

Abstract
Background/aim: Dermal papilla cells (DPCs) regulate hair follicle development. We aimed to investigate the effect of scoparone from Dendrobium densiflorum on DPCs in the induction of stem cell properties and pluripotency-related proteins. Materials and methods: DPC viability was evaluated by the MTT assay. Apoptosis or necrosis of DPCs was determined by Hoecsht33342/PI nuclear staining analysis. Expression of OCT4, NANOG and SOX2 genes was determined using Real-Time Polymerase Chain Reaction (PCR). Immunocytochemistry and western blot analysis were performed to determine pluripotency related proteins. Results: Scoparone increased the expression of pluripotency related transcription factors SOX2 and NANOG, while it had minimal effects on OCT4 levels. Scoparone exerted its stemness-enhancing activity through the up-regulation of Akt-dependent inhibition of GSK3β, resulting in increased cellular levels of β-catenin. Conclusion: Our results show a potential novel activity and mechanism of action of scoparone on human DPCs that could facilitate the development of hair enrichment approaches.
- Scoparone
- dermal papilla
- pluripotency
- stemness
- NANOG
- SOX2
Hair disorders like androgenetic alopecia frequently affect the overall well-being and appearance of an individual; there is now ongoing intensive research covering hair cell biology and the discovery of new compounds benefiting hair growth and maintenance (1). In the mesenchymal compartment of hair follicles, dermal papilla cells (DPCs) at the base play a role as a key signaling center in controlling hair follicle development and sustaining the growing phase of the hair cycle (2, 3). DPCs are also recognized as multipotent stem cells and it has been shown that their stemness is linked to their ability in forming hair follicles (4). In recent years, the Wnt/β-catenin signaling has been demonstrated to provide strong regulatory functions on hair follicle morphogenesis and hair regeneration (5-7). Wnt/β-catenin is known to be an important coregulator of the three master transcription factors (OCT4, NANOG, and SOX2) that promote self-renewal of embryonic stem cells (8-10).
The up-stream regulatory control of Wnt/β-catenin signaling that gathered attention for hair treatment therapeutic approaches is the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (Akt) axis (11). The induction of PI3K/Akt inhibits the ubiquitin-proteasomal degradation of β-catenin by inactivating GSK3β(12), resulting in the increase of cellular levels of β-catenin (12). In addition, PI3K/Akt and Wnt/β-catenin signals are known to facilitate cell growth and survival (13). Cultivation of human DPCs in the presence of a glycogen synthase kinase-3β inhibitor resulted in the induction of Wnt/β-catenin pathway leading to stable hair inductive ability (14). Likewise, another study has demonstrated that basic FGF could induce hair growth (15, 16). Wnt/β-catenin has been found to be an important signaling molecule for hair growth and has the ability to promote the three master transcription factors that enhance self-renewal in DPCs (3, 17); it is therefore likely that Wnt β-catenin signaling will have a positive role in the stemness of DPCs.
Scoparone is a coumarin derivative (Figure 1A) isolated from the Dendrobium densiflorum (Figure 1B). Previous studies have revealed many pharmacological beneficial effects, such as promoting bilirubin clearance as well as anticoagulant, anti-inflammatory, anti-allergic and antioxidant activities (18). However, there are no studies focusing on the effects of scoparone on the stemness of DPCs. Therefore, the aim of this study is to investigate the effects of scoparone on the stemness of dermal papilla cells and to explore its possible mechanism.

Chemical structure of Scoparone (A). Dendrobium infundibulum plant (B). Cytotoxic effects of scoparone on immortalized dermal papilla cells were investigated using the MTT assay (C). Characteristic DPCs treated with 0.25, 5 and 10 μM of scoparone for 24h, visualized under microscopy (D). Scoparone did not induce cellular apoptosis. DPCs were treated with scoparone (0-100 μM) for 24 h. Hoechst 33342 and propidium iodide (PI) were added. Cells were visualized under an inverted fluorescent microscope (E).
