

Abstract
Aim: Identification of mutations and polymorphisms in the cytochrome b gene (Cyb) of mitochondrial DNA (mtDNA) in canine mast cell tumours and determinatiion of their association with the process of neoplastic transformation. Materials and Methods: The samples comprised tumour tissues and blood obtained from 34 dogs of various breeds. Mutations and polymorphisms in the Cyb gene were detected using amplification and sequencing methods. Results: Heteroplasmic mutations were detected at seven positions of mtDNA in 86% of the individuals. Blood and tumour heteroplasmy were recorded at five nucleotide positions of the Cyb gene, whereas tumour heteroplasmy was detected at two positions. Polymorphisms were detected at 14 Cyb gene positions in in the blood of 91% of dogs with mast cell tumours. Conclusion: The presence of numerous mutations and polymorphisms of Cyb in the blood and tumour tissues and the high frequency of heteroplasmy indicate their involvement in the process of neoplastic transformation in dogs.
- Cyb gene
- dog
- mast cell tumours
- mtDNA
- mutations
- mitochondrial DNA
Mast cell tumours (MCTs) are the most frequent canine skin tumours as they account for 7-21% of all diagnosed neoplastic lesions of this organ (1). Boxers are at the greatest risk of the disease, but Shar-Peis, Pugs, Labrador Retrievers, Golden Retrievers, Boston terriers and Pitt-Bull are also frequently mentioned in this context (2). However, in Boxers and Pugs, MCTs are commonly of histologically low or intermediate malignancy grade, whereas more aggressive course of MCTs has been observed in young Shar-Peis. The multifocal form of MCTs has been reported in Golden Retrievers (1, 2). MCTs can develop at any age, but the peak of their incidence is noted between 7.5 and 9 years of age (1). Most studies report no correlation between the sex of an animal and MCT occurrence as well as no gender predilection (1, 2). Due to their complex biological nature and unexplained aetiology, MCTs are still a subject of scientific research. The current knowledge about disturbances in the genetic material of MCTs is primarily focused on nuclear DNA (nDNA). The presence of internal tandem duplication (ITD) mutations in exon 11 of the proto-oncogene KIT proto-oncogene receptor tyrosine kinase (c-Kit) has been shown in 30% of this type of tumour (3, 4). Compared with mutations in exon 11, ITD mutations in exons 8 and 9 of c-Kit have been identified less frequently (5%) (5).
With the improvement of molecular biology techniques, involvement of the mitochondria in carcinogenesis is being increasingly suggested. Mitochondria possess their own genome characterised by higher variability than that of the nuclear genome. The frequency of mutations in mitochondrial DNA (mtDNA) is over 10-fold higher than their frequency in nDNA. Each cell contains from hundreds to thousands of mtDNA copies. When a cell/tissue contains both mutated and normal wild-type mtDNA, the condition is known as heteroplasmy. The presence of only one type of mtDNA (mutated or normal mtDNA) in the cell or tissue is homoplasmy. Mutations in mtDNA result in the synthesis of abnormal proteins of oxidative phosphorylation complexes, impaired cellular respiration, disorders in ATP production, and enhanced production of reactive oxygen species (6-8). Recent studies have documented the significant role of polymorphisms and mutations in mtDNA in the pathogenesis of many human diseases, including tumours (6-13). Somatic mtDNA mutations have also been detected in some canine tumours (14-18). Mutations and polymorphisms in genes coding for the respiratory chain subunits and in the displacement loop (D-loop) region have been found in mammary tumours as well as epithelial and mesenchymal tumours (16-18). Although there are relatively few literature data on the mitochondrial genomics of canine MCTs, mutations and polymorphisms have been detected in the D-loop region (19).
Mitochondrial diseases are associated with disorders in the function and biogenesis of respiratory chain components. Mutations in genes encoding respiratory chain subunit proteins induce changes in cell metabolism and may lead to neoplastic transformation (7, 8). Cytochrome b (CYB) is a respiratory chain protein. It is an integral membrane protein and a component of the complex III subunit. The protein is involved in regulation of oxidoreductase activity, metal ion transfer, and electron transport in the respiratory chain (20). Mutations in the gene encoding cytochrome b have been associated with some human tumour types, e.g. melanomas, mammary tumours, and bladder cancer (12, 21, 22). To our knowledge, there are no literature reports on the occurrence and role of Cyb gene mutations in MCTs, which prompts undertaking such investigations.
