

Abstract
Background/Aim: During pregnancy, maternal liver can be affected by ethanol (ETOH) intake, whose effects depend on concentration levels ingested. This study aims to describe histological and serum marker characteristics of maternal liver during two metabolic conditions: gestation (G), and sustained ETOH intake, in early and late pregnancy. Materials and Methods: Wistar rats were fed with Lieber-DeCarli diet during pregnancy, following an experimental protocol that allows a semi-chronic intake of ETOH (5%). Liver and serum samples were processed for histological characterization and biochemical profiling. Hematoxylin/eosin and Schiff’s Periodic Acid staining were used. Results: During pregnancy, a significant elevation in ballooned and edamatous hepatocytes, and a significant increase in micro and macrovesicular deposits were observed in rats fed with the ETOH diet at gestation days 3G, 8G and 15G. These changes were reverted by 20G. Liver glycogen content increased significantly at 15G. Serum metabolites in pregnant rats fed with the ETOH diet showed a significant reduction in urea (from 3G to 15G), an increase in albumin and uric acid at 20G, and a reduction in creatinine. Number of offsprings and weight of male newborns were reduced by 20% and 14%, respectively. Liver function markers in serum showed no significant changes. Conclusion: ETOH diet intake promotes hepatic histological changes and histological modifications during pregnancy. These results support the assumption that pregnancy is an adaptive procedure that is associated with nutritional conditions and has a strong influence on hepatic histology. They suggest that pregnancy promotes a state of resilience to the liver function during the sustained intake of 5% ETOH.
- Lieber-DeCarli diet
- gestation
- fat vesicles
- ballooned hepatocytes
- ethanol
- maternal resilience
Pregnancy is a physiological state that involves a sequence of adaptations that allow fetal and placental development. During this state, the assimilation and distribution of nutrients is rheostatically regulated to favor the offspring’s growth, and at the same time, the process promotes the correct function of the maternal organism. In this context, a variety of maternal behavioral, metabolic, and physiological adaptations have been categorized by some authors as a new rheostat state (1, 2). For example, within the maternal liver, a discreet fat accumulation in hepatocytes has been reported during different stages of pregnancy without pathological repercussions (3). Many reports have shown the detrimental effects that nutritional and pharmacological insults during pregnancy can cause to the fetus and offspring, but there is little research regarding the maternal consequences of alcohol consumption (4, 5). Ethanol (ETOH) is a natural product that works as a metabolic intermediate; however, it is also a recreational drug and the main component of alcoholic drinks. Even though the pathological consequences of ETOH intake have been extensively reported (6, 7), especially as a hepatotoxic factor, less is known regarding the effects it can have on maternal liver function during pregnancy (8). Experimentally, ETOH can be administered under various protocols; in recent years, one of the most-used models to promote alcoholic liver injury in rodents has been the ad libitum feeding with the Lieber-DeCarli liquid diet containing ETOH 5% for some weeks. This protocol was characterized by moderating the blood alcohol concentrations, and it induced a mild liver injury and a low-grade liver inflammation without fibrosis (9). In this context, it has been reported that 20-65% of women ingest ETOH at some point during pregnancy (10). In early pregnancy, an increase in fat deposits occurs, promoted by maternal hyperphagia and increased insulin sensitivity, which stimulates fatty acid (FA) esterification in adipocytes (1, 2). In late pregnancy, there is an increased lipolytic activity from adipose tissue, promoting elevated concentrations of free FA (5, 6). It has been reported that before and during pregnancy the consumption of ETOH, even moderately related to the development of hepatic injury in the offspring (10-12); nonetheless, little is known about the histological and functional response of maternal hepatic function.
This study aimed to investigate the histological characterization of the liver of pregnant rats fed during early and late pregnancy with the liquid diet of Lieber–DeCarli that involves a sustained 5% of ETOH consumption. The results showed that this experimental diet promoted differential histological features in the maternal liver in early and late pregnancy. Interestingly, some of these alterations were also observed in pregnant rats fed with the control diet without ETOH.
Materials and Methods
Ethical approval. The procedures were carried out in accordance with the Official Mexican Standard: NOM-062-ZOO-1999. This research was reviewed and authorized by the Ethics Committee of the Neurobiology Institute, at the Universidad Nacional Autonoma de Mexico (UNAM) (approval ID 081. A).
Experimental groups. The rats were individually held in cages (21×23.5×38 cm) under controlled 12 h dark/light conditions with lights on at 07:00 am. A progressive study was performed in mothers during gestation (G) days (3, 8, 15, and 20). Early pregnancy was considered from gestational day 3 to 8, and late pregnancy from gestational day 15 to 20.
Sixty female Wistar rats weighing 250-300 g, were randomly divided into 4 groups:
1) Non-pregnant rats fed with control diet (CNP): n=5 (for a period of 7 days). 2) Non-pregnant rats fed with experimental diet containing 5% ETOH (ENP): n=5 (for a period of 7 days). 3) Pregnant rats fed with control diet (CP): n=24, 6 rats in each day. Early pregnancy (3G and 8G); late pregnancy (15G and 20G). 4) Pregnant rats fed with experimental diet containing 5% ETOH (EP): n=24, 6 rats in each day. Early pregnancy (3G and 8G); late pregnancy (15G and 20G).
Characterization of the number and sex of rat offspring. Once the pregnancy had elapsed, and the control and experimental groups had delivered the offspring of each mother, the separation of males and females was carried out, and the offspring was weighed on a scale to obtain their body weight (A&D Weighing series GF-3000, A&D Korea Limited, Seoul, Republic of Korea).
Diet and body weight. A control liquid diet (F1259SP, Lieber-DeCarli 82, Bio-Serv, Flemington, NJ, USA) (54.3% carbohydrate, 17.7% lipid, 16.7% protein at 355 kcal/l) and an experimental liquid diet containing 5% EtOH (F1258SP, Lieber-DeCarli 82, Bio Serv) (25.5% carbohydrate, 29.9% fat, 28.2% protein at 355 kcal/l) were used. Even though both diets are isocaloric, important nutritional differences are evident: the experimental ETOH diet contains 53% less carbohydrates, and 69% more lipids and proteins (13). All groups had access to diets ad libitum as well as water intake. Before pregnancy and during pregnancy, food and water intake were quantified, and body weight was registered using a precision scale (A&D Weighing series GF-3000 scale).
Live tissue and serum sampling. On days 3G, 8G, 15G, and 20G, rats were rapidly decapitated with a guillotine to obtain blood samples; tissues were dissected, weighed, and cut into slices for biochemical and histological processing. Blood was collected in Eppendorf tubes and centrifuged at 400×g for 10 min to obtain the serum, which was stored in aliquots at −20°C until analysis. Dissected liver tissues were obtained and processed for histological analysis.
Histological techniques. Once the livers were weighed and finely dissected, samples were fixed with 4% formaldehyde and stained with hematoxylin-eosin. Shortly thereafter, the livers were cut with dissection equipment, scalpels, and scissors, embedded in paraffin, and 3 μm slices were made with a rotation microtome (Leica RM2135, Leica Microsystems, Nussloch, Germany). A trained pathologist determined the hepatic morphological changes; histological characteristics, such as the number of fat micro- and macrovesicles, as well as ballooned hepatocytes, necrosis, and inflammation (14), using a Nikon eclipse e200 microscope (Tokyo, Japan).
To study PAS-positive staining, liver samples were immersed in Mayer’s hematoxylin solution for 15 min after deparaffinization and hydration. Deparaffinized sections were immersed in 1% periodic acid for 10 min, washed with distilled water for 2 min, immersed in Schiff reagent (Product 191203S, BDH Laboratory Supplies, Poole, UK) for 10 min, and then washed under running tap water for 10 min. Finally, they were dehydrated, cleared, and cover-slipped for quantitative analysis.
Biochemical determination in serum. Alanine aminotransferase (ALT/TGP) and aspartate aminotransferase (AST/TGO) assays were carried out on a Spinreact Spin 120 automated equipment for colorimetric assays.
In order to measure the insulin levels, enzyme-linked immunosorbent assay (ELISA) kits for insulin serum [American Laboratory Products Company (ALPCO) for the North American life science markets (Cat. 80-INSHU-E01.1)] were used.
To determine serum urea and creatinine concentrations, colorimetric methods were used with commercial kits (Spinreact, Santa Coloma, Spain). The values of glucose, the lipid profile, cholesterol levels, and triacylglycerides (TG) were also determined using (Spinreact) commercial kits for colorimetric assays.
Statistical analysis. Statistical analyses were performed with the SPSS Statistics software (IBM, Armonk, NY, USA), considering a value of p<0.05 as statistically significant. The Kolmogorov-Smirnov and the Shapiro Wilk tests were used to test the normality of the experimental variables. To compare non-pregnant rats fed with the control and ETOH diets, Student’s t-test mediated by Levene’s test was used. One-way ANOVA was used to compare the temporal variations of the pregnant rats fed with the control and ETOH diets. Two-way ANOVA was used to compare the control and experimental groups during early and late pregnancy.
Results
Diet consumption, water intake and body weight. Five days before pregnancy, the control and ETOH-fed groups showed an enhanced consumption of the liquid diet (Figure 1A). When the rats were not pregnant, experimental and control groups tended to reduce their intake, showing fluctuations until 12G. During pregnancy, ETOH-fed rats showed lower values at 3G and 5G. After 12G, a reduction in the diet consumption in the experimental group was evident, especially the days before the offspring’s birth (20G). Water intake (Figure 1B) was very similar in both groups of rats, before and during pregnancy, with the exception of the last 3 days of pregnancy (17G to 20G), when the ETOH-fed rats drank significantly less water (Figure 1B). Non-pregnant rats fed with the control diet reached more than 300 g of body weight but, when they got pregnant, their weight decreased more than 30 g. Nevertheless, during pregnancy, this group constantly gained weight until it reached more than 400 g at 20G. In contrast, rats fed the ETOH diet showed a reduction in weight when they became pregnant, with no significant changes in liver function markers. Their weight gains during pregnancy were more discreet, since they reached less than 350 g at 20G.

Food, water intake, and body weight of non-pregnant and pregnant rats. Non-pregnant (rectangle on left side, where the first 7 days are indicated) control group, black circles; experimental group with ethanol (ETOH), white circles. Days shown after the rectangle correspond to pregnancy from day 1 to 21 of gestation (G) (black circles: control group and white circles: ETOH group). (A) Food intake (kcal), (B) Water intake, (C) Body weight. Data are shown as mean±standard error, and *p<0.05.
Data in Figure 1C indicate that nutritional differences between the control and ETOH diets could be related to the lower weight gain shown by the experimental group, especially during the late pregnancy period.
Organ morphometry. The liver/body weight ratio showed no change, neither during early nor late pregnancy, nor when the control and ETOH diet groups were compared (Figure 2).
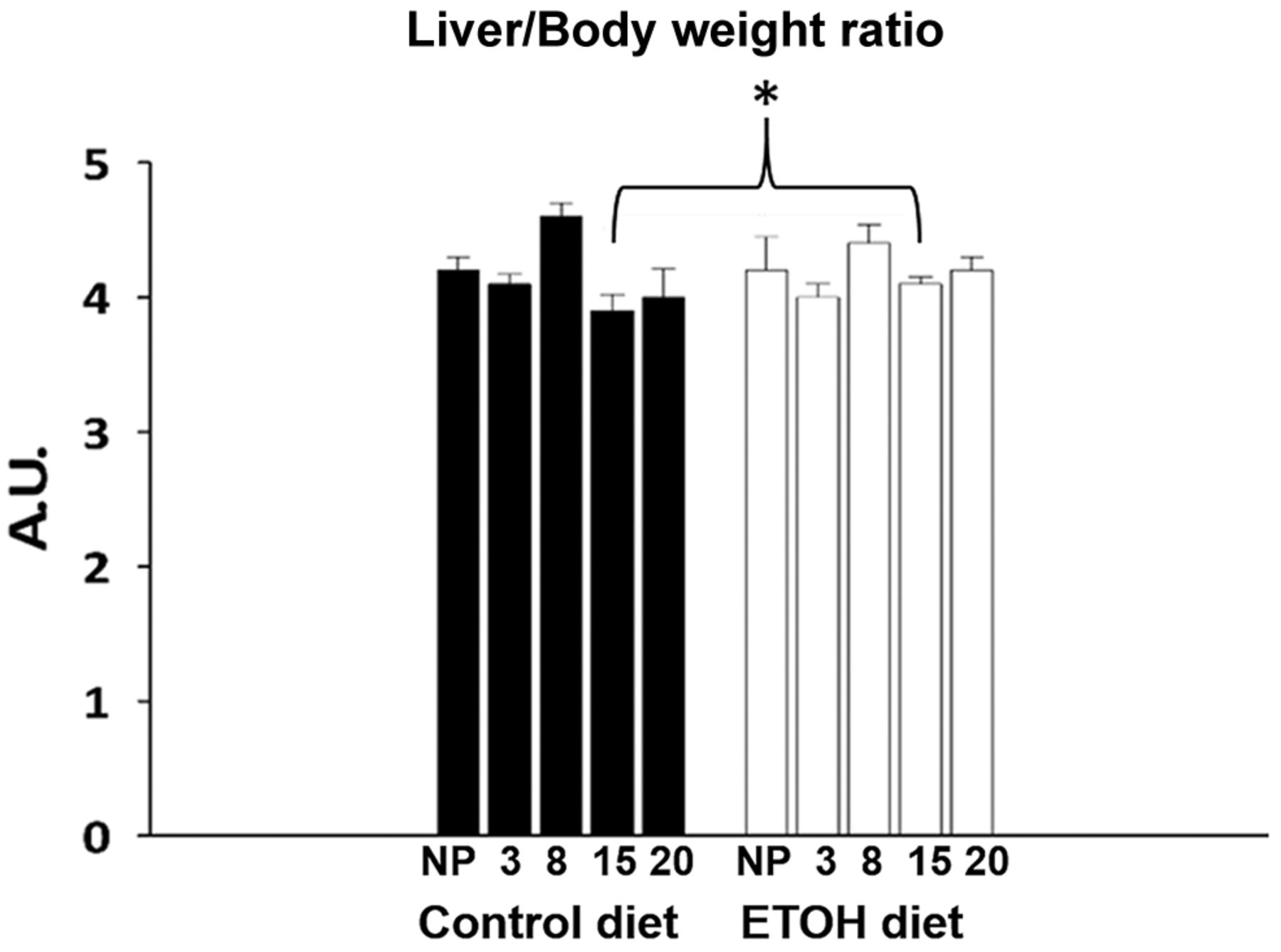
Morphometric characteristics of the liver at different stages of pregnancy. White bars correspond to the ethanol (ETOH) diet group; black bars correspond to the control group. Different temporal points during pregnancy are shown for the liver to body weight ratio for pregnant rats: early pregnancy 3G (day 3 of gestation), 8G, late pregnancy 15G, 20G. Liver to body weight ratio is also shown for non-pregnant (NP) rats. Data shown are mean±standard error. *p<0.05.
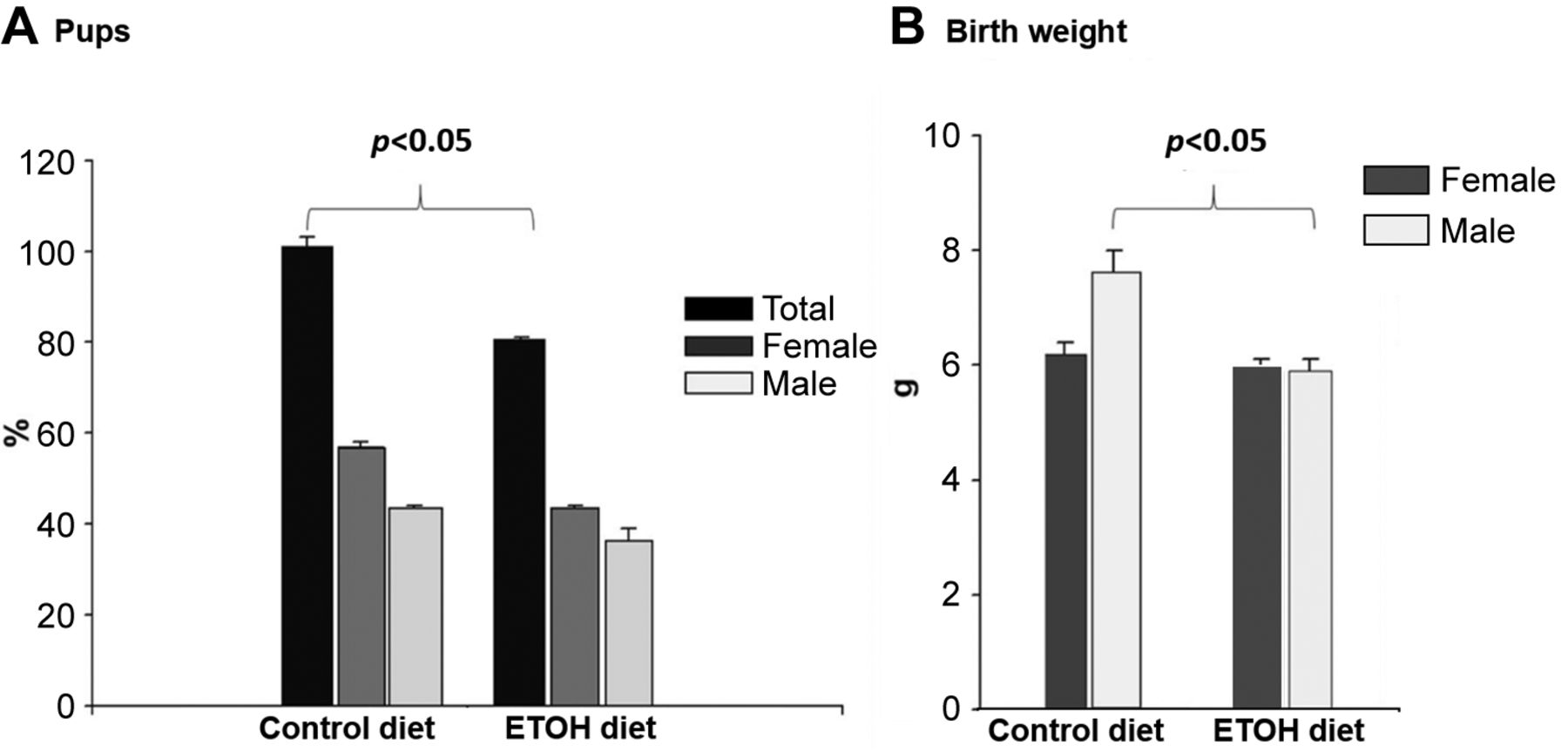
Offspring characteristics. The protocol studied in this project involved two coincident metabolic challenges: pregnancy, and the consumption of an experimental liquid diet containing 5% of ETOH. In this context, it was important to characterize the pregnancy success rate in both control and experimental conditions. Figure 3A shows that the rats fed with ETOH had 20% less offspring, with no important changes in their sex ratio. On its part, Figure 3B shows that the male progeny of rats fed with the control diet had significantly higher weight.

Number of offspring and birth weight of mothers who consumed a diet with ethanol (ETOH) or a control diet. (A) Percentage of offspring at birth. Black bars correspond to the total percentage (males and females), dark gray bars to the percentage of females and light gray to the percentage of males. (B) Birth weight of offspring. Dark gray bars correspond to females and light gray bars to males. Data are shown as mean±standard error. *p<0.05.
In contrast, ETOH fed rats had male and female progenies with similar weight, analogous to the weight shown by the female progeny of the control group.
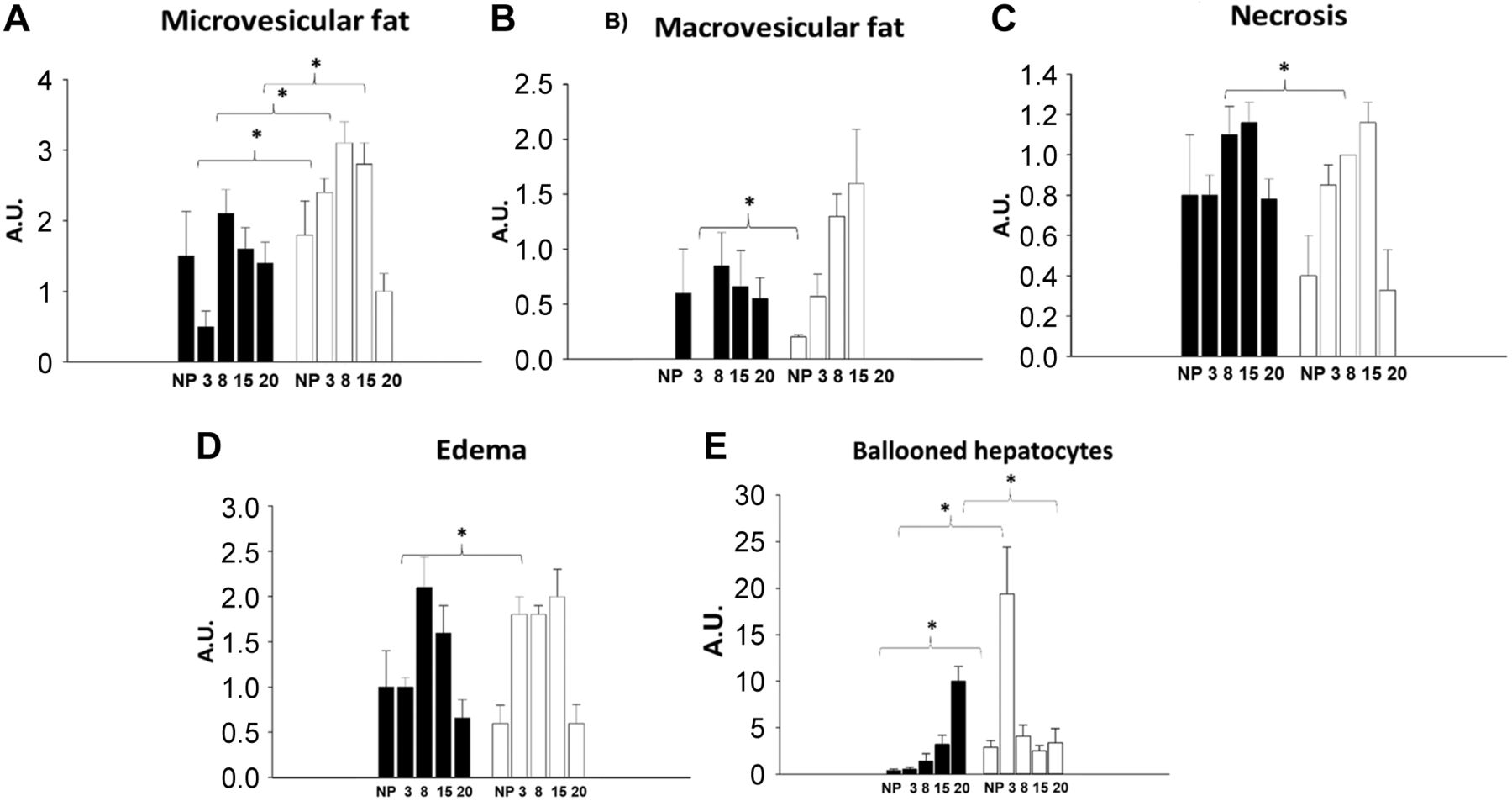
Liver histology. Figure 4 shows a histological liver slice from a non-pregnant rat fed with the control diet. No accumulation of microvesicular cytoplasmic, enlarged hyperchromatic nuclei, moderate cellular edema, or granulated cytoplasm was detected. No damage was observed either. One major finding, identified through the liquid Lieber–DeCarli diet, was that several histological changes were observed even in the control diet. As shown in Figure 4, non-pregnant rats fed with Chow diet (Figure 4A), the control diet (Figure 4B), and the ETOH diet (Figure 4C) showed discreet but detectable lipid vesicles, hepatocytes with edema, and hyperchromatic nuclei, in comparison to the liver image from an intact rat fed with a Chow diet. In addition, the non-pregnant rats fed with the ETOH diet showed some ballooned hepatocytes (Figure 4C). During pregnancy, the following changes were identified in the group fed with the control and ETOH diet (Figure 5). The histological results of the liver have important implications: 1) the Lieber DeCarli diet promotes hepatic alterations, not only because of the ETOH presence, but also given the nutritional impact related to the high content of carbohydrates (Figure 5A, B, C, and D; 2) the group fed with control diet showed a reduction in lipid vesicles in early pregnancy, while it showed a high proportion of edematous and ballooned hepatocytes during late pregnancy (Figure 5A and E); 3) In contrast, the group fed with the ETOH diet showed a very high presence of ballooned hepatocytes at early pregnancy, while an important increase in lipid micro and macrovesicles was the main change observed during late pregnancy (Figure 5).

Liver biopsies in non-pregnant rats. (A) Liver biopsy of non-pregnant rats fed with Chow control diet. Accumulation of fat microvesicles was not observed in the cytoplasm. Enlarged hyperchromatic nuclei, cellular edema, granulated cytoplasm, no damage in central vein (CV) and sinusoids (SI), normal hepatocytes (H), and trabeculae (T) were observed. (B) Liver biopsy of non-pregnant rat with control diet intake. No damage, little accumulation of micro vesicular cytoplasmic (MV), lipid droplets (LP), and moderate cellular edema (E) are observed. (C) Liver biopsy of non-pregnant rat fed with ethanol (ETOH) diet, shows diffuse cell damage with significant cell edema (E), hyperchromatic nuclei (HN), microvesicular cytoplasmic (MC), lipid droplets (LP), and some hepatocytes have clear cytoplasm (BH).
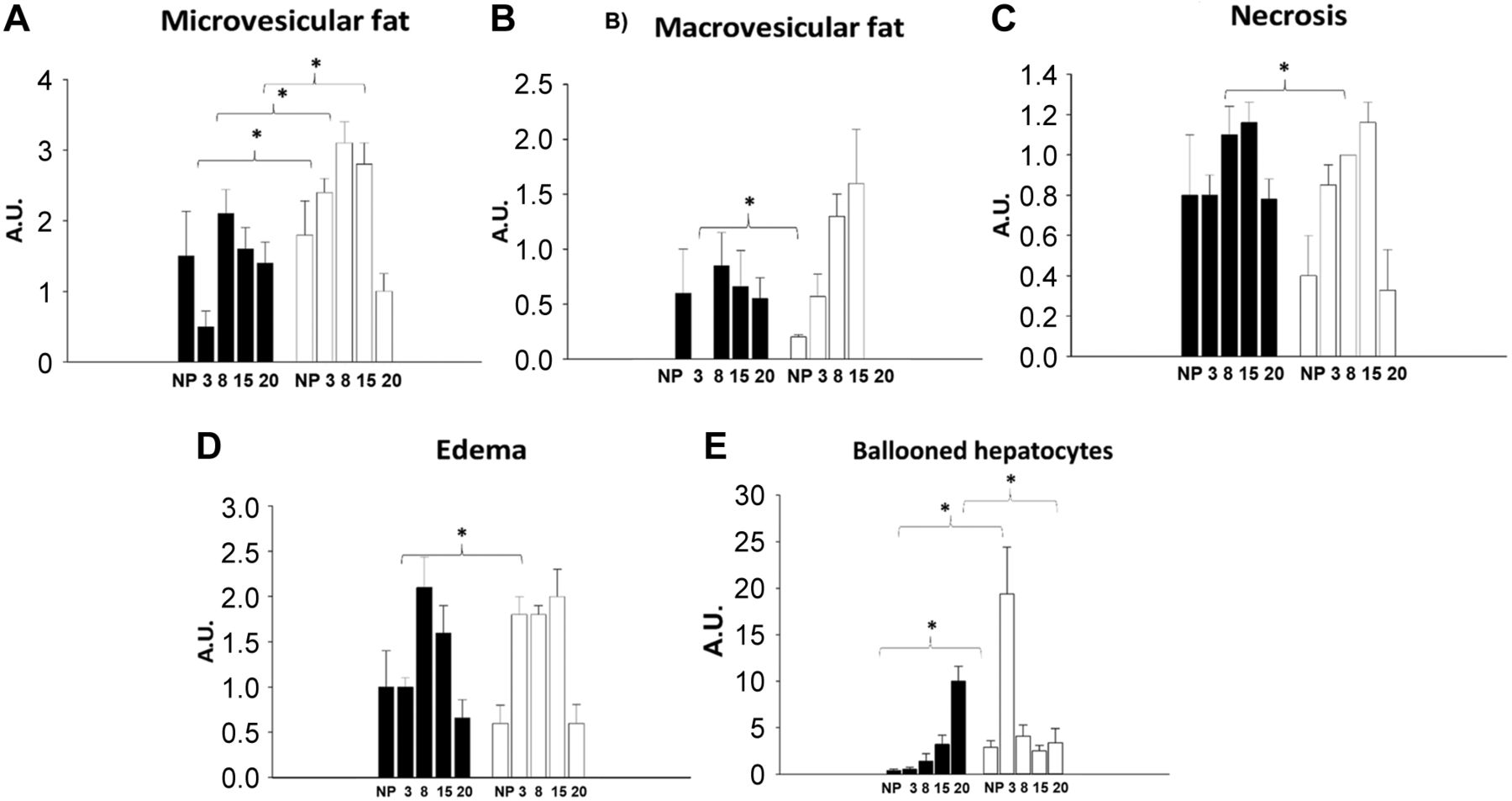
Liver histology by hematoxylin and eosin (H&E) in pregnant rat. Pathological evaluation in the mothers’ liver during pregnancy with control and ETOH diet. Shows proportion in arbitrary units (A.U.) microvesicles of fat item (A), fat macrovesicles of fat item (B), necrosis item (C), edema item (D) and balloon hepatocytes item (E).
Figure 6A shows that lipid vesicles decreased in the control diet-fed group at 3G, while Figure 6B shows that hepatocytes with edema increased in the ETOH diet-fed group, as well as a notorious presence of ballooned hepatocytes was shown for both at 3G. Figure 7B shows that the ETOH diet-fed group displayed a different pattern of hepatic histological changes at 15G, along with a significant elevation in lipid micro and macrovesicles. In turn, a reduction was observed in lipid micro and macrovesicles in the control diet-fed group at 15G (Figure 7A), as well as a return to normal liver histological architecture at 20G (Figure 5A).

Liver histology by hematoxylin and eosin (H&E) in an early pregnant rat. (A) Liver biopsy of pregnant rat on the control diet group at 3G, showed no damage, very little edema (E), hyperchromatic nuclei (HN), while hepatocytes are seen with clear cytoplasm (ballooned hepatocytes) (BH) and micro vesicular lipid droplets (LP). (B) Liver biopsy of pregnant rat in the ethanol (ETOH) diet group at 3G, showed diffuse cell damage with significant cell edema (E), hyperchromatic nuclei (HN), while hepatocytes are seen with clear cytoplasm (ballooned hepatocytes) (BH) and micro vesicular lipid droplets (LP).
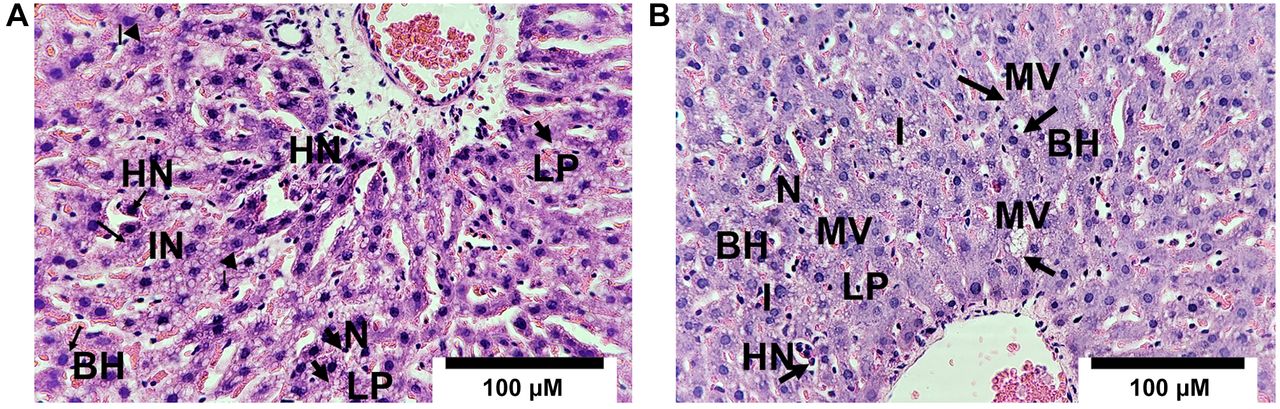
Liver histology by hematoxylin and eosin (H&E) in the biopsy of a late pregnant rat. (A) Liver biopsy of rats pregnant with control diet group at 15G shows prominent cellular damage, with micro vesicular lipid droplets (LP), moderate inflammation (IN), hyperchromatic nuclei (HN), with significant architectural derangement, balloon hepatocytes (BH), necrotic-appearing hepatocytes (N), (B) Liver biopsy of rat pregnant in the control ETOH group at 15G shows cellular damage, with micro vesicular (MV), and macro vesicular lipid droplets, macro vesicular (LP) inflammation (I), hyperchromatic nuclei (HN), with significant architectural derangement, balloon hepatocytes (BH), and necrotic-appearing hepatocytes (N).
PAS staining. The content of hepatic glycogen was similar in non-pregnant rats in both groups, with the control and ETOH diets. During pregnancy, there was a clear tendency to increase the glycogen in the livers of both groups, especially during 8G and 15G. Nevertheless, the pregnant group fed with the ETOH diet showed a very significant elevation in the PAS staining at 15G (temporal profile is shown in Figure 8). All the rats were in the postprandial period.

Liver biopsy of periodic acid Schiff (PAS) histology of non-pregnant (NP) and pregnant rats fed with (white bars) and without ethanol (ETOH) diet (black bars) [(3G and 8G days of gestation (early pregnancy), 15G and 20G of gestation (late pregnancy)]. (A) The glycogenic pattern was stained with PAS. The presence of aldehyde groups formed by previous oxidation of carbohydrates is demonstrated. The glycogen concentration was significantly higher at day 15 of gestation with the ETOH diet. (B) PAS histological images. (H) Hepatocyte, (G) Glycogen. Data are shown as mean±standard error; and *p<0.05. A.U.: Arbitrary unit.
Serum hepatic markers. The temporal profiles of all the serum parameters studied in this project are included in Figure 9. Similarities and differences can be observed in non-pregnant and pregnant rats fed with the control and ETOH diets.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Biomarkers in serum of non-pregnant (NP) and pregnant rats fed with (white bars) and without ethanol (ETOH) diet (black bars) [(3G and 8G of gestation (early pregnancy), 15G and 20G of gestation (late pregnancy)]. Different temporal points during pregnancy are shown: NP (not pregnant) and pregnancy 3 (on day 3 of gestation), 8 (on day 8 of gestation), 15 (on day 15 of gestation), 20 (on day 20 of gestation); Alanine (A), Urea (B), Uric acid (C), Alanine aminotransferase (ALT) (D), Creatinine (E), Glucose (F), Triacylglycerides (G), Cholesterol (H), Insulin (I). Data are shown as mean±standard error; and *p<0.05.
In order to obtain a general metabolic and endocrine profile in early and late pregnancies as well as to characterize the effect of the ETOH diet, a number of serum metabolites and hormones were quantified (see Methods section). Not all the parameters measured showed changes, due to either the pregnancy process or consumption of the ETOH diet. The metabolites and hormones that showed significant changes are presented in Figure 9. Alanine amino transferase (ALT) is one of the major markers of hepatic dysfunction. In our protocol, there was a discreet elevation in the group fed with the ETOH diet at 15G and 20G (late pregnancy), that was significant at 15G. A similar pattern was observed for uric acid and albumin, since a significant increase was seen at 20G in the group fed with the ETOH diet with both metabolites. Rats fed with the ETOH diet showed a significant decrease in two metabolites, creatinine at 20G, and cholesterol at 8G.
Urea in the group fed with the control diet showed a discreet reduction at 20G; in contrast, the rats fed with the ETOH diet showed a generalized decrease in circulating urea during pregnancy and even in the previous non-pregnant state. A similar pattern was observed with insulin, since the control group showed a modest reduction during pregnancy and just a more important decrease at 20G. Again, the group fed with the ETOH diet showed less circulating insulin, starting before pregnancy, and also remaining throughout pregnancy.
Rats fed with the control diet showed a marked hypoglycemia and hypertriacylglyceridemia in late pregnancy (15G and 20G). The same pattern occurred in the group fed with the ETOH diet, only with one exception: at 20G, this group showed a much higher increase in circulating triacylglycerides. Taken together, the data of serum metabolites and hormones suggest metabolic and endocrine adaptations during pregnancy, some of which being modified by the ETOH diet.
Discussion
ETOH is a recreational substance with a well-established social acceptance. In some circumstances, its consumption can lead to serious health problems with important repercussions on the liver function (15). Several experimental models have been implemented to characterize the biological consequences of ETOH consumption. These models involve acute, semi-chronic, and chronic treatments, as well as their application in a variety of doses and physiological conditions (16-20).
In experimental models (male rats), it has been shown that ETOH consumption causes liver damage, which is reflected in the presence of histological alterations, such as fat accumulation and cellular disorganization (21). Furthermore, it has been shown that acute ETOH consumption may represent pathological changes in sinusoidal endothelial cells prior to biochemical or histological liver damage (22). Liver damage caused by ETOH intake has been shown to be associated with the development of hepatic steatosis (>5-10% accumulation of fat in liver cells) and is generally accompanied by dyslipidemia, primarily by the increased levels of triacylglycerides (23, 24). We have not identified any studies that focus and show liver histological and serum biochemical changes during pregnancy due to ETOH consumption. To our knowledge, this is the first report on the histological changes in the liver due to sustained consumption of ETOH during early and late pregnancy, and one that shows that an acute ETOH dietary intake promotes different hepatic histological changes during pregnancy. No correspondence between histological alterations and serum metabolite profile was detect.
Some results suggest that prenatal exposure to ETOH affects fetal body weight, but not liver weight (25), and that prenatal ETOH intake may lead to liver dysfunction in the offspring (26). In one study, consumption (5%) of ETOH for 2 weeks before pregnancy was shown to decrease the number of viable fetuses and lead to abnormal fetal development, and these effects were accompanied by hepatic steatosis during pregnancy. The ETOH metabolic pathways in the liver are well identified and recognized, but considerably less is known on the enzymes and pathways involved in ethanol metabolism during pregnancy progression (27). In the present project, we used the Lieber-DeCarli liquid diet, which is a suitable animal model associated with a sustained intake of 5% ETOH as part of an isocaloric diet (13, 15). This liquid diet was applied throughout the pregnancy to explore possible physiopathological repercussions on the maternal organism.
A noteworthy aspect in our study was that only the Lieber–DeCarli liquid diet control formula was able to promote some histological changes in liver tissue before and during pregnancy. In this regard, it was reported that ETOH did not affect maternal or neonatal plasma insulin levels. Thus, maternal ethanol consumption reduces maternal and neonatal liver glycogen storage and leads to postnatal hypoglycemia in the newborn rat (28).
The mouse model induced by the Lieber–DeCarli ETOH liquid diet plus single ETOH binge feeding induces liver injury, inflammation, and fatty liver, which mimics acute-chronic alcoholic liver injury in patients (29). Exposure to ETOH in lactation has also been associated with long-term deficits in cellular immunity (30). During the progression of steatosis, nutrients and their metabolites can also induce the secretion of adipokines and inflammatory factors by adipocytes, macrophages, and other cells, triggering inflammation (31). A nutritional consideration, in addition to the ETOH content, is that the control diet contains a high proportion of carbohydrates (more than double) and a considerable reduction in fat and protein content (nearly 40% in both). The control group gained weight throughout the experimental protocol, except at the beginning of pregnancy, and had abundant visceral adipose tissue at the end of the process (data not shown). In contrast, the group fed with the ETOH diet gained significantly less weight in late pregnancy. Further studies are needed to describe in detail the mechanisms responsible for the hepatic histological alterations in the two groups of non-pregnant rats. So far, the only difference was that the group fed with ETOH showed ballooned hepatocytes, whereas the control group did not.
A set of adaptations was associated with the pregnancy process in rats fed with the control diet. In the liver, the main finding was a reduction in lipid vesicles on 3G (early pregnancy), whereas on 15G and 20G there was an increase in edematous and ballooned hepatocytes. In addition, PAS staining was discreetly elevated on 8G. These histological alterations did not affect the state of pregnancy; since at birthday a normal number of offspring was observed (12 offspring on average).
Several reported modifications were detected in serum, mainly during late pregnancy, such as reduction in glucose, uric acid, creatinine, and insulin, as well as an increase in circulating triacylglycerides. In experimental models (male rats), it has been shown that ETOH consumption leads to liver damage, which is reflected in the presence of biochemical and histological alterations, including serum ALT and bilirubin levels (22).
The constant presence of ETOH in the diet during pregnancy promoted a 20% reduction in the number of offspring. ETOH also disturbed the hepatic histological adaptations as it promoted the appearance of a large proportion of ballooned hepatocytes on 3G and favored a very significant elevation in fat micro and macrovesicles on 15G. In addition, a very high peak of glycogen content on 15G was observed. In serum, ETOH promoted a significant elevation in albumin, uric acid, ALT, and triacylglycerides on 20G. In contrast, ETOH induced a generalized reduction in urea and insulin throughout the pregnancy.
Limitations of the study were the following: pregnant rats were not fasted for ethical reasons; this experimental design cannot be carried out in primates also for ethical reasons; and the diet intervention is prescribed for moderate damage.
Conclusion
In conclusion, these results support the notion that pregnancy is an adaptive procedure associated with the nutritional condition with strong influence on hepatic histology. This study also indicated that ETOH consumption during pregnancy is capable of modifying histological, metabolic, and endocrine parameters. In our protocol, these modifications did not show a strong impact on pregnancy since the Lieber–DeCarli diet promoted the birth of only 20% fewer offspring. ETOH consumption during pregnancy induced histological changes in the liver that did not compromise the functioning of this metabolic organ. In early pregnancy, diffuse changes were observed with an increase in the accumulation of fat microvesicles, ballooned hepatocytes, and inflammation, on 8G and 15G; they presented a significant increase in micro- and macrovesicular deposits. In late pregnancy (20G), the group reverted all the mentioned hepatic histological changes. Hence, we demonstrated that the combination of two metabolic challenges, namely pregnancy and ETOH intake, did not induce changes in hepatic function even though a very evident hepatic histological modification was detected.
Acknowledgements
The Authors would like to thank the following persons and institutions for their assistance and support of this study: José Martín García Servín, MVZ, for the handling and care of the experimental model; the Animal Care Facility (Bioterio) of the Neurobiology Institute, UNAM; Microscopy Unit of the Institute of Neurobiology (UNAM), PhD Paola Bello Medina for her support in the statistics analysis and Jonathan Rafael Trinidad Gallardo of the Southern University Center, University of Guadalajara for his support in a part integrating histology figures and graphs.
Footnotes
Authors’ Contributions
MDM and MNM participated in the research design. MNM, EARA, JACR, MCB, DLM MAAL conducted the experiments. MDM, MNM and FST performed the data analysis. MNM and MDM wrote the manuscript.
Conflicts of Interest
The Authors have no conflicts of interest in relation to this study.
- Received June 1, 2022.
- Revision received July 8, 2022.
- Accepted July 27, 2022.
- Copyright © 2022, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY-NC-ND) 4.0 international license (https://creativecommons.org/licenses/by-nc-nd/4.0).