Materials and Methods
Compound and reagents. Scoparone was isolated from D. densiforum. Dendrobium densiflorum Lindl. (Orchidaceae) was purchased from Chatuchak market, Bangkok, Thailand in November 2012. Plant identification was performed by Dr. Boonchoo Sritularak. A voucher specimen (BS-112555) is at the Department of Pharmacognosy and Pharmaceutical botany, Faculty of Pharmaceutical Sciences, Chulalongkorn University, Thailand. The whole plant of D. densiflorum (600 g) was dried and powdered, and then macerated with MeOH to give 30 g of MeOH extract after removal of the solvent. The MeOH extract was fractionated by vacuum liquid chromatography (silica gel, hexane-EtOAc, gradient) to give 8 fractions (A-H). Fraction E (2.0 g) was separated by column chromatography (silica gel, hexane-EtOAc, gradient) and further purified on Sephadex LH-20 (acetone) to give Scoparone (61 mg). The structure of scoparone was determined by comparison of its NMR data with previously reported values. (19) Fetal bovine serum (FBS), penicillin/streptomycin, L-glutamine, phosphate buffered saline (PBS) and trypsin were purchased from GIBCO (Grand Island, NY, USA). Propidium iodide (PI) and Hoechst 33342 were obtained from Molecular Probes, Inc. (Eugene, OR, USA). 3-(4,5-dimethyl-thiazol-2-yl)-2,5-Diphenyl tetrazolium bromide (MTT) was obtained from Sigma Chemical, Inc. (St. Louis, MO, USA). Antibody for Akt, activated Akt (phosphorylated Akt at Ser 473), total Akt, inactivated glycogen synthase kinase 3β (phosphorylated GSK3β at Ser 9), GSK3β, GAPDH, and peroxidase-conjugated secondary antibodies were purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
Cell culture. Immortalized dermal papilla cells (DPCs) were acquired from Applied Biological Materials Inc. (Richmond, BC, Canada). DPCs were cultured in Dulbecco’s modified Eagle’s medium (DMEM) (Gibco, Grand Island, NY, USA) supplemented with 10% FBS, 100 units/ml penicillin/streptomycin and 2 mM L-glutamine. The cells were incubated at 37°C with 5% CO2, in a humidified incubator. For subcultures, cells were washed by 1× PBS and detached using a 0.25% trypsin and 0.53 mM EDTA solution.
Cytotoxicity assay. Cells were seeded in 96-well plates at a density of 1×104 cells/well. They were then treated with various concentrations of scoparone for 24 h. DMEM was removed and 100 μl of DMSO were added to dissolve the formed formazan crystals. Color intensity of the crystal formazan product was measured by spectrophotometry at 570 nm using a microplate reader (Anthros, Dur ham, NC, USA); the percentage of cell viability was calculated in relation to untreated cells.
Cell apoptosis assay. After treatment, apoptotic cell death was determined using nuclear staining with Hoechst 33342 and PI. Cells were stained with 10 μM of Hoechst 33342 and 5 μM of PI for 30 min at 37°C. Cells were then visualized and imaged using a fluorescence microscope (Olympus DP70, Melville, NY, USA). Three random fields were captured.
Quantification of DNA by Real-Time Polymerase Chain Reaction (PCR). DPCs were seeded in 6-well plate at a density of 7×105 cells/ml for 24 h. Then, cells were treated with scoparone at concentrations of 0, 2.5, 5 and 10 μM. Cell pellets were collected for RNA extraction by trypsinization. Cells were extracted of RNA by QIAGEN RNeasy mini kit, according to the manufacturer’s instructions. cDNA was then synthesized by the extracted RNA by following the QuantiTect Reverse Transcription protocol kit. cDNA (100 ng/well) was used as a template in qPCR reactions with oligonucleotides specific for the genes of interest (Figure 2A). The relative values were calculated by target genes to the housekeeping gene.

Primer sequences of pluripotency related genes of interest: OCT4, NANOG and SOX2 (A). Scoparone increased the expression of NANOG and SOX2. DPCs were treated with various concentrations of the compound (0, 2.5, 5, 10 μM) for 24 h and OCT4, NANOG and SOX2 expression was measured by Real-Time Polymerase Chain Reaction (PCR). Scoparone at concentrations of 5 and 10 μM significantly increased SOX2 and NANOG expression in DPCs, compared to untreated controls (p<0.05) (B). Data represent mean±SEM (n=3).