The aim of this study was to identify mutations and polymorphisms in the Cyb gene of mtDNA in mast cell tumours in dogs and determine their association with the process of neoplastic transformation.
Materials and Methods
The study involved 34 dogs of different breeds and crossbreed dogs with diagnosed MCTs. The characteristics of the dogs are provided in Table I. The dogs received neither chemotherapy nor hormonotherapy. Tumour tissue and blood samples were collected during surgery from all dogs. Blood samples of 1 ml each were taken from each dog using sterile test-tubes (Medlab Products, Raszyn, Poland) with di-potassium ethylene diaminetetra-acetic acid as an anticoagulant. All tumour tissues were sampled for histopathological and molecular analysis. Part of the tumour tissue for molecular analysis was transferred into sterile containers and kept in a refrigerator for further studies. For histopathology, part of the tissue was fixed in buffered formalin (pH 7.2), processed routinely, embedded in paraffin wax, sectioned into 4-μm fragments, stained with haematoxylin-eosin and toluidine blue, and evaluated under a light microscope for classification. Microscopic classification was performed in accordance with the WHO histological classification (23). The degree of histological malignancy was assessed using a 3-grade scale (GI, GII, GIII) according to Patnaik et al. (24) considering the depth of infiltration, cellular and nuclear pleomorphism, presence of giant cells, appearance of cytoplasmic granules, number of nucleoli, and number of mitotic figures. At the same time a two-tier scale (low grade and high grade) according to Kiupel et al. (25) was carried out, based on the morphology of the cell nucleus and the number of mitotic figures.
Data on dogs with mast cell tumours included in the study.
The main analyses were performed on DNA isolated from tumour tissues and blood samples. DNA was isolated with the use of an automated nucleic acid extraction system QIACube (Qiagen, Hilden, Germany). Isolation of DNA from both whole blood and tumour tissues was obtained in samples from seven dogs (dogs 1-7; Table I). For the remaining 27 dogs, it was only possible to isolate DNA from blood only (dogs 8-34; Table I). The DNA samples were assessed quantitatively and qualitatively by electrophoretic separation in 2% agarose gel and spectrophotometrically by measurements of sample absorbance in a Bio Photometer (Eppendorf, Hamburg, Germany). Amplification of the Cyb gene (724-base pair fragment) was performed using a polymerase chain reaction (PCR) technique in a T100 Thermal Cycler (Bio-Rad, Wroclaw, Poland). Primer sequences used in the analysis, encompassing a mtDNA fragment (Cyb F: 5’ GCACGCAAATGGCGCTTCCA, Cyb R: 5’GCATTGGCTAAGGGGCCGGA), were based on already published data (23). The annealing temperature for all the analysed gene fragments was 58.5°C. Amplicons were sequenced using a Big Dye Terminator Cycle Sequencing kit (Applied Biosystems, Foster City, CA, USA) in Gene Amp PCR system 9700 (Applied Biosystems). The samples were subsequently purified on Centri Sep columns according to the manufacturer's protocol or precipitated with ethanol and sodium acetate according to the protocol provided by the Big Dye kit manufacturer. Extension products were separated on an ABI 377 automated sequencer (Applied Biosystems). Nucleotide sequences were analysed using bioinformatics programs to determine the presence or absence of point mutations and polymorphisms at the analysed loci in tissues (DNA Baser Sequence Assembler v 3.2). The Cyb nucleotide sequences were compared with the reference sequence (Gen Bank: U96639) (26, 27).
Somatic mutations in cytochrome b (Cyb) gene sequence in blood and tumour tissue in cases of canine mast cell tumour (dogs 1-7).
All conducted procedures in the study group were approved by the II Local Ethical Commission for Animal Experiments in Lublin, Poland (Resolution number 80/2014). The samples collected for the study did not required dog owners' consent.

