

Abstract
Background/Aim: Sleep loss is proposed as a trigger for manic episodes in bipolar disorder in humans. It has been shown that sleep and wakefulness can affect changes in mitochondrial gene expression, oxidative phosphorylation (OXPHOS) activity, and morphology in the brain. In this study, we investigated alterations in mitochondrial bioenergetic function in the brain of rats after 72-h rapid eye movement sleep deprivation (REM-SD). Materials and Methods: Alterations in the mitochondrial DNA (mtDNA) copy number were detected in the prefrontal cortex and hippocampus through amplification of mitochondrially encoded NADH dehydrogenase 1 (mt-Nd1) gene using quantitative real-time polymerase chain reaction. The expression levels of mitochondrial biogenesis-related proteins such as peroxisome proliferator-activated receptor gamma coactivator 1 alpha (PPARGC1A), cytochrome c oxidase subunit 4I1 (COX4I1) and sirtuin 3 (SIRT3) were assessed using western blot analysis and immunohistochemistry. Results: We found that REM-SD significantly increased the mtDNA copy number in the hippocampus but not in the prefrontal cortex. In addition, REM-SD increased the protein expression of COX4I1 in the hippocampus. Furthermore, we observed manic-like behaviors in rats exposed to 72-h REM-SD. REM-SD increased locomotion in the open-field test and the time spent in open arms in the elevated plus-maze test. Conclusion: REM-SD may induce mitochondrial dysfunction in the brain, which may be involved in the induction of mania.
- Mitochondria
- rapid eye movement sleep deprivation
- hippocampus
- manic-like behavior
Sleep disturbance has been linked to poor cognition, mood problems, and psychiatric disorders (1-5). Sleep deprivation (SD) has been reported to perturb endocrinal and metabolic functions and the immune system of the brain (6-8). Animal studies have suggested that rodents exposed to SD can act as a model of mania (9-11). Rodents exposed to SD show manic-like behaviors, including hyperlocomotion, irritability, hypersexuality, and aggressiveness, as observed during the manic phase in patients with bipolar disorder (BPD) (9-12). The administration of mood stabilizers, such as lithium and valproic acid, which are used for the prevention and relief of the manic phase in BPD, also reduces these manic-like behaviors in rodents experiencing SD (13-16).
Mitochondria play a key role in cellular energy homeostasis and production through adenosine triphosphate (ATP) synthesis by oxidative phosphorylation (OXPHOS). Mitochondria are highly dynamic organelles, undergoing a balance between fission and fusion, to maintain their shape, distribution, and size (17). The mitochondrial dynamics help eukaryote cells adapt to environmental metabolic demand or stress (18). Previous studies have suggested the involvement of mitochondria in sleep and wakefulness. Animal studies have shown changes in mitochondrial gene expression, OXPHOS activity, and morphology in the brain of rodents exposed to SD or spontaneous wakefulness (19-22). Human studies have also shown alterations in mitochondrial function and OXPHOS activity in individuals with SD or insomnia (23, 24). In particular, many studies have reported increased mRNA and protein levels of the mitochondrial electron transport chains (ETC), such as complex IV subunits mitochondrially encoded cytochrome c oxidase I (mt-CO1) and cytochrome c oxidase subunit 4I1 (COX4I1), complex I subunit mitochondrially encoded NADH dehydrogenase 2 (mt-ND2) subunit, and complex V subunit ATP synthase F1 subunit beta (ATP5F1B) during spontaneous wakefulness (3 h) and after total SD (3 h or 12 h) in the brains of mice or rats (19-22). COX activity was also revealed to be increased in rodents with spontaneous wakefulness (3 h) and total SD (3 h or 12 h) (21, 22). Studies have suggested that these increased levels of mitochondrial complex genes/proteins and COX activity are related to increased cerebral energy demands during wakefulness or SD, resulting from increased neuronal activity (21, 22, 25, 26).
Although studies have reported that wakefulness and total SD during a short period (3-12 h) affect mitochondrial function, increasing mitochondrial gene-expression levels and OXPHOS activity (20-22, 27), there is a lack of knowledge about how mitochondrial bioenergetic function is changed during rapid eye movement (REM)-SD over a relatively long period. In this study, we investigated the changes in mitochondrial biogenesis, which is an important process for regulating the number of mitochondria as well as energy metabolism, in the brain of rats exposed to 72-h REM-SD. In the prefrontal cortex and hippocampus, we analyzed the mitochondrial DNA (mtDNA) copy number and protein expression levels of mitochondrial biogenesis-related proteins such as COX4I1; peroxisome proliferator-activated receptor gamma coactivator 1 alpha (PPARGC1A), which is a positive regulator of mitochondrial biogenesis and respiration (28); and sirtuin 3 (SIRT3), which regulates the acetylation of PPARGC1A (29).
Materials and Methods
Animals. Male Sprague–Dawley rats (8 weeks of age) were obtained from Central Lab. Animal Inc. (Seoul, Republic of Korea). The rats were maintained under a controlled 12-h light/dark cycle (lights on at 7:00 a.m. and off at 7:00 p.m.), temperature (22±2°C), and humidity (50±10°C), with food and water freely available. All animal experiments were conducted in accordance with the animal care guidelines of the National Institute for Health (NIH) Guide and approved by the Animal Care and Use Committee at Kyung Hee University [KHUASP(SE)-18-083].
REM-SD. REM-SD was carried out using the modified multiple platform method (30, 31). Rats were randomly divided into control (n=12) and SD groups (n=12). The rats of the SD group were placed in cages with six small platforms (6 cm diameter) (n=3 per chamber). The cages were filled with water up to approximately 2 cm below the surfaces of the platforms. When the rats entered REM sleep and touched the water due to diminished muscle tone, they were awakened. The rats of the control group were placed in cages with six large platforms (15 cm diameter), also filled with water (n=3 per chamber). On the large platforms, the rats were able to sleep undisturbed; however, the cages were not large enough for rats to walk around freely because of the surrounding water. Food and water were available ad libitum. All rats remained in the cages for 72 h.
Open-field test (OFT). After REM-SD for 72 h, locomotor activity was measured using in-house custom-made black open-field Plexiglas boxes (90×90×50 cm), as previously described (32). For the test, each rat was put in the center of the apparatus and was allowed to explore the open field. Locomotor behavior was recorded for 5 min (test session) using a video camera. Locomotion was analyzed using ImageJ software (National Institutes of Health, Bethesda, MD, USA) and measured as the travelled distance in meters. The testing arena was set up in quiet surroundings, and the test was conducted under dim light. The boxes were cleaned with 70% ethanol after each test.
Elevated plus-maze (EPM) test. After the OFT, EPM testing was performed in the same manner as in a previous study (12). The EPM was composed of two open arms (50×10 cm) and enclosed arms (50×10×30 cm, without roof) and was elevated to a height of 50 cm. Rats were habituated in the testing room for at least 20 min; then, the rats were placed into the center of a cross-maze. Behavior was recorded using a video camera. For 5 min (test session), the time spent and the numbers of entries into the open or closed arms were recorded. The maze was cleaned with 70% ethanol after each test.
Sample collection. After the behavioral tests, the rats were sacrificed. For mtDNA copy number assay and western blot analysis, the prefrontal cortex and hippocampus from each rat were dissected out and stored at -80°C until analysis. For immunohistochemistry, the rats were transcardially perfused with 0.05 M phosphate-buffered saline (pH 7.4) and then with 4% paraformaldehyde solution. The fixed brain samples were stored in 30% sucrose for 2 days, then sectioned in the coronal direction (40 μm) on a freezing microtome at -26±2°C.