Western blot. After treatment (12 and 24 h), the cells (7×105) were washed with cold PBS twice and incubated on ice with lysis buffer containing 20 mM Tris-HCl (pH 7.5), 150 mM sodium chloride (NaCl), 10% glycerol, 1 mM sodium orthovanadate, 50 mM sodium fluoride, 1 mM phenylmethylsulphonyl fluoride and a protease inhibitor cocktail (Roche Molecular Biochemicals, Indianapolis, IN, USA) for 30 min. Cell lysates were collected, and the protein content was determined using the BCA protein assay kit (Bio-Rad Laboratories). Approximately 25 μg of protein from each sample was denatured at 95°C for 5 min and loaded onto 10% SDS-polyacrylamide gels. After gel separation, the proteins were transferred onto 0.45 μm Polyvinylidene fluoride (PVDF) membranes (Bio-Rad), and the transferred membranes were incubated with blocking buffer [5% non-fat dry milk in TBST (25 mM Tris-HCl (pH 7.5), 125 mM NaCl, 0.05% Tween 20)] for 1 h and subsequently incubated with the appropriate specific primary antibody at 4°C overnight. Membranes were then washed 3 times for 5 min with TBST and incubated with horseradish peroxidase (HRP)-conjugated anti-rabbit or anti-mouse IgG isotype-specific secondary antibodies in blocking buffer for 2 h at room temperature. Membranes were then washed again with TBST (3 times for 5 min). The Super Signal West Pico Chemiluminescent substrate (Thermo Scientific, Rockford, IL, USA) was used to detect immune complexes and the signal was quantified by densitometry, using ImageJ version 1.52 (National Institute of Health, Bethesda, MD, USA).
Immunocytochemistry. Cells were pretreated with a PI3K inhibitor (50 μM of LY294002 or 5 μM of wortmannin). 10 μM of Scoparone were then added as described in Figure 3. After treatment, cells were fixed with 4% (w/v) paraformaldehyde for 10 min. The cells were then permeabilized by 0.5% (v/v) Triton-X for 5 min. Cells were incubated with 10% (w/v) FBS for 1 h. Antibodies against β-catenin (1:100) and SOX2 (1:100) (InvivoGen, San Diego, CA, USA) were added and incubated for 10 h at 4°C. Cells were incubated with FITC (Abcam, Cambridge, UK) conjugated Goat Anti-Rabbit secondary antibody (1:250). Nuclei were stained with Hoechst 33342 for 15 min in the dark. Cells were visualized by fluorescence microscopy (Nikon model ECLIPSE Ts2-FL, Tokyo, Japan).