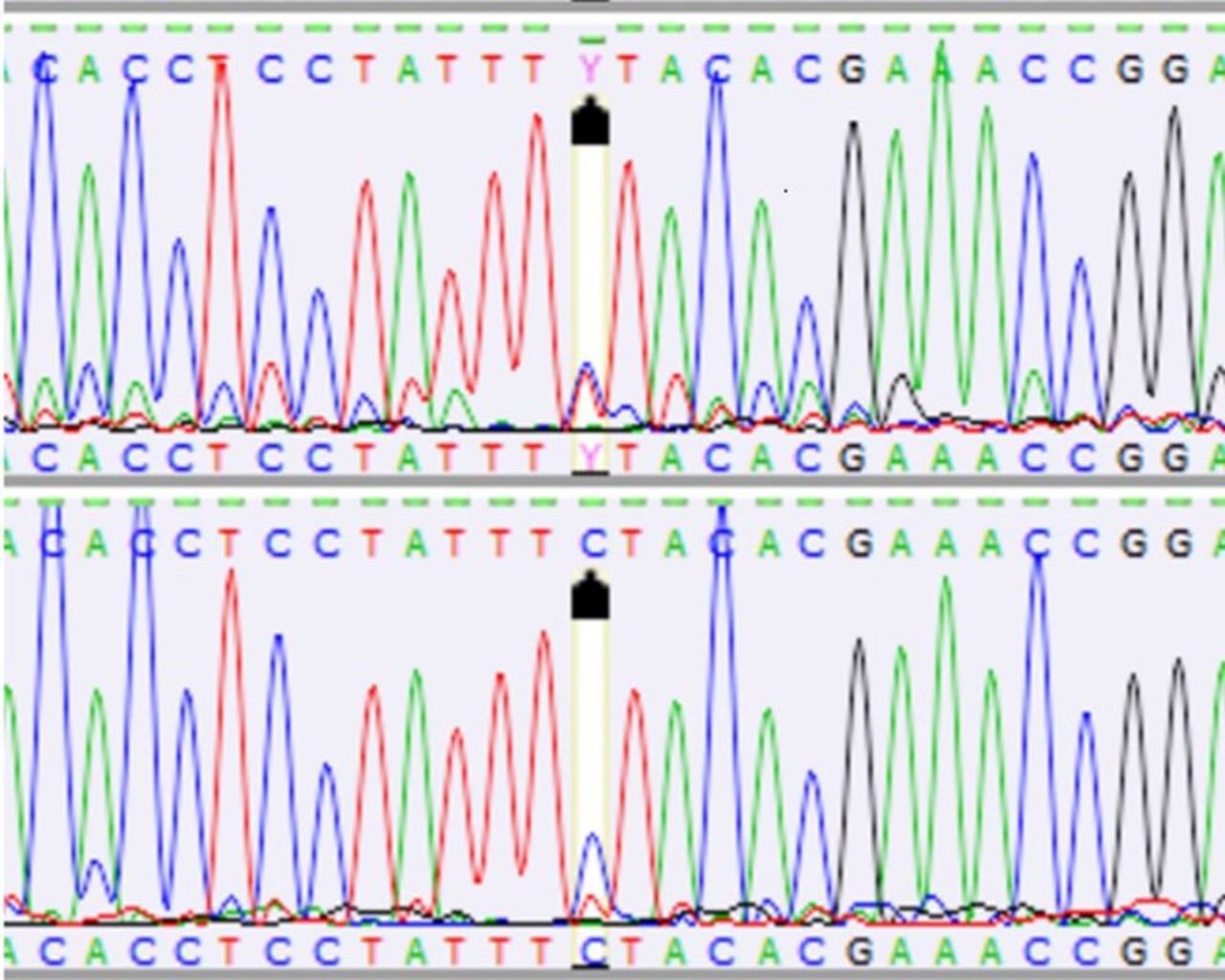
Tumour heteroplasmy (m.14780C/T, upper panel) and blood (lower panel) sequence for mitochondrial cytochrome b (Cyb) in position m.14780 in canine mast cell tumour (dog number 4).

{kind=link}
{kind=link}
Tumour (upper) and blood (lower) mutation (G>A) of mitochondrial cytochrome b (Cyb) in position m.14474 in a canine mast cell tumour (dog number 2).