Genomic DNA isolation and mtDNA copy number assay. The total genomic DNA was isolated using a DNeasy Blood and Tissue kit (Qiagen, Hilden, Germany) from brain tissue according to the manufacturer’s instructions. The mtDNA copy number was determined via quantitative real-time polymerase chain reaction (qRT-PCR). For qRT-PCR, each genomic DNA sample was mixed with primers (Bioneer, Deajeon, Republic of Korea) and SYBR Green Supermix (Bio-Rad, Hercules, CA, USA). The expression of hemoglobin subunit beta gene (Hbb) in the nucleus was used as a control, and the mt-Nd1 gene was used as a target mitochondrial gene. The sequences of the primers used for amplification of the target genes were as follows: 5’-CAGTACTTTAAGTTGGAAAC- 3’ and 5’-ATCAACATAATTGCAGAGC-3’ for Hbb; 5’- TTATCCTCTTATCCGTCCTC-3’ and 5’-TGTTAAGTCGAAGG GAGC-3’ for Nd1. The target genes of the mixture were amplified using a CFX96™ Real-Time system (Bio-Rad, Hercules, CA, USA) with the following protocol: 95°C for 10 min, followed by 40 cycles of 95°C for 15 s, and 57°C for 1 min. The relative mtDNA copy number was determined by the 2-ΔΔCt method (33).
Western blotting. Total proteins from the prefrontal cortex and hippocampus were extracted using RIPA buffer (ATTO, Tokyo, Japan). The concentration of protein samples was measured via a bicinchoninic acid assay. Twenty micrograms of cytosolic protein were separated on 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis gels and then transferred to nitrocellulose membrane (Pall, Port Washington, NY, USA). The membranes were blocked with 5% skim milk in Tris-buffered saline with Tween® 20 (TBST) buffer for 1 h at 4°C, then incubated with primary antibodies to COX4I1, SIRT3, b-actin (ACTB; Cell Signaling Technology, Danvers, MA, USA), and PPARGC1A (Millipore, Billerica, MA, USA) at 4°C overnight. The membranes were washed with TBST buffer and incubated with horseradish peroxidase-labeled secondary antibodies (Santa Cruz Biotechnology, Dallas, TX, USA) for 1 h. The protein bands were visualized using West Femto Maximum Sensitivity Substrate (Thermo Fisher Scientific, Waltham, MA, USA). The amount of protein expression was detected by exposure using X-ray film and analyzed using ImageJ software (National Institutes of Health).
Immunohistochemistry. Free-floating tissue sections of the hippocampus were selected and treated with 1% dihydrogen dioxide. Then the sections were blocked with 10% normal horse serum (Vector Laboratories, Burlingame, CA, USA) for 1 h and probed with mouse primary antibody to COX4I1 (Cell Signaling Technology) overnight at 4°C. The sections were incubated with biotinylated horse antimouse secondary antibody for 1 h, then with the Vector Elite ABC Kit (Vector Laboratories) for 30 min. After staining with diaminobenzidine substrate (Vector Laboratories), the sections were mounted onto gelatin-coated slides and covered with coverslips using Permount (Fisher Scientific, Pittsburgh, PA, USA). Immunostained images were visualized under a light microscope (Olympus BX43; Shinjuku, Tokyo, Japan). COX4I1 immunoreactivity was quantified using ImageJ (National Institutes of Health).
Statistical analysis. The results are expressed as the means±standard errors of the mean (SEMs). For statistical analysis of the results, IBM SPSS Statistics 26 (IBM, Armonk, NY, USA) was used. The normality of the variable distribution of data was assessed by the Shapiro-Wilk test. All data showed a normal variable distribution, thus, differences between groups were compared using the unpaired t-test. Values of p<0.05 were considered as statistically significant.
Results
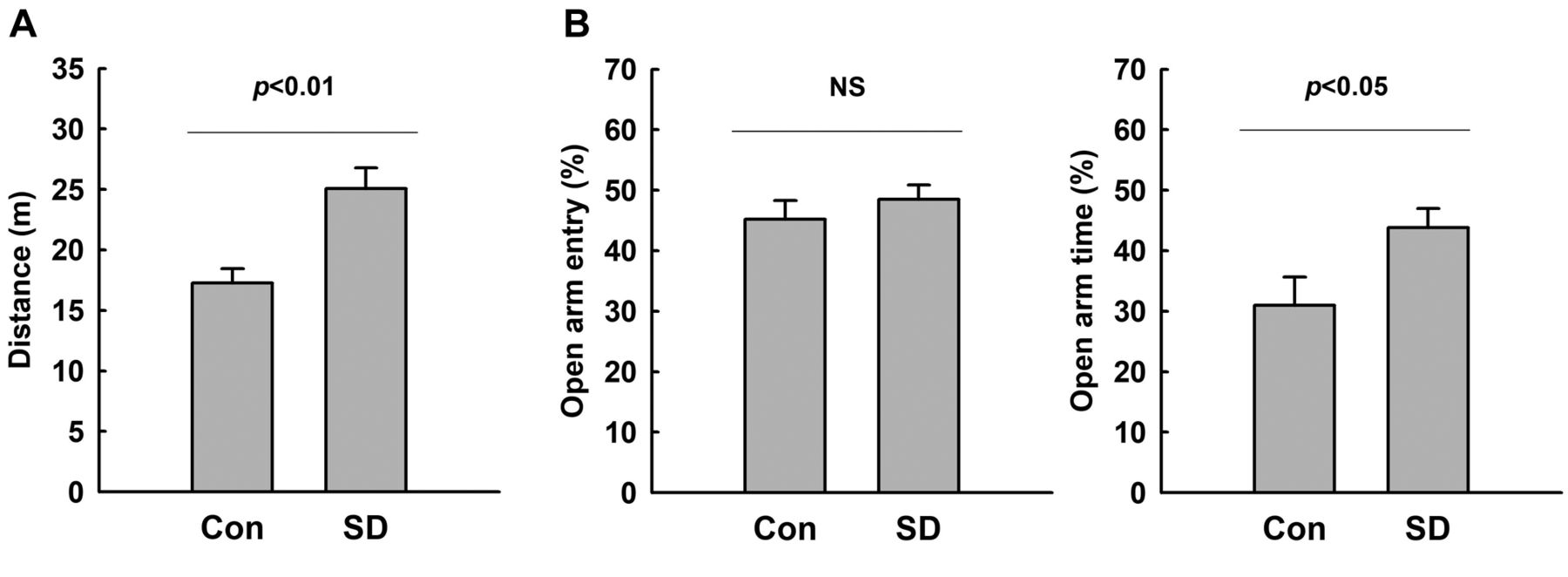
Increased manic-like behaviors of rats exposed to REM-SD. The hyperactive- and impulsive-like behaviors of rats exposed to REM-SD were examined using the OFT and EPM test. As shown in Figure 1A, the OFT showed increased locomotor activity in rats exposed to REM-SD as compared to control rats. In REM-SD rats, the total travel distance was increased by approximately 30%.
In the EPM test, increased frequency of open-arm entries and increased time spent in open arms are considered a sign of impulsive-like behavior (34, 35). We did not observe a significant difference in the number of open-arm entries between REM-SD and control rats. However, REM-SD rats showed a significantly increased time spent in the open arms as compared to control rats (Figure 1B). These results indicate that hyperactive- and impulsive-like behaviors are increased in rats exposed to REM-SD.

Manic-like behaviors in rats after sleep deprivation (SD). Manic-like behaviors in rats after SD were measured using the open-field test (A) and the elevated plus-maze test (B). Hyperactive locomotion was indicated by total travel distance in the open-field test. Impulsive behavior was indicated by the percentage of open-arm entries and of time spent in the open arms. A rat which fell from the apparatus during the elevated plus-maze test was excluded from analysis. The results are presented as the mean±standard error of the mean (n=11-12). The difference between the SD and control groups was determined via unpaired t-test. NS: Not significant, p≥0.05.
Increased mtDNA copy number in the hippocampus of rats exposed to REM-SD. To assess the tissue effect of REM-SD on mitochondrial biogenesis, we compared the relative mtDNA copy number in the prefrontal cortex and hippocampus. The ratio of mt-Nd1 gene in mtDNA to nuclear DNA (Hbb) was increased in the hippocampus of REM-SD rats (Figure 2, p<0.05). However, in the prefrontal cortex, a statistically significant difference in the mtDNA copy number was not detected between control and REM-SD rats.
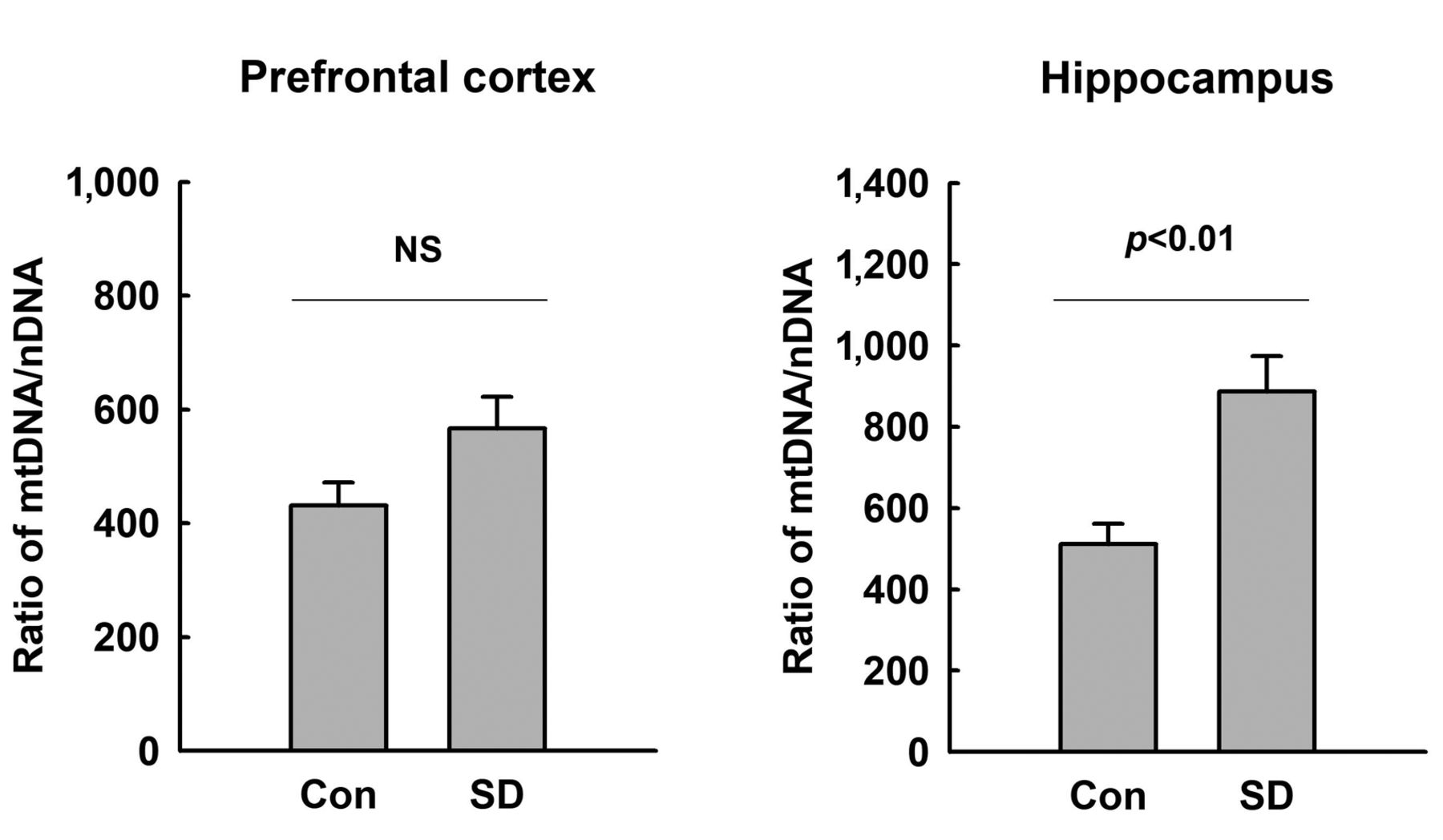
Relative quantification of the mitochondrial DNA (mtDNA) copy number - in the brains of rats after 72-h sleep deprivation (SD). The relative expression of mitochondrially encoded NADH dehydrogenase 1 (mt-Nd1) was measured against that of hemoglobin subunit beta gene (Hbb) in the hippocampus and prefrontal cortex tissue by quantitative real-time polymerase chain reaction. Data are presented as the mean±standard error of the mean (n=8). The difference between the SD and control groups was determined via unpaired t-test. NS: Not significant, p≥0.05.
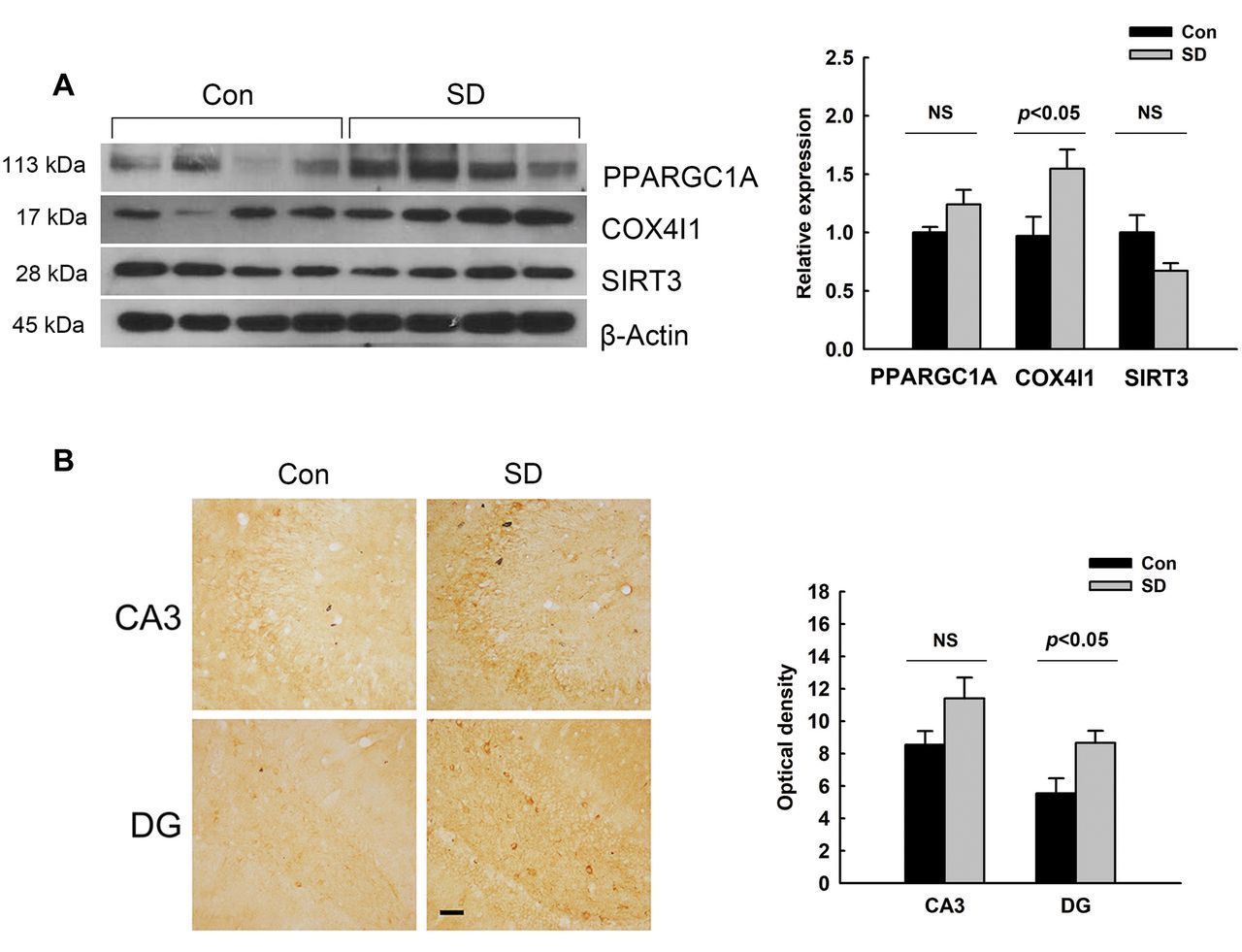
Increase in COX4I1 protein expression in the hippocampus of rats exposed to REM-SD. We assessed the protein expression levels of PPARGC1A, COX4I1, and SIRT3 in the hippocampus of REM-SD and control rats. As shown in Figure 3A, the expression levels of PPARGC1A and SIRT3 were not significantly changed by REM-SD. However, REM-SD did up-regulate protein expression of COX4I1. The immunostaining results also showed that COX4I1 expression was elevated in the hippocampus of rats subjected to REM-SD as compared to control rats (Figure 3B). COX4I1 immunoreactivity was observed in the CA3 and dentate gyrus regions of the hippocampus. In particular, immunostaining for COX4I1 was potently elevated in the dentate gyrus of rats exposed to REM-SD.

{kind=link}
{kind=link}
{kind=link}
Cytochrome c oxidase subunit 4I1 (COX4I1) expression in the hippocampus of rats after sleep deprivation (SD). A: The expression levels of peroxisome proliferator-activated receptor gamma coactivator 1 alpha (PPARGC1A), COX4I1 and sirtuin 3 (SIRT3) protein were determined via western blotting in the hippocampus of control and SD-treated rats. The histogram shows the protein expression levels normalized to that of β- actin (n=4-8). β-Actin expression was used as an internal control. B: COX4I1 immunoreactivity was also observed in the hippocampus of control and SD-treated rats. The optical density of COX4I1 immunoreactivity is shown as the mean±standard error of the mean (n=4). Scale bars=50 μm. The difference between the SD and control groups was determined via unpaired t-test. NS: Not significant, p≥0.05. DG: Dentate gyrus.
Discussion
SD increases energy expenditure in the brain (36-38). Thus, it has been shown that energy metabolism is elevated, and various mitochondrial genes/proteins are overexpressed in SD in order to compensate for this increased energy expenditure (22, 36-38). Indeed, previous studies have shown an up-regulation in mRNA and and/or protein expression levels of complex IV subunits mt-CO1 and COX4I1, complex I subunit mt-ND2, and complex V subunit ATP5F1B in the brain of mice and rats exposed to total SD for 3 h or 12 h (19-22). COX activity was also observed to be increased in the brain of 3-h or 12-h total SD rats (21, 22). On the other hand, another study (39) showed that expression levels of mt-Co1 and Cox4I1 mRNA were not changed in the brain of 8-h total SD rats as compared to rats allowed to sleep undisturbed. The researchers suggested that the increase in mt-Co1 and COX4I1 transcripts was transient and not consistent for up to 8 h of SD (39).
An effect of SD on mitochondrial genes and proteins has also been observed in REM-SD animals during a relatively long period of SD. Streck et al. (15) reported that 96-h REM-SD in mice decreased the activity of succinate:cytochrome c oxidoreductase and COX in the prefrontal cortex, hippocampus, striatum, and/or cerebral cortex (15). Andreazza et al. (40) found that activity of NADH:cytochrome c oxidoreductase was reduced in the striatum and hypothalamus of mice after 72-h REM-SD. These results indicate that REM-SD for a relatively long period reduces the activity of mitochondrial complexes in the brain. However, in our study, we found that 72-h REM-SD increased the mtDNA copy number and the protein level of COX4I1 in the hippocampus. In the context of declining mitochondrial function, mitochondrial biogenesis has been suggested to represent an attempt of cells to increase their aerobic set point, or to maintain a pre-existing aerobic set point (41). Indeed, in the aged brain, increased amounts of mtDNA have been considered a compensatory response to a gradual decline in mitochondrial function (falling OXPHOS capacity) (41-43). Given our results and previous reports, the increase in the mtDNA copy number and expression level of COX4I1 in the brain of rats after REM-SD may be a compensatory response to the reduced activities of complexes of the mitochondrial ETC. SD over a long period (72 h or 96 h) may steadily increase the energy demand in the brain. Despite the steadily increased energy demand, the declining capacity of mitochondrial complexes may promote mitochondrial biogenesis in the brain of REM-SD rats.
Sleep disturbance is commonly observed in patients with mood problems and psychiatric disorders, including BPD (2-5). Sleep loss is proposed as a trigger of manic episodes in BPD in humans (5, 44, 45). In rodents, SD is used as an animal model of mania. Rodents exposed to REM-SD have shown manic-like behaviors such as hyperactivity and impulsivity, increasing the locomotor activity in the OFT, and the frequency of open-arm entries/time spent in the open arms in the EPM test (12-16). In our study, we also observed increased locomotion after REM-SD in the OFT and increased time spent in open arms in the EPM test.
Altered mitochondrial function has attracted attention as a biomarker in stress adaptation and psychiatric disorders. Previous studies have shown alterations in mitochondrial genes and the mtDNA copy number in the postmortem brain of patients with BPD, along with impaired energy metabolism and altered activity of ETC complexes (46-51). Some studies have also reported a reduced level of the gene encoding the complex I subunit NADH:ubiquinone oxidoreductase core subunit V2 (NDUFV2) in the postmortem brain of patients with BPD (49, 50). However, a previous study showed a reduced mRNA level of NDUFV2 in lymphoid cells extracted from patients with BPD, suggesting that the reduction may be due to the promoter polymorphism associated with BPD (52). In addition, Iwamoto et al. showed global up-regulation of mitochondrial genes in the postmortem brain of patients with BPD. They suggested that the down-regulation of mitochondrial genes may be caused by drug treatment. They also found that mitochondrial genes, including genes encoding complex 1-5, were up-regulated in the postmortem brain of medication-free patients with BPD, whereas those were reduced in patients receiving medication (51). These studies indicate that mitochondrial dysfunction may be related to the pathophysiology of BPD, and mitochondrial biogenesis may be induced in the brain of patients with BPD. Our study using a manic-like animal model of rats exposed to REM-SD may also support the involvement of mitochondrial dysfunction in the pathophysiology of BPD.
Our study has some limitations. We used only eight samples per group in the analysis of the mtDNA copy number and western blot, and four samples per group for the immunohistochemistry. In addition, we assessed just three target proteins. Due to the small number of samples and target proteins, we cannot clearly identify the signaling pathways which might lead to mitochondrial biosynthesis or factors related to mtDNA replication. Moreover, we analyzed mitochondrial biogenesis only in the prefrontal cortex and hippocampus. In the future, to evaluate SD-related neuropsychiatric disorders such as BPD, further studies on brain mitochondrial function changes in various signaling pathways associated with changes in energy metabolism need to be considered in more various brain regions.
In conclusion, we found that REM-SD increased the mtDNA copy number and protein expression of complex IV subunit COX4I1, in the hippocampus of rats, together with inducing manic-like behaviors. These results suggest that REM-SD may induce mitochondrial dysfunction in the brain, which may play a role in the pathophysiology of BPD. To our knowledge, our study is the first report of a change of the mtDNA copy number as a marker for mitochondrial dysfunction and mitochondria biogenesis in an SD animal model.
Acknowledgements
The Authors thank Miss Eun Hye Jang for help in the experiments and data collection. This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (2020R1F1A1068990, and 2020R1A2C1009499).
Footnotes
Authors’ Contributions
SAK: conceived and designed research; HJP and SK: carried out all experiments; HJP and SAK performed data analysis; All Authors drafted revised the article. All Authors read and approved the final article.
Conflicts of Interest
The Authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
- Received March 31, 2022.
- Revision received May 13, 2022.
- Accepted May 18, 2022.
- Copyright © 2022, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY-NC-ND) 4.0 international license (https://creativecommons.org/licenses/by-nc-nd/4.0).