated ERK1/2, c-Myc, β-actin, total Lrp6 and Lrp6 phosphorylated on threonine-1572 or serine-1490. (e) Mucosal enrichments from 4-week-old BRafIEC-KO and control murine colons were analyzed by western blotting for the expression of phosphorylated ERK1/2 (pERK), ERK2, total Lrp6 and Lrp6 phosphorylated on threonine-1572 or serine-1490. Five mice per group were analyzed and representative western blot analysis of two mice per group is shown.
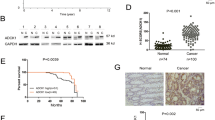
Upon examining the Wnt/β-catenin pathway for components that could be modulated by the MEK/ERK pathway, the Wnt co-receptor LRP6 was found to be phosphorylated on serine-1490 (S1490) and threonine-1572 (T1572) in a MEK-dependent manner in both CRC cells (Figure 6b) and IECs transformed by oncogenic KRAS and MEK1 (Figure 6c). Indeed, ERK1/2 have recently been shown to activate the Wnt/β-catenin signaling via phosphorylation of LRP6 on these sites.49 Lrp6 phosphorylation was also markedly enhanced in IEC-6 cells transformed by the inducible BRAF:ER fusion protein (induction by 4-hydroxytamoxifen). Of note, the induction of phosphorylation was detected within 10 min following ERK activation (Figure 6d), indicating that Lrp6 phosphorylation is an early and direct event occurring following the activation of BRAF signaling. Again, treatment with MEK inhibitors, U0126 or PD184352, significantly decreased Lrp6 phosphorylation induced by oncogenic BRAF. To analyze the potential involvement of the BRAF/ERK signaling pathway in the deregulation of Lrp6 in vivo, we used BRafCA mice carrying a Cre-activated allele of the mouse BRaf gene. These mice express normal BRaf before Cre-mediated recombination after which BRafV600E is expressed at physiological levels.50 These mice were crossed with Villin-Cre transgenic mice, expressing the transgene around embryonic day 15.5 (E15.5) only in IECs, therefore, resulting in the generation of BRafIEC-CA mice. As shown in Figure 6e, western blot analysis of colonic epithelial enrichments showed that phosphorylation of ERK1/2 and Lrp6 was increased in BRafIEC-CA mice in comparison with control littermates.
To confirm that Lrp6 protein was directly phosphorylated by ERK1/2, we used the new specific inhibitor of ERK1/2, SCH772984.51 We observed that treatment with this inhibitor reduced Lrp6 phosphorylation in both IEC-6 transformed by KRasG12V or activated MEK1 (Supplementary Figures S1A, B and D). As expected, SCH772984 clearly inhibited ERK1/2 activity as visualized by sustained decreased phosphorylation of Fra-1, a substrate for ERK1/2.52 Of note, this inhibitor did not prevent long-term ERK1/2 re-phosphorylation as previously reported.51 We have also tested other kinase inhibitors on Lrp6 phosphorylation. Intriguingly, JNK inhibitor (SP600125) apparently decreased Lrp6 phosphorylation in KRAS-transformed cells (Supplementary Figure S1D). However, reduction in Lrp6 phosphorylation seems to be a secondary effect of reduced expression of Lrp6. Indeed, treatment with this inhibitor consistently reduced total Lrp6 expression in all cell lines analyzed by more than 50% as revealed by our densitometric analyses (Supplementary Figure S1D). This suggests that JNK activity may regulate Lrp6 expression (not phosphorylation). Of note, GSK3 inhibition by SB216763 did not alter Lrp6 phosphorylation nor expression (Supplementary Figure S1D).
Finally, we have verified if altered expression of secreted agonists/antagonists of Wnt signalling might be involved in the oncogenic action of the KRAS/MAPK pathway. We therefore used LGK974, an inhibitor of the Wnt-specific acyltransferase porcupine. This inhibitor blocks Wnt secretion and has been shown to potently inhibit Wnt signaling in vitro and in vivo.53 We used the inhibitor at 500 nM, a concentration previously described to abrogate Wnt secretion and signaling.53, 54 However, treatment of caMEK-transformed cells or BRAF:ER cells (stimulated or not with tamoxifen) with LGK964 did not alter their transformed morphology nor Lrp6 phosphorylation (Supplementary Figures S2A–C). Similar results were obtained when we treated the cells with DKK-1, an extracellular antagonist of Lrp6 (Supplementary Figures S2A–C). This suggests that Wnt secretion did not contribute to induce Lrp6 phosphorylation and morphological transformation upon oncogenic activation of the BRAF/MEK/ERK pathway.
Oncogenic KRAS signaling triggers β-catenin/TCF4 complex activation via Lrp6 phosphorylation
To analyze whether MEK-dependent Lrp6 phosphorylation was responsible for the increased β-catenin/TCF4 activity observed in cells transformed by oncogenic activation of KRAS signalling, IECs transformed by oncogenic KRAS were transfected with an increasing amount of wild-type LRP6 or LRP6–5A mutant, a mutant in which its serine/threonine residues in each particular PPPS/TP motif was replaced by alanine. As illustrated in Figure 7a, expression of wild-type LRP6 increased TOPFLASH luciferase activity in KRAS-transformed cells in a dose-dependent manner. By contrast, expression of the LRP6-5A mutant reduced TOPFLASH activity, acting in a dominant-negative manner. Similarly, LRP6-5A abrogated BRAFV600E-induced TOPFLASH activity following tamoxifen stimulation of IEC-6 cells expressing the BRAF:ER fusion protein (Figure 7b).

Oncogenic KRAS signaling triggers β-catenin/TCF4 complex activation via LRP6 phosphorylation. (a) IEC-6 KRASG12Vcells were co-transfected with 0.3 μg of luciferase TOPFLASH/FOPFLASH reporters and increasing concentrations of plasmids expressing or not (E.V., empty vector), wild-type LRP6 or LRP6-5A mutant. Twenty-four hours after transfection, cells were lysed and luciferase activity was measured. The increase in luciferase activity was calculated relative to the level observed in E.V.-expressing cells, which was set at 1. Values were also normalized with Renilla-luciferase vector. Results are the mean±s.e. of at least three separate experiments. (b) IEC-6 BRAFV600EER were co-transfected with 0.3 μg of luciferase TOPFLASH/FOPFLASH reporters and 0.4 μg of plasmids expressing or not (E.V., empty vector) wild-type LRP6 or LRP6-5A mutant. Twenty-four hours after transfection, cells were stimulated or not with 250 nM tamoxifen for an additional 24 h after which luciferase activity was measured as described in a. Significantly different from untreated cells at *P<0.05; **P<0.01 or ***P<0.0001 (Student’s t-test). (c) caMEK-expressing cells were stably infected with lentiviruses encoding for a control shRNA (scrambled sequence, shControl) or encoding Lrp6-specific shRNAs (shLrp6A, B or C). After selection, stable table cell populations were lysed and protein lysates were analyzed by western blot for Lrp6 and β-actin protein expression. (d) Cell populations were cultured in soft agarose for 3 weeks before MTT staining. The number of colonies was determined using the ImageJ software. Results are the mean±s.e. of at least three independent experiments. ***, significantly different from shControl cells at P<0.05 (Student's t-test). ****P<0.005.
These results prompted us to analyze Lrp6 contribution in transformation induced by activated MEK/ERK signaling. Because anchorage-independent growth potential may correlate better with tumorigenic growth in vivo, we determined whether Lrp6 inhibition correlated with the inhibition of tumor cell growth in soft agar rather than on plastic. As shown in Figures 7c, d, Lrp6 silencing significantly inhibited anchorage-independent growth of caMEK-transformed cells.
LRP6 phosphorylation on serine-1490 and threonine-1572 is increased in colorectal tumors
MEK/ERK signaling is thought to be affected in early stages of CRC formation because of frequent mutations in KRAS or BRAF.55 Thus, we first verified the expression and phosphorylation status of LRP6 in human colorectal adenomas, all of which exhibit APC-inactivating mutations (exon 15) in combination with KRAS- (G12D, G13D and Q61H) or BRAF (V600E)-activating mutations. As shown in Figure 8a, all adenomas displayed significantly higher phosphorylation levels of LRP6 on both serine-1490 and threonine-1572 in comparison with their corresponding benign margin. Densitometric analysis confirmed the increased ratio of phosphorylated LRP6 relative to LRP6 expression (Figure 8b). Similar increase in LRP6 phosphorylation was observed in a series of adenocarcinomas (n=53) in comparison with their corresponding normal margins (Figure 8c). Lastly, no significant association was, however, observed between increased LRP6 phosphorylation and tumor stage (Supplementary Table S1). Representative western blot analysis on eight paired adenocarcinomas at different stages is shown in Figure 8d.

LRP6 phosphorylation on serine-1490 and threonine-1572 is increased in colorectal adenomas and adenocarcinomas. (a) Expression of total ERK2, LRP6 and phosphorylated LRP6 on serine-1490 and threonine-1572 was investigated by western blotting in seven paired colorectal adenomas (M: normal margins and A: adenomas). (b) Levels of phosphorylated LRP6 were normalized to the levels of total LRP6 levels in each tissue specimen. Tumor-relative phosphorylated/total LRP6 ratios were matched as reference to its normal samples (set at 1) resulting in a dimensionless value (arbitrary units (AU)). Analyzed by paired t-test and * indicates significantly different from normal margins at P⩽0.05. (c) Expression of LRP6 and phosphorylated LRP6 on serine-1490 and threonine-1572 was further investigated in a series of 53 paired specimens (M: resection margins and AC: primary adenocarcinomas) by western blot. Expression levels of phosphorylated LRP6 on serine 1490 and 1572 were normalized to the intensity β-actin expression and to a reference sample, resulting in a dimensionless value (AU). Densitometry of LRP6 phosphorylation in tumor tissues relative to their matched normal samples was analyzed by paired t-test. Significantly different from healthy resection margins **P⩽0.005 and ***P⩽0.001. (d) Representative immunoblot analysis of total LRP6 and LRP6 phosphorylated on threonine 1572 and serine 1490 performed on protein extracts from eight paired resection margins and advanced adenocarcinomas (AC). Tubulin expression is shown as a control of protein loading.
Discussion
Gain-of-function mutations in KRAS and BRAF genes are detected in up to 60% of colorectal tumors at a relatively early stage of the carcinogenic process.56 The nonoverlapping occurrence of BRAF and KRAS mutations suggests that aberrant BRAF downstream signaling is a critical mechanism for KRAS-mediated oncogenesis in CRC.57 In this regard, expression of a constitutively active mutant of MEK1 or MEK2 in normal IECs is sufficient to induce morphological transformation, anchorage-independent growth and tumorigenicity in mice.23, 24, 25, 26 Importantly, aberrant activation of KRAS/BRAF/MEK signaling in these cells triggers EMT characterized by the loss of epithelial polarity and expression of junctional proteins, particularly E-cadherin (Figure 1).24, 29
The cadherin/catenin-based adhesion system is the major means by which epithelial cells adhere to one another.34 β-catenin, a central structural component of this adhesion complex, also acts as a transcriptional co-activator in the Wnt signaling pathway in epithelial cells.34, 35, 36 Despite this finding, there has been much speculation over whether the cadherin-bound and signal transduction pools of β-catenin are functionally interchangeable.58, 59, 60, 61, 62, 63 In the present study, we show that the nuclear transcriptional activity of β-catenin was enhanced upon sustained oncogenic stimulation of normal IECs by KRAS, BRAF or MEK. Importantly, expression of a dominant-negative TCF4 mutant that inhibits β-catenin/TCF4 transactivation severely attenuated morphological transformation and tumorigenic potential induced by oncogenic MEK1. Thus, these data suggest that KRAS/MAPK signaling may utilize the Wnt/β-catenin pathway to induce transformation in IECs.
In keeping with these results, the transcriptional activity of the β-catenin/TCF complex was reduced by MEK inhibitors in human CRC cell lines, suggesting that MEK activation also potentiates Wnt signaling in CRC. Interestingly, these effects were evident despite the presence of APC mutations. This is consistent with the recent observation that Wnt signaling components are significantly enriched in KRAS-dependent CRC cells compared with KRAS-independent cells, despite both classes having comparable APC mutations.64 Accordingly, Horst et al.65 showed that forced expression of the KRAS oncogene in the Caco-2 CRC cell line leads to nuclear accumulation of β-catenin and increases Wnt activity, whereas blocking the tyrosine kinase receptor EGFR, an activator of MAPK signaling, has the converse effect. In addition, the authors observed colocalization of nuclear β-catenin and phosphorylated ERK staining in primary colon tumor xenografts. Likewise, Phelps et al.66 recently reported that KRAS and RAF1 oncogenes promote nuclear accumulation of β-catenin in APC-deficient cells. Overall, these studies reveal that the KRAS/MAPK pathway can regulate β-catenin signaling in human CRC cells.
Very few studies have explored the molecular mechanisms underlying the enhancement of β-catenin transcriptional activity by oncogenic RAS/MAPK signaling. Oncogenic KRAS signalling could increase the stability of β-catenin through its phosphorylation at serine 552.47, 48 However, we found in CRC cells that MEK inhibition reduced β-catenin accumulation into the nucleus and its interaction with TCF4 without affecting its global expression, its phosphorylation on serine 552 (or residues 37, 41, 86, 142 and 654) or its stabilization (data not shown). On the other hand, although the downregulation of E-cadherin on the cell membrane may contribute to nuclear β-catenin accumulation upon oncogenic activation of KRAS/MAPK signaling in normal IECs (the present study;67), additional mechanisms must likely be involved. Indeed, data from BRAF:ER cells suggest that E-cadherin downregulation is not necessary for Wnt activation by oncogenic MAPK signaling. Indeed, activation of BRAF:ER fusion protein in IEC-6 cells induced Lrp6 phosphorylation within 10 min and β-catenin transcriptional activity (TOPFLASH) within 16–24 h without inducing concomitant downregulation of E-cadherin during this time period (data not shown). Furthermore, in human CRC cells, E-cadherin and β-catenin expression and interaction remained unaltered following MEK inhibition. Interestingly, we observed that the Frizzled co-receptor LRP6 was phosphorylated on serine-1490 and threonine-1572 in a MEK-dependent manner in human CRC cells. Furthermore, MEK-dependent phosphorylation of Lrp6 was also observed upon sustained oncogenic activation of KRAS, BRAF and MEK1 in IECs, thus providing a mechanism integrating KRAS/MAPK and canonical Wnt/β-catenin signalings during intestinal transformation. Of note, sustained activation of ERK signaling by the BRafV600E allele in mice also resulted in increased Lrp6 phosphorylation in the colonic epithelium. Whether this increased Lrp6 phosphorylation was associated with higher Wnt/β-catenin signaling in normal colonocytes will require further analyses. Indeed, conflicting literature exists with regard to the role of MAPK signaling in the activation of the Wnt/β-catenin pathway in normal intestinal epithelial cells. On the one hand, data from different groups68, 69 have shown that the activation of the KRasG12D or BRafV637E alleles in mouse intestinal epithelium induced serrated tumorigenesis without enhancing the Wnt signaling. Wnt signaling induction was, however, found in a substantial part of BRafV637E-induced high-grade tumors.69 On the other hand, Carragher et al.70 demonstrated that crypt hyperplasia induced by the BRafV600E allele was rapidly associated (within 3 days) with Wnt pathway activation as visualized by the accumulation of nuclear β-catenin in crypt cells. Moreover, Phelps et al.66 reported that KRas or BRaf activation is needed for the nuclear translocation of β-catenin in colonic adenoma. Therefore, nuclear translocation of β-catenin might be regarded as a potentially RAS/MAPK-controlled step in canonical Wnt signaling.71, 72
Interestingly, both the serine-1490 and threonine-1572 residues are localized within the PPPS/TP motifs of the LRP6 co-receptor, motifs which are required for Wnt/β-catenin signal transduction. Indeed, removal of any of the five PPPS/TP motifs impairs Wnt signaling, whereas removal of all five motifs results in complete loss of signaling.73, 74 Thus, given their importance, the PPPS/TP motifs may represent a major site for modulation of the Wnt/β-catenin pathway by other signaling systems. In this respect, we found that expression of LRP6-5A, a mutant in which its serine/threonine residues in each particular PPPS/TP motif is replaced by alanine (LRP6-5A), markedly reduced TOPFLASH activation in KRASG12V and BRAFV600E-transformed IECs. On the other hand, we found that Lrp6 silencing significantly reduced anchorage-independent growth of caMEK-transformed cells, hence confirming the contribution of this receptor in oncogenic action of KRAS/MAPK signaling in intestinal epithelial cells. Owing to the lack of phospho-specific antibodies, we could not verify whether Lrp5 was also phosphorylated in a MEK-dependent manner. Indeed, we compared LRP5 and LRP6 amino-acid sequences and we found that the cytoplasmic domain of LRP5 also contains serine-1503 and threonine-1578, both residues localized in PPPSPxS motifs. Therefore, we cannot exclude that the LRP5 protein is also targeted by oncogenic KRAS signaling.
Of further interest, we found increased LRP6 phosphorylation on serine-1490 and threonine-1572 in colorectal tumors. The fact that LRP6 phosphorylation was already deregulated in human adenomas strongly suggests that phosphorylation of this protein may be involved in early stages of colorectal carcinogenesis. Unfortunately, we did not detect any significant association between LRP6 phosphorylation and the presence of mutations in KRAS or BRAF in the 53 adenocarcinomas analyzed. However, a greater number of colorectal tumors needs to be analyzed in order to clearly determine whether there is a link between LRP6 phosphorylation and KRAS or BRAF mutations. Otherwise, the fact that increased phosphorylation of LRP6 was observed in tumors exhibiting wild-type KRAS and BRAF suggests that this phosphorylation may be induced by other kinases (for example, CK1 and GSK3, see below) or oncogenic pathways activated in these CRC specimens.
In presence of Wnt, LRP6 is phosphorylated within the PPPS/TP motifs by GSK3β and multiple CK1 members, providing docking sites to bind Axin1 and GSK3, thereby sequestering both proteins away from the β-catenin destruction complex.73, 74 However, in our cell models, blockade of MEK reduced LRP6 phosphorylation and decreased β-catenin nuclear localization and transcriptional activity without affecting β-catenin expression. One could speculate that phosphorylation of LRP6 by ERK triggers a signaling cascade controlling β-catenin nuclear localization and activity, independently of Axin or the degradation complex. Such signaling cascade operating in parallel with β-catenin stabilization has recently been documented in other biological systems.75 Indeed, activation of LRP6/Rac1/JNK2 signaling has been suggested to mediate nuclear accumulation of β-catenin independently of its stabilization during canonical Wnt signaling in stromal cells from the bone marrow.75 Intriguingly, in our cell models such as KRAS-transformed IECs, JNK activity seems to control especially LRP6 expression and not phosphorylation suggesting that LRP6 can be regulated at multiple levels. Hence, the question of how MEK-dependent phosphorylation of LRP6 promotes β-catenin transcriptional activity remains unresolved and will need to be addressed in the future.
The cellular context of RAS mutations in CRC is complex. KRAS mutations usually occur early in the course of human disease, not long after APC or β-catenin mutations.1 The contribution of RAS mutations to colon carcinogenesis is therefore linked to an altered Wnt signaling pathway. Consistent with the multihit hypothesis for colon tumor development, previous reports in mice indicate that the addition of Ras mutations to Apc loss causes an increase in adenoma size, number and invasiveness in addition to promoting the expansion of cells bearing putative stem cell markers within the tumors.5, 67, 76, 77, 78 Furthermore, Phelps et al.66 observed that homozygous loss of Apc alone in the zebrafish intestine was insufficient to cause β-catenin nuclear translocation; rather, the nuclear accumulation of β-catenin, and associated proliferation, required the additional activation of KRas or Raf1 oncogenes. Interestingly, Myd88-dependent ERK activation was shown to drive intestinal tumorigenesis in Apcmin/+ mice.79 However, aside from these observations, a cohort of adenomas from Familial Adenomatous Polyposis (FAP) patients was recently analyzed and nuclear β-catenin staining was observed in the vast majority of cases, whereas the presence of KRAS mutations, detected in only 10% of the adenomas, was independent of β-catenin subcellular localization.80 In addition, cells homozygous for a targeted Apc mutation, and without evidence of any additional mutations at other loci, do show constitutive Wnt activation based on reporter assay.76 Thus, although there is little doubt that oncogenic KRAS/MAPK signaling synergizes with APC mutations in tumorigenesis by fully enhancing nuclear β-catenin translocation and activity, it has still remained unclear whether sustained MAPK activation is essential for human adenoma progression (please refer to the comment of Fodde and Tomlison71, 72).
In the future, it will be quite relevant to verify whether inhibiting LRP6 function can interfere with β-catenin signaling and tumoral properties of human CRC cells exhibiting APC mutation with or without KRAS or BRAF mutations. This would be a very important finding due to the frequency of such mutations in human CRC and the difficulty to target β-catenin, KRAS and BRAF.81 In this respect, the potential utility of LRP6 blocking antibodies to inhibit Wnt signaling in tumorigenesis was recently demonstrated.82
In conclusion, our study demonstrates that the Wnt/β-catenin signaling pathway acts as a novel target of MEK/ERK signaling involved in human colorectal tumorigenesis. Whereas further studies are needed to pinpoint the molecular mechanisms by which MEK-dependent LRP6 phosphorylation induces β-catenin/TCF4 activity, the present study provides a novel fundamental insight into how oncogenic KRAS/MAPK signaling controls epithelial oncogenesis in the intestine.
Materials and methods
Materials
The antibodies against β-catenin, c-myc, phosphorylated c-jun (serine-63), anti-HA, Fra1 and total ERK1/2 were from Santa Cruz Biotechnologies (Santa Cruz, CA, USA). Antibodies recognizing phosphorylated ERK1/2, LRP6 (serine-1490) and β-catenin (serine-552) as well as total LRP6 and TCF4 were obtained from Cell Signalling (Danvers, MA, USA). Antibodies against E-cadherin and β-catenin were from BD Pharmingen (Mississauga, ON, USA). Antibodies recognizing β-actin, phosphorylated LRP6 (threonine-1572) and β-catenin (tyrosine-142) were purchased from Millipore (Billerica, MA, USA). Antibodies recognizing phosphorylated β-catenin (tyrosines-86 and -654) were purchased from Abcam (Toronto, ON, Canada). The MEK inhibitor U0126 was purchased from LC Laboratories (Woburn, MA, USA) and ICG-001 was purchased from AbMole BioSciences (Kowloon, Hong Kong, China). DKK-1 was purchased from R&D Systems (Minneapolis, MN, USA). LGK974 was purchased from Adooq Bioscience (Irvine, CA, USA). The specific ERK inhibitor SCH7729846 and MEK inhibitor PD184352 were provided by Pfizer Inc. (Groton, CT, USA). For immunofluorescence, goat anti-rabbit AlexaFluoro488 fluorescein isothiocyanate-labeled or goat anti-mouse AlexaFluoro568-labeled secondary antibodies were from Molecular Probes (Life Technologies, Grand Island, NY, USA). Horseradish peroxidase antibodies were purchased from Amersham Biosciences (Pittsburg, PA, USA), whereas alkaline phosphatase-conjugated antibodies were purchased from Promega (Madison, WI, USA).
Expression vectors
Expression vectors for hemagglutinin-tagged wild-type MEK1 (wtMEK) and caMEK, with a conversion of S218 and S222 RAF1-dependent regulatory phosphorylation sites to aspartic residues, were provided by Dr J. Pouysségur (Nice, France). The HA-wtMEK and HA-caMEK constructs were subcloned into the retroviral expression vector pLXIN (Clontech, Mountain View, CA, USA), as previously described.23, 24 The pRL-SV40 Renilla luciferase reporter vector was from Promega (Nepean, ON, Canada). Expression vectors encoding for human ΔNTCF4, wt LRP6, LRP6-5A and KRASG12V were all provided by Addgene (Cambridge, MA, USA). The TCF reporter constructs TOPFLASH and its negative control FOPFLASH as well as the c-myc promoter reporter (4 × TBE2) and its control (4 × TBE2-mutated) were also purchased from Addgene.
Cell culture
The rat intestinal epithelial crypt cells IEC-6 stably overexpressing wtMEK, caMEK, pBABE, KRASG12V or BRAFV600E:ER were all generated after retroviral infection and were previously characterized and cultured as described.23, 24, 83 The colon carcinoma cell lines HT29 and DLD-1 (ATCC) were, respectively, cultured in McCoy's and RPMI media containing 10% fetal calf serum (Wisent, St-Bruno, QC, Canada).
Western blot analysis and immunoprecipitations
Protein extractions and western blot analyses were performed as described.24 Immunoprecipitations were performed exactly as performed previously.84
Immunofluorescence
Immunofluorescence experiments were performed as previously described.24 Pictures were acquired on a Zeiss confocal Laser Scanning Microscope (Carl Zeiss Canada, Toronto, ON, Canada) with a × 100/1.46 oil objective and analyzed using the Carl Zeiss software (ZEN). Nuclei were stained with the DNA dye 4,6-diamidino-2-phenylindole and negative controls (no primary antibody) were included in all experiments.
Transient transfection and luciferase assays
TOPFLASH assays: cells were transfected using Lipofectamine 2000 (Life Technologies) with 0.3 μg of TOPFLASH reporter, whereas the FOPFLASH luciferase vector was used as a control for transfection efficiency. C-MYC-luciferase assays: cells were transfected with 0.3 μg of c-myc promoter reporter (4 × TBE2) with the control (4 × TBE2-mut) reporter used as a control for transfection efficiency. The pRL-SV40 Renilla luciferase reporter vector (4 ng) was also co-transfected. After specific transfection times, luciferase activity was measured according to the Promega protocol, as described.84
RNA extraction and quantitative RT–PCR (qRT–PCR) analyses
Total RNA was isolated using the Totally RNA extraction kit (Life Technologies). RT–PCR analysis was performed using AMV-RT (Roche Diagnostics, Laval, QC, Canada) according to the manufacturer’s instructions. c-Myc, Axin2 and Lef-1 qPCR analyses were performed using a LightCycler apparatus (Roche Diagnostics). Experiments were run and analyzed with the LightCycler software 4.0 according to the manufacturer’s recommendations using the QuantiTect SYBR Green PCR Kit (Qiagen, Toronto, ON, Canada). All samples were run in triplicate. Target expression was quantified relatively to Tbp, Gapdh or Pbgd expression. PCR analyses were performed as described previously.24 Primers are available upon request.
Cell proliferation, soft agarose and migration/invasion assays
Experiments were performed starting with cell populations after 8 days post selection and subsequently plated for growth assay in six-well plates (100 000 cells per well). The number of cells was calculated every 2 days for a period of 7 days using a Cell particle counter. Soft agarose and migration/invasion assays were performed as reported.24, 83, 85
LRP6 silencing
To silence the expression of Lrp6 in caMEK cells, we used small hairpin RNA (shRNA) plasmids containing 29-mer shRNA sequence in green fluorescent protein (GFP) vector targeted against the rat Lrp6 gene expression. Noneffective 29-mer-scrambled shRNA cassette in pGFP-V-RS Vector (Origene, Rockville, MD, USA) was used as a control. GFP was used to monitor the efficiency of transfection. Lentiviruses produced in 293T cells were used for infection according to Invitrogen recommendations (ViralPower Lentiviral Expression System). Cells were infected with either control or Lrp6-shRNA lentiviruses. After selection with puromycin (1 μg/ml), cells were used for further studies.
Experiments in mice
1-Female nude CD1 nu/nu mice were purchased from Charles River (Wilmington, MA, USA). Tumor growth: 1 × 106 of control and experiemental cells suspended in DMEM were contralaterally injected into the subcutaneous tissue of mice.24 Tumor volume was determined by external measurement according to the formula d2 × D/2. Experimental tail vein assays: The tail vein of 5-week-old mice was injected with 1 × 106 cells suspended in 100 μl DMEM. Animals were killed at any sign of respiratory distress or weight loss, or after 28 days post injection.24 Lungs were maintained in Bouin's fixative for 2 days. 2-BRafCAmice carrying a Cre-activated allele of the murine BRaf gene were kindly provided by Dr Martin McMahon (Cancer Research Institute, UCSF/Diller Family Comprehensive Cancer Center).50 Mutations were genotyped according to published protocols.50 The C57BL/6 12.4 KbVilCre transgenic line (#004586) was provided by Jackson Laboratory (Bar Harbor, MA, USA). Mutations were genotyped as described previously.86 All experimental protocols were approved by the Ethics Committee for Animal Experimentation of the Université de Sherbrooke.
Human tumors
Samples of colorectal tumors and paired normal tissues (at least 10 cm from the tumor) were obtained from patients undergoing surgical resection. Patients did not receive neoadjuvant therapy. Tissues were obtained after patient’s written informed consent, according to the protocol approved by the Institutional Human Subject Review Board of the Centre Hospitalier Universitaire de Sherbrooke. All tissues were frozen in liquid nitrogen within 15 min from resection as recommended by the Canadian Tumor Repository Network (www.ctrnet.ca). Paired tissues were lysed and immunoblotted as previously described.85 Genomic DNA was extracted from formalin-fixed paraffin-embedded tissue using a FFPE DNA Isolation Kit for Cells and Tissues (Qiagen; Electron Microscopy & Histology platform at the Université de Sherbrooke). APC (exon 15), KRAS (exons 1 and 2) and BRAF (exon15) were amplified with PCR, and the presence of mutations was detected by direct sequencing (Plateforme de Séquençage et de Génotypage des Génomes du CRCHUL, QC, Canada).
Data presentation
Assays were performed in either duplicate or triplicate. Typical results shown are representative of three independent experiments. Densitometric analyses were performed by using the Image J software. Results were analyzed by the Student’s t-test and were considered statistically significant at P<0.05 or lower.
References
Fearon ER, Vogelstein B . A genetic model for colorectal tumorigenesis. Cell 1990; 61: 759–767.
Gregorieff A, Clevers H . Wnt signalling in the intestinal epithelium: from endoderm to cancer. Genes Dev 2005; 19: 877–890.
Taketo MM . Mouse models of gastrointestinal tumors. Cancer Sci 2006; 97: 355–361.
Ashton GH, Morton JP, Myant K, Phesse TJ, Ridgway RA, Marsh V et al. Focal adhesion kinase is required for intestinal regeneration and tumorigenesis downstream of Wnt/c-Myc signalling. Dev Cell 2010; 19: 259–269.
Sansom OJ, Meniel VS, Muncan V, Phesse TJ, Wilkins JA, Reed KR et al. Myc deletion rescues Apc deficiency in the small intestine. Nature 2007; 446: 676–679.
Worthley DL, Leggett BA . Colorectal cancer: molecular features and clinical opportunities. Clin Biochem Rev 2010; 31: 31–38.
Pretlow TP, Pretlow TG . Mutant KRAS in aberrant crypt foci (ACF): initiation of colorectal cancer? Biochim Biophys Acta 2005; 1756: 83–96.
Smakman N, Borel Rinkes IH, Voest EE, Kranenburg O . Control of colorectal metastasis formation by K-Ras. Biochim Biophys Acta 2005; 1756: 103–114.
Saucier C, Rivard N . Epithelial cell signalling in colorectal cancer metastasis. In: Beauchemin N, Huot J (eds) Metastasis of Colorectal Cancer vol.14. Springer: Netherlands, 2010, pp 205–241.
Dasari A, Messersmith WA . New strategies in colorectal cancer: biomarkers of response to epidermal growth factor receptor monoclonal antibodies and potential therapeutic targets in phosphoinositide 3-kinase and mitogen-activated protein kinase pathways. Clin Cancer Res 2010; 16: 3811–3818.
Dienstmann R, Tabernero J . BRAF as a target for cancer therapy. Anticancer Agents Med Chem 2011; 11: 285–295.
Ikenoue T, Kanai F, Hikiba Y, Obata T, Tanaka Y, Imamura J et al. Functional analysis of PIK3CA gene mutations in human colorectal cancer. Cancer Res 2005; 65: 4562–4567.
Deng G, Bell I, Crawley S, Gum J, Terdiman JP, Allen BA et al. BRAF mutation is frequently present in sporadic colorectal cancer with methylated hMLH1, but not in hereditary nonpolyposis colorectal cancer. Clin Cancer Res 2004; 10: 191–195.
Li WQ, Kawakami K, Ruszkiewicz A, Bennett G, Moore J, Iacopetta B . BRAF mutations are associated with distinctive clinical, pathological and molecular features of colorectal cancer independently of microsatellite instability status. Mol Cancer 2006; 5: 2.
Yuen ST, Davies H, Chan TL, Ho JW, Bignell GR, Cox C et al. Similarity of the phenotypic patterns associated with BRAF and KRAS mutations in colorectal neoplasia. Cancer Res 2002; 62: 6451–6455.
Rajagopalan H, Bardelli A, Lengauer C, Kinzler KW, Vogelstein B, Velculescu VE . Tumorigenesis: RAF/RAS oncogenes and mismatch-repair status. Nature 2002; 418: 934.
Ramos JW . The regulation of extracellular signal-regulated kinase (ERK) in mammalian cells. Int J Biochem Cell Biol 2008; 40: 2707–2719.
Aliaga JC, Deschenes C, Beaulieu JF, Calvo EL, Rivard N . Requirement of the MAP kinase cascade for cell cycle progression and differentiation of human intestinal cells. Am J Physiol 1999; 277: G631–G641.
Rivard N, Boucher MJ, Asselin C, L'Allemain G . MAP kinase cascade is required for p27 downregulation and S phase entry in fibroblasts and epithelial cells. Am J Physiol 1999; 277: C652–C664.
Fang JY, Richardson BC . The MAPK signalling pathways and colorectal cancer. Lancet Oncol 2005; 6: 322–327.
Eggstein S, Franke M, Kutschka I, Manthey G, von Specht BU, Ruf G et al. Expression and activity of mitogen activated protein kinases in human colorectal carcinoma. Gut 1999; 44: 834–838.
Lee SH, Lee JW, Soung YH, Kim SY, Nam SW, Park WS et al. Colorectal tumors frequently express phosphorylated mitogen-activated protein kinase. APMIS 2004; 112: 233–238.
Boucher MJ, Jean D, Vezina A, Rivard N . Dual role of MEK/ERK signalling in senescence and transformation of intestinal epithelial cells. Am J Physiol Gastrointest Liver Physiol 2004; 286: G736–G746.
Lemieux E, Bergeron S, Durand V, Asselin C, Saucier C, Rivard N . Constitutively active MEK1 is sufficient to induce epithelial-to-mesenchymal transition in intestinal epithelial cells and to promote tumor invasion and metastasis. Int J Cancer 2009; 125: 1575–1586.
Komatsu K, Buchanan FG, Katkuri S, Morrow JD, Inoue H, Otaka M et al. Oncogenic potential of MEK1 in rat intestinal epithelial cells is mediated via cyclooxygenase-2. Gastroenterology 2005; 129: 577–590.
Voisin L, Julien C, Duhamel S, Gopalbhai K, Claveau I, Saba-El-Leil MK et al. Activation of MEK1 or MEK2 isoform is sufficient to fully transform intestinal epithelial cells and induce the formation of metastatic tumors. BMC Cancer 2008; 8: 337–2407-8-337.
Sebolt-Leopold JS, Dudley DT, Herrera R, Van Becelaere K, Wiland A, Gowan RC et al. Blockade of the MAP kinase pathway suppresses growth of colon tumors in vivo. Nat Med 1999; 5: 810–816.
Ikenoue T, Hikiba Y, Kanai F, Aragaki J, Tanaka Y, Imamura J et al. Different effects of point mutations within the B-Raf glycine-rich loop in colorectal tumors on mitogen-activated protein/extracellular signal-regulated kinase kinase/extracellular signal-regulated kinase and nuclear factor kappaB pathway and cellular transformation. Cancer Res 2004; 64: 3428–3435.
Cagnol S, Rivard N . Oncogenic KRAS and BRAF activation of the MEK/ERK signalling pathway promotes expression of dual-specificity phosphatase 4 (DUSP4/MKP2) resulting in nuclear ERK1/2 inhibition. Oncogene 2013; 32: 564–576.
Nandan MO, McConnell BB, Ghaleb AM, Bialkowska AB, Sheng H, Shao J et al. Kruppel-like factor 5 mediates cellular transformation during oncogenic KRAS-induced intestinal tumorigenesis. Gastroenterology 2008; 134: 120–130.
Brondello JM, Brunet A, Pouyssegur J, McKenzie FR . The dual specificity mitogen-activated protein kinase phosphatase-1 and -2 are induced by the p42/p44MAPK cascade. J Biol Chem 1997; 272: 1368–1376.
Brunet A, Pages G, Pouyssegur J . Growth factor-stimulated MAP kinase induces rapid retro-phosphorylation and inhibition of MAP kinase kinase (MEK1). FEBS Lett 1994; 346: 299–303.
Gopalbhai K, Meloche S . Repression of mitogen-activated protein kinases ERK1/ERK2 activity by a protein tyrosine phosphatase in rat fibroblasts transformed by upstream oncoproteins. J Cell Physiol 1998; 174: 35–47.
Tian X, Liu Z, Niu B, Zhang J, Tan TK, Lee SR et al. E-cadherin/beta-catenin complex and the epithelial barrier. J Biomed Biotechnol 2011; 2011: 567305.
Yochum GS, Cleland R, Goodman RH . A genome-wide screen for beta-catenin binding sites identifies a downstream enhancer element that controls c-Myc gene expression. Mol Cell Biol 2008; 28: 7368–7379.
Clevers H . Wnt/beta-catenin signalling in development and disease. Cell 2006; 127: 469–480.
Naishiro Y, Yamada T, Takaoka AS, Hayashi R, Hasegawa F, Imai K et al. Restoration of epithelial cell polarity in a colorectal cancer cell line by suppression of beta-catenin/T-cell factor 4-mediated gene transactivation. Cancer Res 2001; 61: 2751–2758.
van de Wetering M, Sancho E, Verweij C, de Lau W, Oving I, Hurlstone A et al. The beta-catenin/TCF-4 complex imposes a crypt progenitor phenotype on colorectal cancer cells. Cell 2002; 111: 241–250.
Emami KH, Nguyen C, Ma H, Kim DH, Jeong KW, Eguchi M et al. A small molecule inhibitor of beta-catenin/CREB-binding protein transcription. Proc Natl Acad Sci USA 2004; 101: 12682–12687.
Rowan AJ, Lamlum H, Ilyas M, Wheeler J, Straub J, Papadopoulou A et al. APC mutations in sporadic colorectal tumors: A mutational "hotspot" and interdependence of the "two hits". Proc Natl Acad Sci USA 2000; 97: 3352–3357.
Yang J, Zhang W, Evans PM, Chen X, He X, Liu C . Adenomatous polyposis coli (APC) differentially regulates beta-catenin phosphorylation and ubiquitination in colon cancer cells. J Biol Chem 2006; 281: 17751–17757.
Hart M, Concordet JP, Lassot I, Albert I, del los Santos R, Durand H et al. The F-box protein beta-TrCP associates with phosphorylated beta-catenin and regulates its activity in the cell. Curr Biol 1999; 9: 207–210.
Piedra J, Miravet S, Castano J, Palmer HG, Heisterkamp N, Garcia de Herreros A et al. p120 Catenin-associated Fer and Fyn tyrosine kinases regulate beta-catenin Tyr-142 phosphorylation and beta-catenin-alpha-catenin Interaction. Mol Cell Biol 2003; 23: 2287–2297.
Brembeck FH, Schwarz-Romond T, Bakkers J, Wilhelm S, Hammerschmidt M, Birchmeier W . Essential role of BCL9-2 in the switch between beta-catenin's adhesive and transcriptional functions. Genes Dev 2004; 18: 2225–2230.
Roura S, Miravet S, Piedra J, Garcia de Herreros A, Dunach M . Regulation of E-cadherin/Catenin association by tyrosine phosphorylation. J Biol Chem 1999; 274: 36734–36740.
Taurin S, Sandbo N, Qin Y, Browning D, Dulin NO . Phosphorylation of beta-catenin by cyclic AMP-dependent protein kinase. J Biol Chem 2006; 281: 9971–9976.
Fang D, Hawke D, Zheng Y, Xia Y, Meisenhelder J, Nika H et al. Phosphorylation of beta-catenin by AKT promotes beta-catenin transcriptional activity. J Biol Chem 2007; 282: 11221–11229.
He XC, Yin T, Grindley JC, Tian Q, Sato T, Tao WA et al. PTEN-deficient intestinal stem cells initiate intestinal polyposis. Nat Genet 2007; 39: 189–198.
Krejci P, Aklian A, Kaucka M, Sevcikova E, Prochazkova J, Masek JK et al. Receptor tyrosine kinases activate canonical WNT/beta-catenin signalling via MAP kinase/LRP6 pathway and direct beta-catenin phosphorylation. PLoS ONE 2012; 7: e35826.
Dankort D, Curley DP, Cartlidge RA, Nelson B, Karnezis AN, Damsky WE J et al. Braf(V600E) cooperates with Pten loss to induce metastatic melanoma. Nat Genet 2009; 41: 544–552.
Morris EJ, Jha S, Restaino CR, Dayananth P, Zhu H, Cooper A et al. Discovery of a novel ERK inhibitor with activity in models of acquired resistance to BRAF and MEK inhibitors. Cancer Discov 2013; 3: 742–750.
Casalino L, De Cesare D, Verde P . Accumulation of Fra-1 in ras-transformed cells depends on both transcriptional autoregulation and MEK-dependent posttranslational stabilization. Mol Cell Biol 2003; 23: 4401–4415.
Liu J, Pan S, Hsieh MH, Ng N, Sun F, Wang T et al. Targeting Wnt-driven cancer through the inhibition of Porcupine by LGK974. Proc Natl Acad Sci USA 2013; 110: 20224–20229.
Proffitt KD, Virshup DM . Precise regulation of porcupine activity is required for physiological Wnt signalling. J Biol Chem 2012; 287: 34167–34178.
Fernandez-Medarde A, Santos E . Ras in cancer and developmental diseases. Genes Cancer 2011; 2: 344–358.
Malumbres M, Barbacid M . RAS oncogenes: the first 30 years. Nat Rev Cancer 2003; 3: 459–465.
Niault TS, Baccarini M . Targets of Raf in tumorigenesis. Carcinogenesis 2010; 31: 1165–1174.
Daugherty RL, Gottardi CJ . Phospho-regulation of Beta-catenin adhesion and signalling functions. Physiology (Bethesda) 2007; 22: 303–309.
Bienz M . beta-Catenin: a pivot between cell adhesion and Wnt signalling. Curr Biol 2005; 15: R64–R67.
Gumbiner BM . Regulation of cadherin adhesive activity. J Cell Biol 2000; 148: 399–404.
Harris TJ, Peifer M . Decisions, decisions: beta-catenin chooses between adhesion and transcription. Trends Cell Biol 2005; 15: 234–237.
Nelson WJ, Nusse R . Convergence of Wnt, beta-catenin, and cadherin pathways. Science 2004; 303: 1483–1487.
Perez-Moreno M, Fuchs E . Catenins: keeping cells from getting their signals crossed. Dev Cell 2006; 11: 601–612.
Singh A, Sweeney MF, Yu M, Burger A, Greninger P, Benes C et al. TAK1 inhibition promotes apoptosis in KRAS-dependent colon cancers. Cell 2012; 148: 639–650.
Horst D, Chen J, Morikawa T, Ogino S, Kirchner T, Shivdasani RA . Differential WNT activity in colorectal cancer confers limited tumorigenic potential and is regulated by MAPK signalling. Cancer Res 2012; 72: 1547–1556.
Phelps RA, Chidester S, Dehghanizadeh S, Phelps J, Sandoval IT, Rai K et al. A two-step model for colon adenoma initiation and progression caused by APC loss. Cell 2009; 137: 623–634.
Wang C, Zhao R, Huang P, Yang F, Quan Z, Xu N et al. APC loss-induced intestinal tumorigenesis in Drosophila: roles of Ras in Wnt signalling activation and tumor progression. Dev Biol 2013; 378: 122–140.
Bennecke M, Kriegl L, Bajbouj M, Retzlaff K, Robine S, Jung A et al. Ink4a/Arf and oncogene-induced senescence prevent tumor progression during alternative colorectal tumorigenesis. Cancer Cell 2010; 18: 135–146.
Rad R, Cadinanos J, Rad L, Varela I, Strong A, Kriegl L et al. A genetic progression model of Braf(V600E)-induced intestinal tumorigenesis reveals targets for therapeutic intervention. Cancer Cell 2013; 24: 15–29.
Carragher LAS, Snell KR, Giblett SM, Aldridge VSS, Patel B, Cook SJ et al. V600EBraf induces gastrointestinal crypt senescence and promotes tumour progression through enhanced CpG methylation of p16INK4a. EMBO Mol Med 2010; 2: 458–471.
Zeller E, Hammer K, Kirschnick M, Braeuning A . Mechanisms of RAS/beta-catenin interactions. Arch Toxicol 2013; 87: 611–632.
Fodde R, Tomlinson I . Nuclear beta-catenin expression and Wnt signalling: in defence of the dogma. J Pathol 2010; 221: 239–241.
Tamai K, Zeng X, Liu C, Zhang X, Harada Y, Chang Z et al. A mechanism for Wnt coreceptor activation. Mol Cell 2004; 13: 149–156.
Wolf J, Palmby TR, Gavard J, Williams BO, Gutkind JS . Multiple PPPS/TP motifs act in a combinatorial fashion to transduce Wnt signalling through LRP6. FEBS Lett 2008; 582: 255–261.
Wu X, Tu X, Joeng KS, Hilton MJ, Williams DA, Long F . Rac1 activation controls nuclear localization of beta-catenin during canonical Wnt signalling. Cell 2008; 133: 340–353.
Janssen KP, Alberici P, Fsihi H, Gaspar C, Breukel C, Franken P et al. APC and oncogenic KRAS are synergistic in enhancing Wnt signalling in intestinal tumor formation and progression. Gastroenterology 2006; 131: 1096–1109.
Luo F, Brooks DG, Ye H, Hamoudi R, Poulogiannis G, Patek CE et al. Mutated K-ras(Asp12) promotes tumourigenesis in Apc(Min) mice more in the large than the small intestines, with synergistic effects between K-ras and Wnt pathways. Int J Exp Pathol 2009; 90: 558–574.
Haigis KM, Kendall KR, Wang Y, Cheung A, Haigis MC, Glickman JN et al. Differential effects of oncogenic K-Ras and N-Ras on proliferation, differentiation and tumor progression in the colon. Nat Genet 2008; 40: 600–608.
Lee SH, Hu LL, Gonzalez-Navajas J, Seo GS, Shen C, Brick J et al. ERK activation drives intestinal tumorigenesis in Apc(min/+) mice. Nat Med 2010; 16: 665–670.
Obrador-Hevia A, Chin SF, Gonzalez S, Rees J, Vilardell F, Greenson JK et al. Oncogenic KRAS is not necessary for Wnt signalling activation in APC-associated FAP adenomas. J Pathol 2010; 221: 57–67.
Kahn M . Can we safely target the WNT pathway? Nat Rev Drug Discov 2014; 13: 513–532.
Ettenberg SA, Charlat O, Daley MP, Liu S, Vincent KJ, Stuart DD et al. Inhibition of tumorigenesis driven by different Wnt proteins requires blockade of distinct ligand-binding regions by LRP6 antibodies. Proc Natl Acad Sci USA 2010; 107: 15473–15478.
Bergeron S, Lemieux E, Durand V, Cagnol S, Carrier JC, Lussier JG et al. The serine protease inhibitor serpinE2 is a novel target of ERK signalling involved in human colorectal tumorigenesis. Mol Cancer 2010; 9: 271–4598-9-271.
Simoneau M, Coulombe G, Vandal G, Vezina A, Rivard N . SHP-1 inhibits beta-catenin function by inducing its degradation and interfering with its association with TATA-binding protein. Cell Signal 2011; 23: 269–279.
Langlois MJ, Bergeron S, Bernatchez G, Boudreau F, Saucier C, Perreault N et al. The PTEN phosphatase controls intestinal epithelial cell polarity and barrier function: role in colorectal cancer progression. PLoS ONE 2010; 5: e15742.
Madison BB, Dunbar L, Qiao XT, Braunstein K, Braunstein E, Gumucio DL . Cis elements of the villin gene control expression in restricted domains of the vertical (crypt) and horizontal (duodenum, cecum) axes of the intestine. J Biol Chem 2002; 277: 33275–33283.
Acknowledgements
We thank Guillaume Arguin and Gérald Bernatchez for technical assistance. The biobank of colorectal cancer specimens was supported by a Team grant on digestive epithelium from the Canadian Institutes of Health Research. This research was supported by a grant from the Canadian Institutes of Health Research Grant to Nathalie Rivard (MT-14405). Étienne Lemieux is a student scholar from the Fonds de la Recherche en Santé du Québec (FRSQ). Nathalie Rivard and Julie C Carrier are members of the FRSQ-Funded Centre de Recherche du CHUS. Nathalie Rivard is a recipient of a Canadian Research Chair in colorectal cancer and inflammatory cell signalling. This research was supported by a grant from the Canadian Institutes of Health Research to NR (MT-14405).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on the Oncogene website
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Lemieux, E., Cagnol, S., Beaudry, K. et al. Oncogenic KRAS signalling promotes the Wnt/β-catenin pathway through LRP6 in colorectal cancer. Oncogene 34, 4914–4927 (2015). https://doi.org/10.1038/onc.2014.416
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/onc.2014.416
This article is cited by
-
Wnt signaling in colorectal cancer: pathogenic role and therapeutic target
Molecular Cancer (2022)
-
Triple-negative breast cancer: understanding Wnt signaling in drug resistance
Cancer Cell International (2021)
-
Random survival forests identify pathways with polymorphisms predictive of survival in KRAS mutant and KRAS wild-type metastatic colorectal cancer patients
Scientific Reports (2021)
-
Gain and loss of function mutations in biological chemical reaction networks: a mathematical model with application to colorectal cancer cells
Journal of Mathematical Biology (2021)
-
Landscape of transcriptome variations uncovering known and novel driver events in colorectal carcinoma
Scientific Reports (2020)