Results
Somatic mutations or polymorphisms in the Cyb gene were detected in all the dogs with MCTs. The analysis revealed a relatively high level of molecular genetic variation between the Cyb gene sequence in the tumour tissue and blood of individual animals (dogs 1-7) (Table II). There were mutations in 12 nucleotide positions of the Cyb gene in the blood/tumour tissue of the examined animals (Table II). All the changes were substitution mutations. A majority of the mutations identified were heteroplasmic. Heteroplasmic mutations were detected in seven out of the 12 nucleotide positions (m.14527T/C, m.14634C/T, m.14674C/T, m.14780C/T (Figure 1), m.14972A/C, m.15010C/T, m.15032A/C) in six out of seven of the individuals. In five out of seven of the individuals, heteroplasmy was found in at least one Cyb gene nucleotide position both in the tumour tissue and in blood (Table II). Blood and tumour heteroplasmy was recorded in five nucleotide positions of the Cyb gene (m.14634C/T, m.14674C/T, m.14780C/T, m.15010C/T, m.15032A/C), whereas tumour heteroplasmy was detected in positions m.14527T/C and m.14972A/C. Homoplasmic mutations in mtDNA were detected in five out of the 12 positions in the analysed fragment (m.G14474A (Figure 2), m.G14504C, m.G14671A, m.T14930C, m.T14977C) (Table II). Mutations in mtDNA positions m.14780, m.14634, and m.14674 exhibited the highest frequency (five, four, and four out of the seven dogs, respectively) (Table II). There was an inverse correlation between the number of mutations and tumour grade in the classification according to Kiupel et al. (15). In high-grade MCTs, there were two mutations and only one of the six substitutions was non-synonymous (Table II). In the case of low-grade MCTs, there were from four to five mutations, with one to three non-synonymous substitutions (Table II). The age of the dogs was not correlated with the number of mutations (Table II). Five out of the 12 mutations detected in the canine MCTs resulted in a change in amino acid (Table II). The heteroplasmic Cyb gene mutation m.C14634C/T caused a serine-to-phenylalanine substitution (p.S151F) in four dogs. Mutation m.A15032A/C resulted in a substitution of isoleucine by leucine (p.I284L) in two dogs. Another three non-synonymous mutations were found in single individuals. The heteroplasmic mutation (A→A/C) detected in nucleotide position m.14972 in tumour induced a threonine-to-proline substitution (p.T264P). A guanine-to-alanine substitution was noted in position m.14474 of mtDNA in blood; it resulted in a substitution of valine by methionine (p.V98M). Similarly, a blood non-synonymous mutation (G→C) in position m.14504 of mtDNA led to a substitution of valine by leucine (p.V108L) in the polypeptide chain. Five out of seven of the examined dogs exhibited from one to three non-synonymous mutations. The presence of only synonymous mutations was detected in two individuals (Table II).
Polymorphisms in the mitochondrial cytochrome b (Cyb) gene in blood in cases of canine mast cell tumour (dogs 1-34).
Polymorphisms in the blood of dogs with MCTs, in comparison with the canine reference sequence, were detected in 14 Cyb gene positions in 31/34 (91%) of the examined dogs (Table III). Four of them (m.G14474A, m.C14634T, m.A14972C, m.A15032C) induced amino acid substitutions in the protein (p.V98M, p.S151F, p.T264P, and p.I284L, respectively) (Table III). The highest frequency, i.e. in 20/34 (59%) of the dogs, was noted for polymorphisms m.C14634C/T and m.C14674C/T. In 33/34 (97%) of the dogs with MCTs, there were from one to seven polymorphisms in the Cyb gene (Table III). In the case of polymorphisms, heteroplasmy was detected in one to four nucleotide positions of the Cyb gene in 31/34 (91%) of the dogs (Table III).
Sex-specific polymorphisms were also observed. Polymorphism in position m.14474 was detected in three females (no. 16, 21, 26) and in five females (no. 1, 9, 25, 28, 32) in position m.15010. In turn, polymorphism in m.14972 was found only in three male dogs (2, 31, 33) (Table III). Tumour grade-specific polymorphisms according to the classification proposed by Kiupel et al. (15) were also noted. Substitutions in positions m.15010 and m.15032 were found in low-grade MCTs, but were not detected in high-grade tumours (Tables I and III). The polymorphisms in positions m.14634, m.14674, and m.14977 were more frequent in the case of low-grade than in the high-grade tumours. The number of polymorphisms ranged from 1 to 7 in low-grade MCTs and from 0 to 5 in high-grade tumours. In our study, we did not observe any relationship between the age and breed of the dogs with polymorphisms. The mean number of polymorphisms per individual was higher in the group with low-grade tumours (median=3.5) than in the high-grade MCTs (median=3.0).
Discussion
Previous investigations on the aetiology of MCTs associated them with changes in nDNA. The only published report of the mitochondrial genomics of MCTs was focused on the D-loop, which controls replication and transcription of mtDNA. Somatic mutations in seven positions of the D-loop nucleotide sequences were detected in 47% of dogs with MCTs, while polymorphisms were identified in 94% of the dogs (19). The previous and present research confirmed the relatively high molecular genetic variation in the mitochondrial genome in MCTs, which may be related to tumorigenesis. The mutations were detected in both low-and high-grade tumours. However, an inverse correlation was noted between the number of mutations and the tumour grade according to Kiupel et al.'s classification (25) (Table II). The c-Kit nDNA mutation in exon 11 described so far in MCTs was associated only with tumours with a higher malignancy grade and worse prognosis; it was not observed in tumours with a more benign character (3, 4). In turn, in the case of the Cyb gene, a greater number of mutations were found in low-grade tumours. These mutations probably appear in the initial stage of cancer and might even initiate the carcinogenic process. This is supported by the occurrence of such mtDNA mutations in the human population also in dysplastic changes regarded as pre-cancerous states (7, 8). Mutations of genes in mtDNA encoding respiratory chain proteins, including cytochrome b, induce disturbances in electron transport and promote generation of reactive oxygen species. It should be noted that the later leads to the initiation and development of cancer and damage to the nuclear genome (7, 8). Whether there are correlations between mutations in the nuclear and mitochondrial genomes should be elucidated by further studies focused on both genomes.
Most changes within the mtDNA nucleotide sequences of neoplastic cells are of homoplasmic nature, i.e. all DNA molecules in the cell are the same (6, 8). This was not confirmed in our studies, however, since the majority of the mutations were heteroplasmic (Table II). This may be associated with the initial stage of carcinogenesis. At the beginning of the neoplastic process, cells may contain mutant and wild-type mtDNA copies. Only subsequent divisions of mutant cells can lead to homoplasmy. The presence of mutations in the Cyb gene in low-grade tumours may also indicate an early stage of carcinogenesis. However, this needs further study. In earlier reports on mitochondrial genomics in canine tumours, the phenomenon of heteroplasmy was detected in single cases (14-18). One report of a mitochondrial defect in the Cyb gene in canine tumours described heteroplasmic mutations in sebaceous gland epithelioma and centroblastic lymphoma. The mutation in the tumour tissue was found in position A587A/C. It was also found in position 738A/C in the lymphoma (16). In a previous study of the D-loop in MCTs, heteroplasmy was detected in two cases. In one individual, heteroplasmy was present in the blood and tumours in position m.C15815C/T; in another dog, heteroplasmic changes were found only in tumour tissue in positions m.C15912T/C and m.C16025T/C (19). Coller et al. (28) suggest that homoplasmy is the result of random segregation of mitochondria during division. During consecutive divisions, heteroplasmy may persist or homoplasmy may occur through genetic drift (7), which results in elimination of rare variants of mtDNA. The mutual proportions of mutant and wild-type mtDNA can vary during division, leading to changes in the cellular phenotype. Phenotypic expression has clinical importance and exerts an impact on the manifestation of disease symptoms (29). MCTs are characterised by a varied clinical course. They take the form of small, demarcated, single or multiple tumours and infiltrate surrounding tissues (30). Can the clinical picture be associated with the phenomenon of heteroplasmy? It is difficult to provide an answer to this question at this stage of the research; however, the high frequency of this phenomenon in the Cyb gene in MCTs, compared with other tumour types, is puzzling (16). Five out of seven individuals exhibited heteroplasmy in both their tumour tissue and blood (Table II). In the case of malignant tumours such as MCT, the presence of mutations in blood may be related to the transport of tumour cells with the mutant variant of the gene into the bloodstream through tumour blood vessels (7). The occurrence of mutations in blood can be the basis for further investigations focused on identification of prognostic markers and tumour progression.