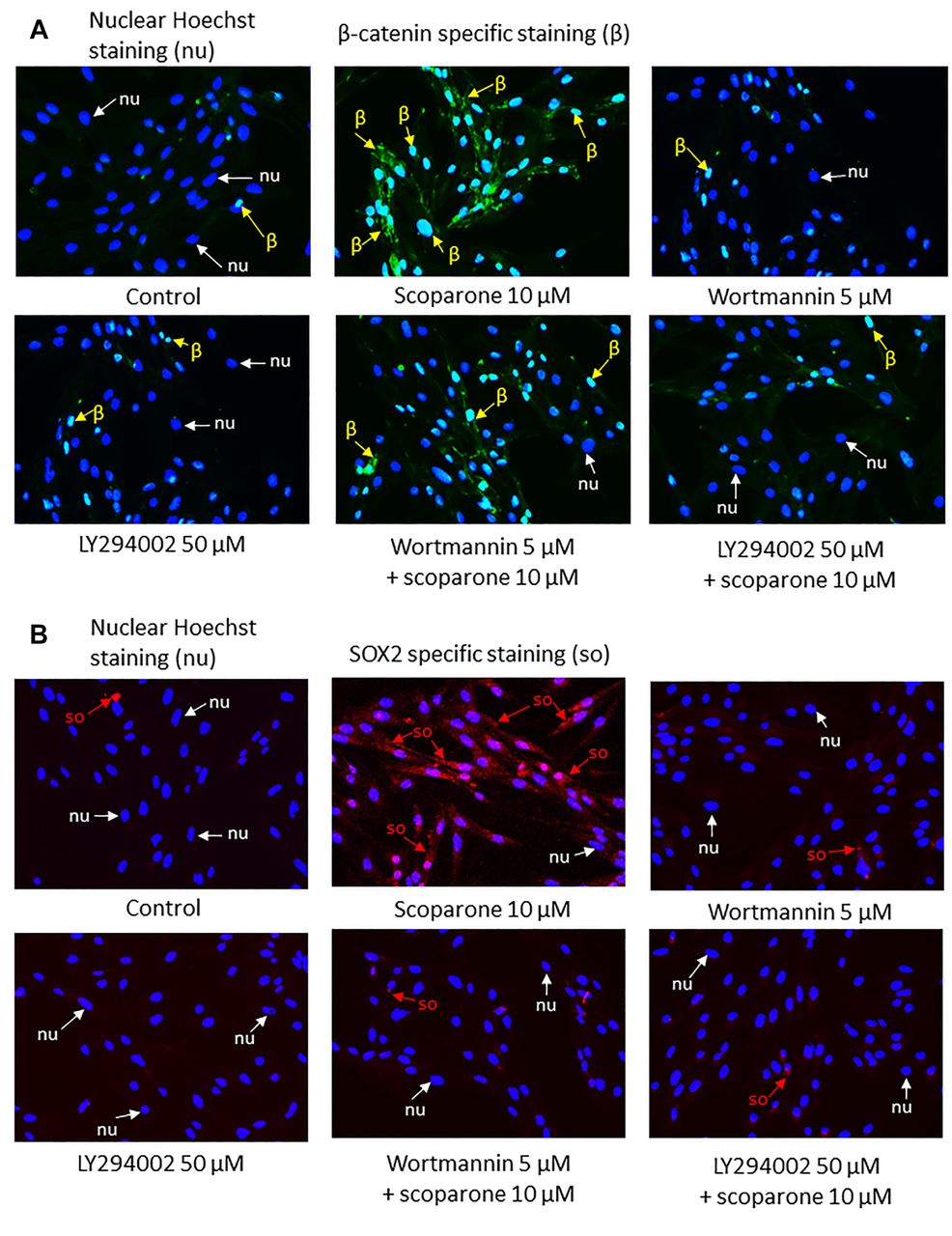
Scoparone increased the expression of β-catenin and SOX2 via the PI3K/Akt pathway. Cells were pretreated with PI3K inhibitors and the expression of β-catenin (A) or SOX2 (B) was measured by immunocytochemistry. Scoparone alone induced expression of β-catenin and SOX2, compared to controls. In the presence of PI3K inhibitors (LY294002 or wortmannin), β-catenin and SOX2 were found to be reduced (A, B).
Statistical analysis. Statistical differences between multiple groups were analyzed using an analysis of variance (ANOVA) followed by the Tukey HSD post-hoc test. The data is presented as the mean±standard error of the mean (SEM) of three independent experiments with three replicates per experiment. All p-values of less than 0.05 were considered as statistically significant.
Results
Cytotoxicity of scoparone on human dermal papilla cells. To test the effect of scoparone, derived from Dendrobium densiflorum (Figure 1A, B), we first determined the cytotoxic concentrations of the compound in immortalized human dermal papilla cells (DPCs). Briefly, 80% confluent DPCs were treated with scoparone (0, 2.5, 5, 10, 25, 50 and 100 μM) and cell viability was determined after 24 h by MTT viability assay. Results showed that scoparone at concentrations up to 100 μM was considered non-toxic to the cells (Figure 1C). As MTT assay determined the viability of the cells via monitoring function of mitochondrial enzymes, we next confirmed the non-toxic concentrations of the compound by other means. Hoechst 33342 and PI staining were utilized to detect the direct apoptotic and necrotic cell death, respectively. Apoptotic cells that exhibited condensed or fragmented DNA in the nucleus were barely detected in response to 0-100 μM scoparone. Results suggested that scoparone at concentrations of 0-100 μM did not induce cell death, as indicated by normal nuclei stained with Hoechst and negative PI staining (Figure 1E).
Scoparone increases stem cell transcription factors. Pluripotent stem cell transcription factors are known to exert a direct impact on stemness of stem cells and are widely recognized as positive stem cell markers. In order to evaluate the scoparone’s effect on stemness, DPCs were treated with the compound at 0, 2.5, 5, 10 μM for 24 h and were then subjected to an analysis of pluripotency related genes (OCT4, NANOG, and SOX2) by real-time polymerase chain reaction (PCR). Scoparone at concentrations of 5 and 10 μM significantly increased SOX2 and NANOG expression in DPCs compared to controls (p<0.05, Figure 2B).
Scoparone increases stemness of DPCs via the PI3K/Akt pathway. Signaling via the PI3K/Akt was known to have crucial roles in the self-renewal of embryonic stem cells (20). In addition, treatment of DPCs with scoparone significantly augmented the stem cell-related transcription factors as shown above. Since Wnt/β-catenin signaling has been shown to be the most important signaling in hair regeneration and the β-catenin stability in the cells is tightly regulated by the function of Akt via its inhibition on GSK3β (21, 22), we aimed to investigate the potential regulatory activity of scoparone on the PI3K/Akt/β-catenin axis in DPCs. The cellular level of active Akt (phosphorylated at Ser 473), total Akt, β-catenin, inactivated GSK3β (phosphorylated at Ser 9) and total GSK3β was evaluated by western blot analysis.
At 12 h of treatment, scoparone significantly affected Wnt/β-catenin related proteins compared to untreated control cells. In the cells treated with 2.5, 5 and 10 μM of the compound, as opposed to controls, phosphorylated Akt was found to increase 1.53-, 1.38- and 1.64-fold (p<0.05, <0.05 and <0.01), respectively. Phosphorylated GSK3β increased by 2.12-, 1.62- and 2.14-fold (p<0.01, 0.01 and <0.05), respectively. In the cells treated with 5 and 10 μM of the compound, β-catenin was significantly increased 1.25- and 1.45-fold (p<0.001 and <0.01), respectively (Figure 4A, C). In 24 h of treatment, 10 μM of the compound caused induction of phosphorylated Akt 1.72-fold (p<0.05), compared to the control. In the cells treated with 5 and 10 μM of the compound, phosphorylated GSK3β was increased 2.08- and 1.97-fold (p<0.05 and <0.05), respectively. In the cells treated with 2.5 and 5 μM of the compound, β-catenin was significantly increased by 1.39- and 1.36-fold (p<0.05 and <0.05), respectively (Figure 4B, C). To validate the mechanistic control of PI3K/Akt pathway on β-catenin and SOX2, as well as test the effect of scoparone on the PI3K/Akt activation, a pharmacological experiment using specific PI3K inhibitors was performed. The cells were treated with scoparone in the presence or absence of specific PI3K inhibitors 50 μM LY294002 or 5 μM wortmannin and subjected to immunofluorescence analysis for β-catenin and SOX2 detections. Immunocytochemistry analysis showed that scoparone led to increased β-catenin and SOX2. Importantly, in the presence of 50 μM of LY294002 or 5 μM of wortmannin, β-catenin and SOX2 were found to be reduced (Figure 3A, B). These results indicate that scoparone has positive effects on the stemness of DPCs via the activation of PI3K/Akt-dependent regulation of β-catenin.

The effect of scoparone on the expression of Wnt/β-catenin related proteins. DPCs were treated with scoparone for 12 and 24 h. The expression levels of Akt, phosphorylated Akt, GSK3β, phosphorylated GSK3β, β-catenin and GAPDH in DPCs at 12 and 24 h were measured by Western blot (A, B). Relative protein levels were calculated by densitometry (C). Data represent mean±SEM (n=3). *p<0.05 compared to untreated control.
Discussion
DPCs play crucial roles in regulating the formation and development of hair follicles; in the mature follicle these cells could form the multipotent stem cell reservoir. The activity and function of DPCs has been demonstrated to be dependent on the stemness of these cells (4, 23, 24). Wnt/β-catenin signaling provides strong support to hair follicle morphogenesis and regeneration (5-7). In addition, Wnt/β-catenin is known to be an important coregulator of three master transcription factors (OCT4, NANOG and SOX2) that promote self-renewal of embryonic stem cells (8-10). It has been documented that the hair inducing property of DPCs gradually declines during culture (25, 26). The Akt axis has been shown to regulate the survival, growth and inductivity of DPCs (13). In addition, the inhibition of GSK3β using a specific GSK3β inhibitor in DPCs resulted in an activation of Wnt/β-catenin pathway and sustained the hair inductive potential (14). Knowing that Wnt/β-catenin is important for hair growth and has the ability to promote the three master pluripotency genes, it is likely that it may have a positive role in the stemness of DPCs.
In this study, we show that scoparone was able to preserve DPCs stemness during culture. Treatment of DPCs with scoparone significantly increased the cellular level of pluripotency related genes SOX2 and NANOG, while it had minimal effects on OCT4 (Figure 2B). Wnt/β-catenin signalling is important for maintaining stem cell properties (21, 26, 27). β-catenin regulates the expression of proteins facilitating stem cell functions by being a coactivator that, combined with T-cell factor/lymphoid enhancing factor (TCF/LEF), results in increased target gene expression (12). The GSK3β functions as β-catenin’s cellular level regulator by phosphorylating β-catenin, leading to its degradation. Active Akt inhibits GSK3β activity through phosphorylation at serine 9 (12). Subsequently, the activation of Akt results in increased cellular levels of β-catenin. The up-regulated β-catenin proteins translocate to the nucleus where they stimulate target genes (Figure 5). In this study, in order to elucidate the mechanism of action of scoparone, we found that scoparone exerted its stemness enhancing activity through the up-regulation of Akt-dependent inhibition of GSK3β, as the protein levels of active Akt (phosphorylated Akt) were increased, followed by the induction of inactive GSK3β (Figure 4C). It was shown that GSK3β is responsible for β-catenin degradation. We further found that the inhibitory effect of scoparone on GSK3β resulted in the increase of cellular levels of β-catenin, a stem cell controller (Figure 4C). To confirm this mechanism of action of scoparone via the PI3K/Akt pathway, DPCs were pretreated with PI3K inhibitors and the expression of β-catenin and SOX2 was analyzed by immunocytochemistry. The results demonstrated that scoparone alone induced the expression of β-catenin and SOX2, while their expression was found to be reduced in the presence of LY294002 or wortmannin (Figure 3A, B). These results suggest that the stemness sustaining effect of scoparone is due to the activation of the Akt/β-catenin pathway. They reveal a novel activity and mechanism of action of scoparone that could facilitate the development of hair enrichment approaches.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Schematic overview of the proposed mechanism of action of scoparone based on evidence from the present study, as well as previous literature. Scoparone has positive effects on the stem cell properties of human DPCs via Akt-dependent induction of cellular β-catenin. The increased β-catenin proteins enter the nucleus and stimulate the expression of target genes, including OCT4, NANOG and SOX2 which are transcription factors associated with pluripotency and self-renewal. Activation of Akt also directly phosphorylates OCT4, and as a transcription factor, phosphorylated OCT4 together with phosphorylated SOX2 regulates the transcription of NANOG.
Conclusion
This study provides new information for maintaining the stemness of dermal papilla by scoparone. The effect was mediated through Akt activation. Considering the stemness preserving properties, scoparone appears to be a promising natural agent that warrants further research for potential clinical applications.
Acknowledgements
This work was supported by the Pan Rajdhevee Group Public Co., Ltd. The authors express their gratitude to the Faculty of Pharmaceutical Sciences, Chulalongkorn University for providing laboratory resources.
Footnotes
This article is freely accessible online.
Authors’ Contributions
NS, PC and TT performed experiments and drafted the article. NS and TT performed experiments. BS isolated the tested compound. PC designed and supervised the research and wrote the article. PS supervised the research.
Conflicts of Interest
The Authors declare that there are no conflicts of interest.
- Received May 11, 2021.
- Revision received May 30, 2021.
- Accepted May 31, 2021.
- Copyright © 2021 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved