

Abstract
Background/Aim: Macrophages and biomaterial-induced multinucleated giant cells (BMGCs) are central elements in the tissue reaction cascade towards bone substitute materials (BSM). The enzymatic detection of the lytic enzyme tartrate-resistant acid phosphatase (TRAP) has manifoldly been used to examine the so-called “bioactivity” of BSM. The present study aimed to compare the detection validity and expression pattern of the TRAP enzyme using enzymatic and immunohistochemical detection methods in the context of biocompatibility analyses of BSM. Patients and Methods: Biopsies from 8 patients were analyzed after sinus augmentation with a xenogeneic bone substitute. Analysis of both macrophage and BMGC polarization were performed by histochemical TRAP detection and immunohistochemical detection of TRAP5a. Histomorphometrical analysis was used for comparison of the TRAP detection of BMGCs. Results: The enzymatic TRAP detection method revealed that in 7 out of 8 biopsies only single cells were TRAP-positive, whereas most of the cells and especially the BMGCs were TRAP-negative. The immunohistochemical detection of TRAP5a showed moderate numbers of stained mononuclear cells, while the majority of the BMGCs showed signs of TRAP5a-expression. The enzymatic TRAP detection was comparable to the results obtained via immunohistochemistry only in one case. The histomorphometrical analysis showed that significantly more mononuclear and multinucleated TRAP-positive cells were found using immunohistochemical TRAP5a-staining compared to the enzymatic TRAP detection method. Also, significantly more TRAP-negative BMGCs were found using the enzymatic TRAP detection. Conclusion: The immunohistochemical detection of TRAP is more accurate for examination of the bioactivity and cellular degradability of BSM.
- Tartrate-resistant acid phosphatase
- TRAP
- immunohistochemistry
- biomaterial
- biocompatibility
- inflammation
- macrophages
- multinucleated giant cells
- osteoclasts
- cellular biology
- biomaterial-associated giant cells
The degradability of a bone substitute material (BSM) is an important factor of its clinical applicability as it should optimally act as a an osteoconductive scaffold being resorbed even simultaneously to the bony regeneration process to meet the requirements of the concept of “creeping substitution” (1, 2). Two different degradation pathways are known for calcium phosphate (CaP)-based BSMs and their proportionate extent of the degradation behavior depends on various material factors (3, 4): (a) solution-mediated extracellular dissolution and (b) cell-based resorption. The dissolution behavior of a BSM can be measured via standardized in vitro tests that have manifoldly been described (5, 6). Also, the cellular degradability has manifoldly been analyzed via in vitro models using monocytic precursor cells of different origin that were stimulated to osteoclasts (6, 7). However, the in vivo analysis of this process – analyzed via both preclinical and clinical studies – is more accurate due to the problem of inadequate in vitro simulation of multicellular events.
In this context, the cellular reactivity to biomaterials, such as BSM, has extensively been analyzed in the last decades (8). It was initially assumed in the field of (dental) biomedical research that the multinucleated giant cells especially induced by calcium phosphate-based materials are also osteoclasts and their induction shows the osteoinductive potential of such biomaterials even due to their chemical comparability with native bone matrix (9-13). Thereby, a manifoldly applied enzymatic staining method to locate osteoclasts via enzymatic histochemical detection of tartrate-resistant acid phosphatase (TRAP) molecules was adapted for the evaluation of biomaterial degradation (13). The TRAP molecule is a metalloprotein enzyme involved in signal transduction, activation, proliferation, and differentiation (14, 15). TRAP is expressed by different catabolic cells including osteoclasts and alveolar macrophages (16). In osteoclasts, TRAP is expressed within the area of the ruffled border and intracellular compartments such as the (phago-) lysosomes or the Golgi cisternae during trans- or phagocytosis (15). This enzyme has been shown to be involved in the process of bone matrix degradation (15). Thus, the detection of TRAP within the different above-mentioned compartments of osteoclast shows that matrix degradation occurs not only extracellularly within the resorption lacunae but also intracellularly within the different vesicles (15). The over-expression of TRAP has been correlated with different diseases, which has made it an important marker in diagnostic pathology (17, 18). These include leukemic reticuloendotheliosis (hairy cell leukemia), Gaucher’s disease, HIV-induced encephalopathy, osteoclastoma and osteoporosis, and metabolic bone diseases (17-20). Mainly, TRAP has been established as a marker to track the status of osteoporosis (14, 20, 21). Therefore, this detection method was used as no other procedure was available for many decades. Thereby, TRAP staining has manifoldly been used to examine the so-called “bioactivity” of BSMs (22-25). Moreover, TRAP expression, as a marker of osteoclastic activity, and its increased expression in implantation beds of BSMs has been correlated with osteo-inductivity based on the well-known bilateral cross-connection between osteoblasts and osteoclasts (9). However, more knowledge about the tissue reactivity to BSM and even of the TRAP enzyme family has been gained in the last years.
Moreover, it has been shown that most biomaterials, even both natural and synthetic BSM, do not induce a physiological but inflammatory tissue reaction, the so-called “foreign body reaction to biomaterials” (25-27). It has also been found that the multinucleated cells induced by BSM are inflammatory and can be related to the foreign body giant cell (FBGC) type rather than being osteoclasts, as physiological cells involved in the bone turnover process (26, 27). Additionally, it was shown that that this cell type expresses both pro- and anti-inflammatory cytokines like M1 and M2 macrophages (28, 29). Macrophages play essential roles in the host tissue reaction to biomaterials via the induction of pro-inflammatory and/or anti-inflammatory cascades (28). The pro-inflammatory cascade is induced by the ‘classically activated’ M1 macrophages and is characterized by the production of cytotoxic cytokines and lytic enzymes. The anti-inflammatory cascade is induced by the ‘alternatively activated’ M2 phenotype and is characterized by the production of healing and anti-inflammatory cytokines. Moreover, it is assumed that the polarization of biomaterial-induced macrophages between M1 and M2 phenotypes must be balanced to allow for proper tissue integration, vascularization, biodegradation, and tissue regeneration (28, 30). Due to their central position, it is nowadays assumed that both related phenotypes are key players in the material-mediated healing cascade through expression of a broad variety of molecules such as heme oxygenase-1 (HO-1) and mannose receptor (MR) (25). Based on the similarity of the multinucleated giant cells to macrophages, in this material-related cascade, this special cell type has been designated as “biomaterial-induced multinucleated giant cells” (BMGCs) to distinguish them from “pure” FBGCs (25, 31, 32). Their involvement in the phagocytosis of biomaterials, such as BSM, makes both cell types very important. Both macrophages and BMGCs are involved in the pro-inflammatory arm of the tissue reaction cascade, which is also associated with the biodegradation of a biomaterial (32). In this context, it has shown that both macrophages but especially BMGCs express TRAP as a marker of their lytic activity (17, 33).
The research on this topic has furthermore revealed that TRAP is secreted in two different isoforms, i.e., TRAP 5b and TRAP 5a, which derive via post-translational modification (Table I) (14, 16, 17). The isoforms have a structural difference, TRAP 5a is the intact protein and TRAP 5b is the cleaved protein made of two subunits (21, 34). TRAP 5b is generally expressed by osteoclasts under physiological conditions like bone remodeling (16, 34). TRAP 5a was found to be expressed by cells of monocytic origin under pro-inflammatory conditions (21). TRAP 5a is inhibited by heparin, which possesses anti-inflammatory properties (21, 35).
The enzymatic histochemical staining of TRAP, that is routinely used in diagnostic pathology, detects TRAP proteins without isoform specificity (36). Moreover, the detection via the enzymatic staining procedure might be inaccurate. This staining procedure is carried out in three main steps: i) tissue sections are immersed in a solution containing the substrate of the enzyme, ii) induced-fit reactions occur between the respective enzyme and the substrate, and iii) the tissue sections are immersed in a solution with a marker to bind with the enzyme-substrate complex (37). In the enzymatic staining the conservancy of the enzyme activity must always be considered as it can be impacted, e.g., by a loss in the amount of enzyme or removal of enzyme activators. Therefore, tissue sections prepared for enzymatic staining are either minimally fixated with formalin, or frozen-fixated. Formalin is the most commonly used fixation medium in histology and pathology to crosslink macromolecules and consequently preserve tissue structures (38). Formaldehyde-macromolecule reactions are reversible through the washing of formalin-embedded samples. In contrast, the principle of immunohistochemical detection is to bind respective epitopes with a primary antibody (39). In case of immunohistochemical stainings, the primary antibody is then bound with a secondary antibody that is afterwards visualized by means of a marker molecule. The signal can be increased using a biotinylated secondary antibody combined with conjugated streptavidin or the peroxidase-anti-peroxidase (PAP) method. Thus, this staining method is generally assumed to increase the detection sensitivity and accuracy.
The present study was conducted to confirm the hypothesis of a pro-inflammatory origin of BMGCs and the validity of the histochemical TRAP detection. Thereby, a new antibody against TRAP5a was used and its validity compared to the routinely used enzymatic histochemical staining was examined. Histological and histomorphometrical analyses based on previously described methods were carried out on 8 human biopsies collected after sinus augmentation using a xenogeneic BSM (25, 39).
Materials and Methods
Clinical procedure. The sinus biopsies were obtained from 10 patients at the Department of Oral and Maxillofacial Surgery of the University Hospital Cologne. The tissue was obtained prior to insertion of dental implant after 6 months. Thereby, approval of the local ethics committees of the University of Cologne, Duesseldorf and Muenster, Germany, were initially obtained. Written informed consent was obtained from the patient(s) to publish this paper. Sinus floor elevation was conducted using the xenogeneic BSM Bio-Oss® (Geistlich Pharma AG, Wolhusen, Switzerland). In brief, initial antibiotic administration (Amoclav 500 mg (Hexal AG, Holzkirchen, Germany) or Clindamycin 600 mg (Clindasaar, MIP Pharma GmbH, Blieskastel, Germany) combined with oral disinfection via chlorhexidine solution (Chlorhexamed Fluid, 0.1% GlaxoSmithKline Consumer Healthcare GmbH, Bühl, Germany) for 30 s were performed. The sinus augmentation procedure was then started with mucoperiostal reflection and creation of a lateral bone window using diamond burs followed by local anesthesia (Ultracain D-S, Sanofi-Aventis GmbH, Frankfurt, Germany). Then, elevation of the Schneiderian membrane and implantation of a porcine pericardium-based collagen membrane (Alpha Bio’s Graft®, Alpha Bio, Petach Tikva, Israel) were performed. Afterwards, the xenogeneic BSM that was rehydrated with sterile saline solution was inserted into the sinus cavity. Finally, a second collagen membrane was inserted for covering of the BSM and the gingiva was closed via suturing (PGA Resorba, Resorba, Nuremberg, Germany). At six months, the biopsies were obtained via trephine burs (4-mm diameter, Ustomed, Tuttlingen, Germany) and tooth implants (SPI®, Alpha Bio) were implanted followed by final wound closure via suturing.
Histological preparation and staining methods. The following histological workup was conducted as previously published (39). Briefly, the biopsies were initially decalcified in 10% Tris-buffered ethylenediaminetetraacetic acid (Carl Roth, Karlsruhe, Germany) at 37°C for 10 days followed by dehydration in a series of increasing alcohol concentrations and final xylol exposure. Then, the biopsies were embedded in paraffin followed by sectioning by means of a rotation microtome (SLEE, Mainz, Germany) to a thickness of 3-5 μm. The respective first slide was used for enzymatic TRAP-detection, as previously described (39, 40). Briefly, a TRAP-buffer was prepared (pH value of 5) and was added to the prepared staining solution. The slides were incubated in the prepared solution overnight at 370C. Subsequently, a second staining using hematoxylin for 10 min followed by bluing for 5 min was performed. The second slide of each tissue explant was used for immunohistochemical detection of the TRAP5a molecule. In brief, immunohistochemistry was initiated with a pre-treatment with citrate buffer (pH value of 6) for 20 min in a water bath at 960C followed by an equilibration with TBS-T buffer (pH value of 9) as antibody-retrieving step. Afterwards, the slides were treated with H2O2 (from: UltraVision™ Quanto Detection System, ThermoFisher Scientific, Germany) followed by blocking using avidin and biotin blocking solutions (Avidin/Biotin Blocking Kit, Vector Laboratories, Newark, CA, USA). Then, the tissue slides were incubated with the TRAP5a antibody for 30 min followed by treatment with the secondary antibody (goat anti-rabbit IgG-B, 1:200, Santa Cruz Biotechnology, Dallas, TX, USA). After that, the avidin-biotin-peroxidase complex (ABC) (30 min), the DAB Quanto chromogen and substrate (10 min) or the Histostain-Plus IHC Kit including AEC (20 min) were applied (all: ThermoFisher Scientific) (Table I). A final staining via hematoxylin for 10 min combined with by bluing for 5 min was performed.
Both the histological and histomorphometrical analysis were conducted based on previously described protocols developed by Barbeck et al. (25, 26, 41-43). Initially, the histological analysis covered the observation of the staining results with special regard to comparison of the two TRAP detection methods. Microphotographs were obtained using a light microscope (Axio Scope. A1, Zeiss, Oberkochen, Germany) with a connected digital cam (Axiocam 105, Zeiss).
For the histomorphometrical analysis a scanning microscope combined of an Axio Scope. A1 microscope in combination with a scanning table and the ZEN Core software V3 (all: Zeiss) was used to digitize the slides. The Zen Core software was also used for cell counting, as previously published (5, 6, 9-13). Briefly, the cell numbers were counted via the “count tool” of the software and related to the total implant area (cells/mm2).
Statistical analysis. The statistical analysis was conducted by combination of an initial normality test (Shapiro–Wilk test), analysis of variance (ANOVA) and a subsequent LSD post-hoc test using the GraphPad Prism 9 software (GraphPad Software Inc., La Jolla, CA, USA). Statistically different values were stated as significant in case of p-values less than 0.05 (*p<0.05) or as highly significant in case of p-values less than 0.01 (**p<0.01) or less than 0.001 (***p<0.001). Afterwards, the graphs were created based on the means and standard deviations.
Results
Histopathological results. The histopathological analysis revealed that only TRAP-positive macrophages were detected using the enzymatic approach and in 8 out of 10 biopsies TRAP-negative BMGCs were found (Figure 1A and B). In one case, TRAP-positive BMGCs were observed on the surface of the xenogeneic BSM (Figure 1C). Using immunohistochemical staining to detect TRAP5a, macrophages and BMGCs stained positive in all biopsies. The BMGCs were found on the surface of the xenogeneic BSM (Figure 1B and C).
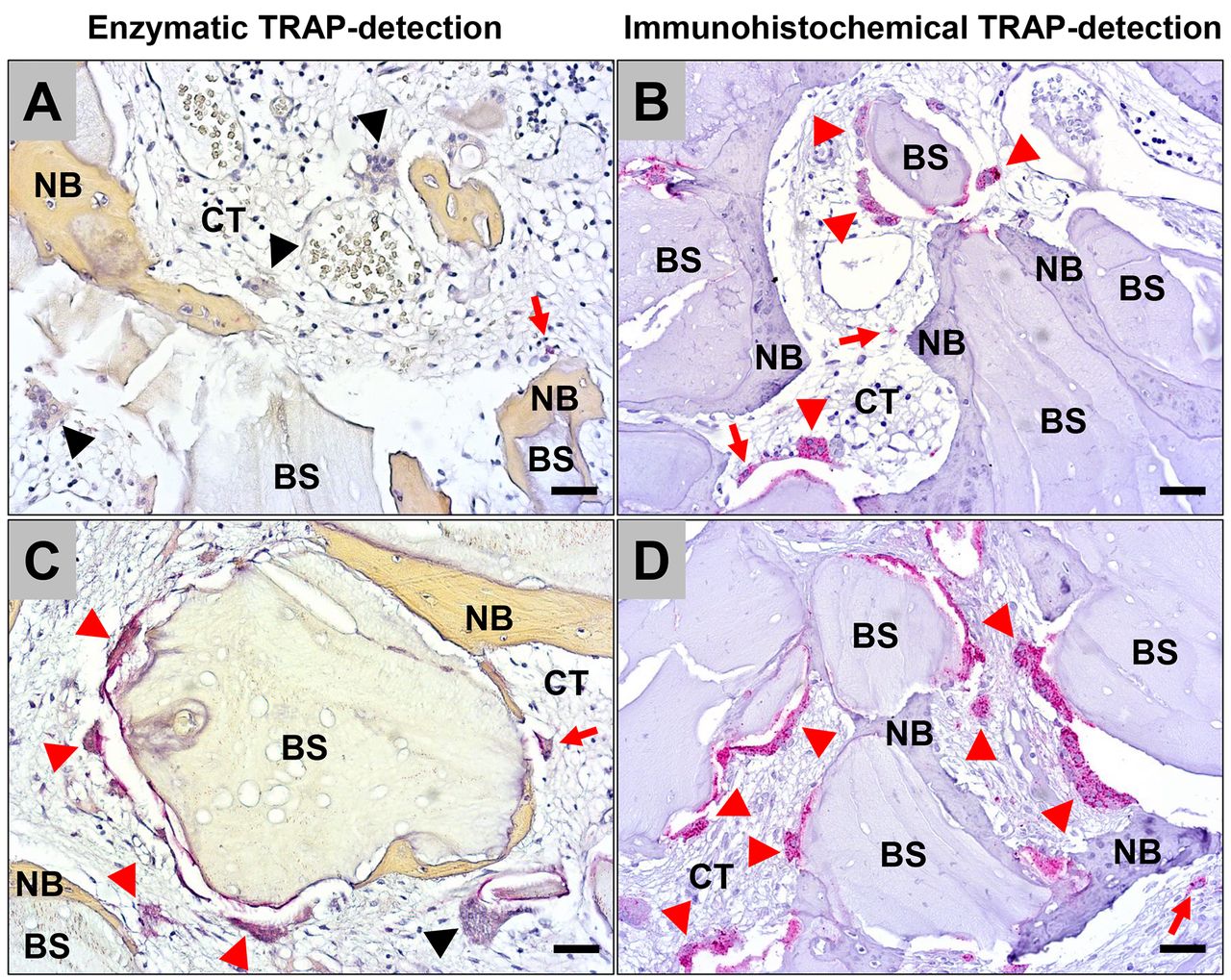
Exemplary microscopic images of the comparison of the enzymatic TRAP-detection method (A and C) and the immunohistochemical TRAP5a-detection method (B and D) within the implantation beds of the xenogeneic bone substitute (BS). (A) In 8 out of 10 biopsies only single cells (red arrows) were TRAP-positive, while most of the cells and especially the biomaterial-associated multinucleated giant cells (BMGCs) (black arrowheads) were TRAP-negative. (B) In contrast, moderate numbers of mononuclear cells (red arrows) and the majority of the BMGCs (red arrowheads) showed signs of TRAP-expression. (C) and (D) Only in one case the enzymatic TRAP detection provided results comparable to those obtained via immunohistochemistry. CT: Connective tissue; NB: newly formed bone tissue. (A and C: enzymatic TRAP-staining, B and D: TRAP5a-immunohistochemistry, 200× magnifications, scalebars=20 μm).
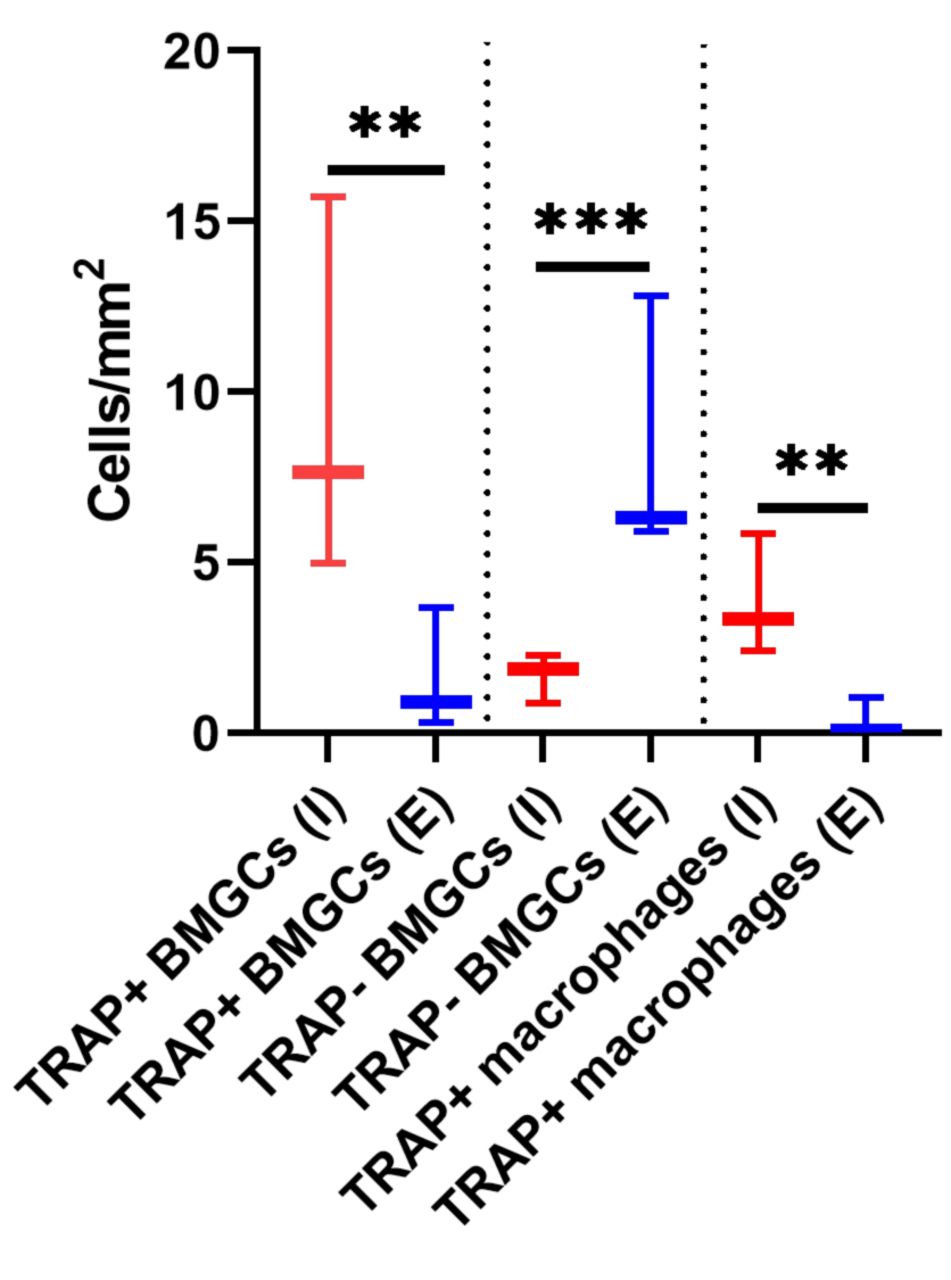
Histomorphometrical results. The histomorphometrical analysis revealed that 9.45±5.59 TRAP-positive BMGCs per mm2 were detectable after immunohistochemical detection, while 1.64±1.79 positive BMGCs/mm2 were identified using the enzymatic TRAP staining (Figure 2). Statistically, these numbers were significantly different (**p≤0.01). Moreover, 1.69±0.72 TRAP-negative BMGCs per mm2 were detected using the immunohistochemical staining, and 8.35±3.86 BMGCs/mm2 were identified using the enzymatic histochemical staining methodology (***p≤0.001).
{kind=link}
{kind=link}
Results of the histomorphometrical measurements of the TRAP-positive and TRAP-negative biomaterial-associated multinucleated giant cells (BMGCs) as well as macrophages expressing TRAP detected via immunohistochemistry (I) and the enzymatic staining methods (E) based on 8 biopsies (**p<0.01 and ***p<0.001).
Additionally, 3.87±1.77 TRAP-positive macrophages per mm2 were detected using the immunohistochemical staining method, and 0.39±0.57 cells/mm2 were found using the enzymatic technique (**p≤0.01).
Discussion
The enzymatic histochemical tartrate-resistant acid phosphatase (TRAP) detection has been a diagnostic tool in pathology, specifically in the diagnosis of metabolic bone diseases (17-20). It was transferred to the field of bone substitute materials (BSM) (13). The staining was assumed to allow for the measurement of the so-called “bioactivity” of multinucleated giant cells that resided at the surface of these biomaterials, which were assumed to be osteoclasts. It was described that this cell type exhibits an osteoclastic activity that is usually correlated with osteoinductivity due to the molecular interaction of the anabolic (osteoblasts) and catabolic (osteoclasts) cells in the context of bone remodeling (18). However, the research in this field revealed within the last years that these multinucleated giant cells seem to be of the foreign body giant cell type and might thereby be inflammatory–in contrast to osteoclasts that are physiological multikaryons (33).
Nevertheless, the enzymatic TRAP staining does not allow for distinction of the TRAP isoenzymes 5a and 5b, which means that the important distinction between the physiological cellular phenotype and inflammatory phenotype cannot be made by this method. In this context, it its noticeable that TRAP5a is a marker for “unphysiological” inflammatory conditions, while TRAP5b is known to be expressed by osteoclasts (21). Moreover, the expression of TRAP5a has been correlated with the presence of reactive oxygen species (ROS) (36, 44). It has been assumed that a redox-active iron ion within the binuclear iron center of the TRAP molecule might have the ability to interact with hydrogen peroxide to produce highly destructive ROS, which means that the expression of TRAP5a can indicate a pro-inflammatory tissue reaction to the implanted biomaterial (32, 45). Thus, the expression of this molecule seems to be directly connected to the degradation process of biomaterials, such as BSM, which explains the necessity of TRAP detection for the analysis of the cellular bioactivity of phagocytes towards a biomaterial.
Based on this knowledge the present study was aimed (a) to analyze the validity of the enzymatic TRAP detection method and (b) to confirm the assumption of the pro-inflammatory phenotype of BMGCs. Thereby, a new antibody against TRAP 5a was used and its validity compared to the routinely carried out enzymatic histochemical staining was examined. The results of this study suggest that the immunohistochemical staining technique of TRAP 5a is more accurate than the enzymatic methodology. That is because significantly more macrophages and BMGCs were found to be TRAP-positive using the immunohistochemical process.
This result may indicate that the enzymatic process is less specific to the isoenzymes as the immunohistochemical staining identified more TRAP-positive cells. Moreover, many studies have reported that the TRAP isoforms have optimal enzyme activities at certain pH values, i.e., 5a is more optimal at 5.2 and 5b at 5.8 (15). This observation suggests that the enzymatic staining process could detect more TRAP-positive cells, since the pH of the straining solutions are around the optimal pH values of TRAP5a and TRAP5b. However, and even though the immunohistochemical staining process has much more fluctuating pH values (from 6 to 9 due to the antibody-retrieval process), this method still resulted in more TRAP-positive cell counts. The increased occurrence of TRAP-negative cells in the enzymatic staining might be a consequence of the loss of enzyme activity due to formalin fixation, tissue processing, and paraffin embedding, which can additionally impact enzyme activity (37, 38). The biopsies go through similar steps during immunohistochemical staining; however, a retrieval step demasks the targeted epitopes. The results of this study suggest that the demasking step in immunohistochemical staining had a major advantageous influence on the accuracy of the TRAP detection. Furthermore, immunohistochemical staining of TRAP5a seems to identify mononuclear macrophages that are under lytic activity more accurately, while the enzymatic staining methodology cannot afford this detection specificity. This observation is a further proof that the immunohistochemical staining of TRAP is more suitable for the investigation of the bioactivity of biomaterials such as BSM. Altogether, this result leads to the conclusion that most of the studies based on the enzymatic TRAP detection method presented inaccurate data about the resorbability of the analyzed BSM, which can be prevented in the future based on the present data.
Another noticeable result of the present study is the detection of TRAP5a expression even in BMGCs. The dichotomy of osteoclasts and BMGCs that were found on the surface of implants has always been of interest (15, 27, 45, 46). Although both cell types share the same monocytic origin, they differ in important factors: i) Osteoclasts are physiological cells that resorb native bone matrix within the process of bone remodeling (47). In contrast, BMGCs as inflammatory cells also exhibit a lytic activity. However, this cell type is degrading and resorbing foreign bodies including many biomaterials such as synthetic bone substitute materials (45, 48, 49). ii) Osteoclasts are involved in the tissue regeneration process via their well described crosstalk with osteoblasts in both physiological and pathological situations (47). Interestingly, BMGCs as “inflammatory cell types” can also be involved in the successful integration of a biomaterial by promoting tissue healing via expression of anti-inflammatory cytokines or signaling molecules such as the vascular endothelial growth factor (VEGF) or in the rejection process mediating encapsulation of a biomaterial/foreign body via fibrosis, and the release of pro-inflammatory cytokines (33). iii) Morphologically, osteoclasts are distinguished from BMGCs by the presence of ruffled borders within the resorption cavity (15, 22). iv) Finally, osteoclasts express TRAP5b and BMGCs are believed to express the “inflammatory” TRAP5a isoform (15, 17, 45).
The results of the present study that show TRAP5a expression in all material-adherent multinucleated giant cells clearly shows that the BSM induced an inflammatory tissue reaction instead of a physiological cell reactivity. Thus, this observation shows that the biomaterial is recognized as a foreign body instead of being integrated in a physiological bone remodeling process – although being of “natural” origin. Moreover, this observation questions the application of the enzymatic histochemical staining of TRAP to the field of bone graft research. Thus, the difference between the two cell types led to the misinterpretation of enzymatic TRAP staining in the context of BSM as BMGCs were seen on the surface of bone grafts and stained positive to TRAP, which was perceived as an “osteoclastic” activity (45, 50). This observation led to the conclusion that BSM results in an upregulation of TRAP expression indicating osteoinductive activity especially because there is an osteoblast-osteoclast intercellular signaling and osteoinductive bone grafts lead to the recruitment/differentiation of cells into osteoblasts (51). It was perceived that these newly recruited osteoblasts would then signal osteoclasts to arrive to the surface of the biomaterial and orchestrate a lytic activity that caused them to stain positive using the enzymatic TRAP staining (50, 51). However, this concept seems to be incorrect and the presence of TRAP-positive BMGCs on the surface of a BSM does not necessarily mean that this biomaterial is indeed osteoinductive.
One proof that contradicts this concept is the fact BMGCs can also exist on biomaterials that are not used for bone regeneration, for instance, in subcutaneous implantation models (48, 52, 53), or the fact that some BSMs did not induce this cell type and are only osteoconductive (54, 55). Another proof is the result of the present examination, where the immuno-histochemical staining of the macrophage-specific isoform TRAP5a, resulted in higher numbers of positive BMGCs adhering to the BSM. This means that the presence of BMGCs on the surface of BSM does not indicate that the biomaterial is osteoinductive. Instead, it suggests that this biomaterial has certain physicochemical characteristics that facilitate the deposition of a protein layer upon implantation, which will consequently induce the formation and attachment of BMGCs (33). This result creates a paradigm shift regarding the interpretation of the TRAP-staining in the context of (bone) biomaterials. The upregulated expression of TRAP should not be seen as an indicator of osteoinductivity but rather an indicator of the lytic and pro-inflammatory activities of macrophages and BMGCs that are residing within the implantation bed of the investigated biomaterial. Nonetheless, an increased expression of TRAP in implantation beds is not necessarily a negative manifestation or a sign of implant failure. Warranted by the new bone growth seen in the analyzed biopsies, it has been shown that macrophages and BMGCs can polarize between pro- and anti-inflammatory phenotypes, which is required for a balanced (bone) tissue growth and wound healing (29). Furthermore, the pro-inflammatory activity of BMGCs has been shown to cause the upregulation of the vascular endothelial growth factor (VEGF), leading to higher vascularization of the implantation bed as one of the essential requirements of successful bone regeneration (24, 56).
In general, the immunohistochemical staining of TRAP seems to be more advantageous in multiple facets. One is the increased sensitivity, suggested by the results of this study, which makes it a more suitable tool in the research of (bone) biomaterials. Another advantage could be that the immunohistochemical staining of TRAP is less time consuming and can be easily incorporated into any laboratory that carries out immunohistochemical-based detections. The only change needed in the process is the primary antibody, while the rest of the process is a standard procedure (39). However, enzymatic histochemical staining is a separate standard procedure that targets an enzyme, specifically tailored to conserve its activity (25, 37). Even though enzymatic histochemical staining is sensitive to tissue fixation via formalin, formalin is the most common medium for chemical fixation and is the most accessible to physicians to directly preserve biopsies (38, 39). An immunohistochemical staining provides an alternative that might not interfere with formaldehyde-based fixation, as a demasking of the epitopes is carried out before application of the primary antibody (39). This technique can also be further utilized to differentiate between cells expressing TRAP5b (osteoclasts) and TRAP 5a (BMGCs/macrophages).
A limitation of the present study is the lack of a TRAP5b detection, which is based on the fact that such antibodies are until now only available for ELISA assays, for instance, but not for immunohistochemistry. Interestingly, Halleen et al. produced an antibody against TRAP5b molecule and successfully detected osteoblastic activity in bone physiology and metabolic diseases (21, 34, 57-59). However, this antibody is still not commercially available. Combination of these two primary antibodies used in immunohistochemistry, the one used to detect TRAP5a and the other used to show TRAP5b expression, can potentially further differentiate between osteoblasts and BMGCs, and finally confirm their dichotomy.
Acknowledgements
The Authors thank Mrs. Annica Pröhl for her excellent technical assistance.
Footnotes
Authors’ Contributions
Conceptualization, M.B.; methodology, D.R., T.F. and M.B.; software, M.B.; investigation, S.A., A.J. and M.B.; resources, D.R., T.F. and M.B.; data curation, S.A. and M.B.; writing – original draft preparation, S.A. and M.B.; writing – review and editing, S.A., D.R., T.F., M.B. and O.J.; visualization, S.A. and M.B.; supervision, M.B.; project administration, M.B. All Authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The Authors declare no conflicts of interest in relation to this study.
- Received June 14, 2022.
- Revision received July 12, 2022.
- Accepted July 14, 2022.
- Copyright © 2022, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY-NC-ND) 4.0 international license (https://creativecommons.org/licenses/by-nc-nd/4.0).