Due to their influence on the structure of proteins of the oxidative phosphorylation chain, polymorphisms alter the course of cell processes and modify cell susceptibility to neoplastic transformation (6, 11). So far, intensive research on the role of polymorphisms in oncogenesis has been carried out on breast cancer in women (9-12, 31, 32). Correlations have been found between polymorphism in m.G10398A and m.T10400C of mitochondrial NADH-dehydrogenase subunit 3 (MT-ND3) and a high risk of breast cancer development (11, 32). Investigation conducted by Igman et al. (31), helped to establish the patterns of 63 polymorphisms present in patients with breast cancer. In turn, Grzybowska-Szatkowska et al. (9) detected the greatest number of polymorphisms in the sequence of the mitochondrially encoded NADH: ubiquinone oxidoreductase core subunit 2 gene (MT-ND2).
It has been found that polymorphisms can affect the function of mitochondria, especially if they lie on conservative domains of the mitochondrial proteins. These changes can promote the selective prevalence of mutant mtDNA over the wild-type mtDNA, which may be involved in the process of carcinogenesis (10). The negative effect of polymorphisms on protein function has been confirmed in malignant canine tumours (16, 18). Ślaska et al. have shown harmful effects of changes in polypeptides in four positions: T193N, V98M, V118M, and H196P on the structure and function of proteins (14). This promotes disorders in electron transport and the energy level in cells.
In this study, polymorphisms in blood were detected in all individuals (Table III). The highest frequency of polymorphisms was found in two positions: m. C14634C/T and m.C14674C/T, which may predispose to the development of MCT.
As in the case of mutations, the high frequency of heteroplasmy, which was noted in as many as 91% (31/34) of the dogs in one to four Cyb gene nucleotide positions, is surprising (Table III). In the initial stage of carcinogenesis, cells contain mutant and wild-type mtDNA copies. The use of polymorphisms as tumour markers could be a very useful tool in the diagnostics of MCTs. To our knowledge for the first time, sex-specific polymorphisms were detected. The polymorphisms in positions m.14474 and m.15010 were only found in females, whereas the polymorphism in m.14972 was present only in males (Table III). Literature data demonstrate no relationship between MCT development and the animal's sex (1, 2). Molecular analysis here revealed sex-specific polymorphisms that may be associated with the risk of MCT development in females and males. However, further investigations with a greater number of individuals are required. Furthermore, the polymorphisms in position m.15010C/T m.15032A were detected only in low-grade MCTs (Table III). Could the presented polymorphisms be prognostic markers? The answer to this question requires further research considering the course of the tumour disease and survival rates. To date, a correlation of polymorphisms in position m.8242C/T of the ATP synthase F0 subunit 6 gene (Atp6) and m.7383T/C of the cytochrome c oxidase subunit II gene (Cox2) with a large body size and older age has been noted only in female dogs with mammary tumours. In our previous and available studies on mitochondrial genomics in dogs, there was no relation between polymorphism and breed susceptibility (14-19). However, interesting results of molecular analyses of nuclear genes were obtained in Golden Retriever dogs: polymorphisms in the G protein subunit alpha i2 gene (Gnai2) and hyaluronidase genes have been associated with the risk of MCT development (33).
Conclusion
The present investigations demonstrated high molecular genetic variation in the cytochrome b-encoding gene in MCTs. There are many scientific theories regarding cancer transformation. One of them assumes that mutations are a result of tumour diseases (6, 10). However, the vast majority of researchers support the view that mutations and polymorphisms promote tumour development. This hypothesis seems to be confirmed in the present study. The presence of numerous mutations and polymorphisms in blood and tumour tissues and the high frequency of heteroplasmy indicate their involvement in the process of neoplastic transformation. It cannot be excluded that these mutations and polymorphisms are directly associated with the development of mast cell tumours in dogs and may be used for diagnosis of tumours. As the results come from a limited number of cases, further studies on the significance of mutations and their association with neoplastic transformation as well as the biological behaviour of canine MCTs are required.
Footnotes
This article is freely accessible online.
Conflicts of Interest
The Authors declare that they have no competing interests in regard to this study.
- Received September 5, 2018.
- Revision received October 8, 2018.
- Accepted October 12, 2018.
- Copyright© 2019, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved