

Abstract
Background: Tumor-infiltrating lymphocytes (TILs) and tertiary lymphoid structures (TLSs) are prognostic markers in triple-negative breast cancer (TNBC). Our study analyzed the relationship between cluster of differentiation (CD)11c-positive dendritic cells (DCs) and TILs and TLSs to elucidate mechanisms of TIL influx. Materials and Methods: Immunohistochemical staining for CD4, CD8, and CD11c in tissue microarrays from 681 patients with TNBC was performed. The proportions of TILs and TLSs were reviewed. Two additional TNBC gene expression datasets were used. Results: CD11c expression showed a significantly positive correlation with the level of TILs and the number of CD4+ and CD8+ T-cells, as well as an abundance of TLSs. CD11C gene expression was also significantly correlated with expression of CD4, CD8, and genes related to TLSs in both datasets. Conclusion: We demonstrated a strong correlation of CD11c expression, which represents DCs, with TILs and TLSs in TNBC. Further investigation is warranted to identify therapeutic modalities that facilitate recruitment and activation of DCs.
- Breast cancer
- dendritic cell
- CD11c
- tumor-infiltrating lymphocyte
Although triple-negative breast cancer (TNBC) shows aggressive clinical features, patients with higher levels of tumor-infiltrating lymphocytes (TILs) and tertiary lymphoid structures (TLSs) in the tumor microenvironment have better clinical outcomes (1, 2). In order to explore mechanisms of TIL influx in the TNBC microenvironment, we previously analyzed the relationship between the expression of endoplasmic reticulum stress (ERS) molecules protein kinase R-like endoplasmic reticulum kinase (PERK), phosphorylated eukaryotic initiation factor 2α (p-eIF2α), and X-box binding protein (XBP1), and danger-associated molecular patterns (DAMPs), including high mobility group B1 and N1 (HMGB1 and HMGN1) proteins in tumor cells, and the level of TILs in patients with TNBC. Higher expression of ERS molecules in cancer cells was associated with higher cytoplasmic expression of HMGB1 and HMGN1, which are nuclear proteins and known to be released in the context of ERS and higher TIL levels in TNBC (3, 4). We also confirmed that HMGB1/HMGN1 were released into the extracellular space from several TNBC cell lines upon pharmacological ERS, which suggests that ERS triggers the release of HMG proteins following TIL influx in TNBC (3).
Tumor cells, dendritic cells (DCs; representative antigen-presenting cells in adaptive immunity), and cytotoxic T-cells are organizationally coordinated during antitumor immune responses. DAMPs, including HMGB1, were originally found to be released by damaged tissue but have also been discovered to be released by different tumor types (5). It is suggested that HMGB1 secretion also has roles in cancer (5, 6). HMGB1 binds to toll-like receptor 4 (TLR4) on DCs, which exerts an important role in subsequent T-cell activation (7, 8).
The association between DAMPs and TILs, however, has not been thoroughly investigated clinically. Murakami et al. reported that high expression of major histocompatibility complex class I polypeptide-related sequence A and B (MICA/B), a DAMP component that is specifically induced by chemotherapy, and irradiation, was associated with a low ratio of regulatory T-cell (Tregs) to TILs in patients with pancreatic cancer who underwent neoadjuvant chemoradiotherapy (9). The control group, who did not receive neoadjuvant therapy, showed no such significant association (9). In colon cancer, HMGB1 expression in both the nucleus and cytoplasm was reported to be inversely associated with the amount of both cluster of differentiation (CD)3+ and CD45RO+ T-cells infiltrating the tumor stroma (10). In lung adenocarcinoma, high densities of infiltrating DCs have been reported to be associated with cytoplasmic HMGB1 expression and with greater numbers of TILs (11).
In order to determine the relationship between DCs and T-cells, our study analyzed CD11c, CD8 and CD4 expression and the level of TILs and presence of TLSs in surgical specimens of TNBC without previous systemic therapy. The expression of genes related to DCs, T-cells, and TLSs was investigated using The Cancer Genome Analysis (TCGA) data (http://www.gdac.broadinstitue.org), as well as biopsy specimens from patients with TNBC who received neoadjuvant chemotherapy.
Materials and Methods
Patients and specimens. We analyzed tissue microarrays from 681 previously described preoperative chemotherapy-and radiotherapy-naïve patients with TNBC who underwent surgery for primary breast cancer between 2004 and 2010 at the Asan Medical Center, Seoul, Korea (12). All patients underwent adjuvant systemic treatment, and 471 patients (69.2%) without lymph node metastasis were treated with four cycles of adjuvant anthracycline and cyclophosphamide (AC) (adriamycin 60 mg/m2 and cyclophosphamide 600 mg/m2). The remaining 210 patients with lymph node metastasis were treated with four cycles of AC followed by either four cycles of paclitaxel (175 mg/m2) or four cycles of docetaxel (75 mg/m2). Exemption from informed consent, after de-identification of information, was approved by the Institutional Review Board of the Asan Medical Center. This study was performed in accordance with the Declaration of Helsinki.
The hematoxylin and eosin-stained slides were reviewed by two pathologists (H.J.L. and G.G.). The histological grade of breast cancer was evaluated according to the modified Bloom–Richardson grading system (13). The slides were evaluated for stromal TIL levels using full sections in 10% increments (if TIL was less than 10%, one out of 0, 1, or 5% was chosen), and the proportion of TLSs in the adjacent area of the invasive tumor was graded as: None: No TLS formation in the area adjacent to the tumor; little: TLSs occupying an area of less than 10% of the circumference of the tumor; moderate: 10% to 50% coverage; and abundant: greater than 50% coverage (2, 14).
Immunohistochemical analysis. Formalin-fixed, paraffin-embedded tissue microarray sections were stained with an automatic immunohistochemical staining device (Benchmark XT; Ventana Medical Systems, Tucson, AZ, USA). Antibodies for CD11c (1:1000; Abcam, Cambridge, UK), CD4 (1:4; Ventana Medical System), and CD8 (1.200; Dako, Glostrup, Denmark) were used. CD11c-immunostained slides were scanned using a Vectra 3.0 microscope system (PerkinElmer, Waltham, MA, USA). The entire slide (×4) was scanned first and then we selected and took images (×20) from tissue microarray cores. Integrated optical density was obtained from the sum of multiplying the average intensity of 3,3’-diaminobenzidine (DAB) staining and the number of pixels of DAB-stained areas.
The immunostained tissue microarray slides for CD4 and CD8 were scanned using a digital microscope scanner (Pannoramic 250 FLASH; 3DHISTECH Ltd., Budapest, Hungary) as previously described (12). The number of CD4+ and CD8+ cells was determined using the NuclearQuant module of the Pannoramic Viewer 1.15.2 (3DHISTECH Ltd).
Gene expression analysis. Two sets of TNBC gene expression data were used to analyze the relationship of CD11c positivity with T-cells and TLSs. The first set consisted of 55 TNBC biopsy cases prior to neoadjuvant chemotherapy (15). Total RNA (100 ng) extracted from formalin-fixed paraffin-embedded biopsy specimens was assayed on an nCounter Digital Analyzer with nCounter GX Human Immunology V2 kit (NanoString, Seattle, WA, USA) (16). Data were normalized by scaling with the geometric mean of built-in control gene probes for each sample and were log2-transformed. The second set consisted of 189 TNBC cases from TCGA invasive breast cancer data downloaded from the Broad GDAC Firehose website (https://gdac.broadinstitute.org). the mRNA expression (RNA seqV2 RSEM) of eight immune-related genes was analyzed: CD4, CD8A, CD8B, C-C motif chemokine ligand 21 (CCL21), C-X-C motif chemokine ligand 12 (CXCL12), CXCL13, lymphotoxin beta (LTB), and CD11c (17).
Statistical analyses. All statistical analyses were carried out using SPSS version 20 (IBM Corp., Armonk, NY, USA). Spearman correlation, chi-square test, linear by linear test, log-rank test, and Cox proportional hazards regression model were used, as appropriate. All tests were two-sided with a 95% significance level.
Results
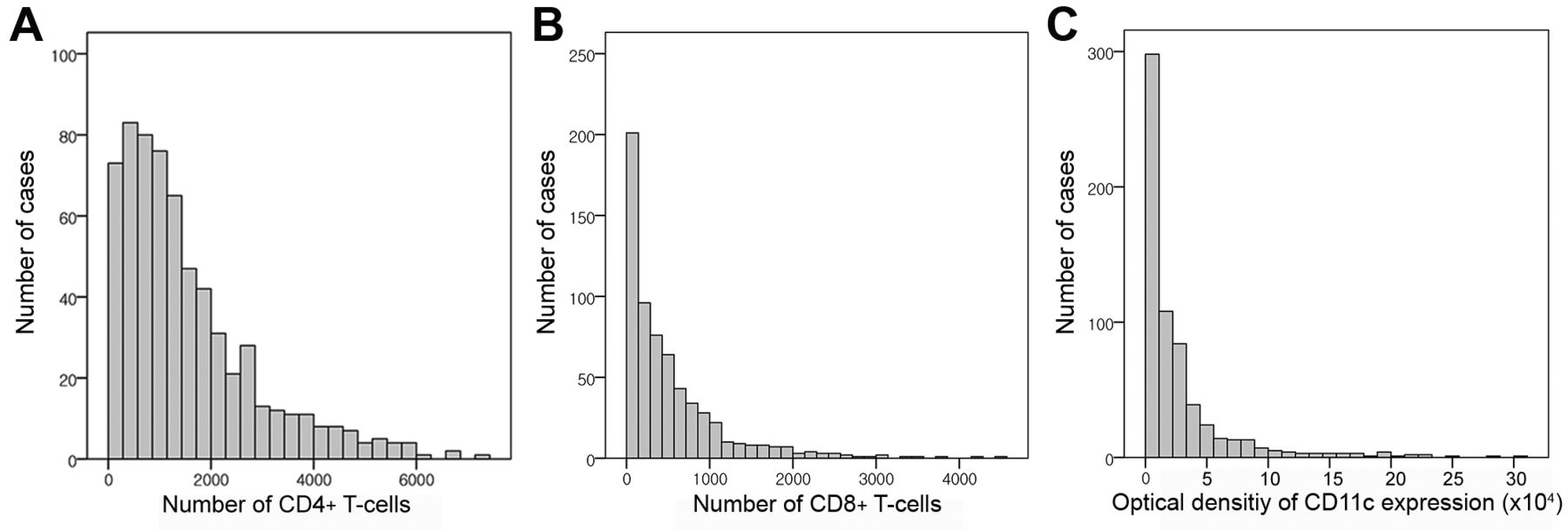
T-Cell and CD11c distribution. On microscopic examination, CD11c positivity was predominantly observed in areas with larger numbers of TILs and TLSs (Figure 1). Counting DCs is difficult due to their extended dendrites, therefore the integrated optical density for CD11c positivity was used to quantify DCs in the TNBC microenvironment (18). The distributions of CD4+ and CD8+ T-cells and CD11c integrated optical density were skewed to the left (Figure 2). The median CD4+ and CD8+ T-cell counts per tissue microarray core were 1171 (range=6-7,161) and 335 (range=1-4,443), respectively. The median integrated optical density for CD11c positivity was 12,703 (range=12.7-309,903.6).
Comparison of clinicopathological variables according to CD11c expression in patients with TNBC. The integrated optical density of CD11c positivity was significantly positively correlated with TIL levels (Spearman's rho=0.483, p<0.001) and number of CD4+ (rho=0.591, p<0.001) and CD8+ T-cells (rho=0.515, p<0.001, Figure 3). In order to analyze clinicopathological correlation, tumors were divided into two groups based on the median integrated optical density for CD11c positivity. Tumors with higher CD11c expression had significantly higher histological grades (p=0.011) and more abundant surrounding TLSs (p<0.001, Table I). Pathological T stage and presence of lymph node metastasis and lymphovascular invasion did not vary by CD11c expression level.

Histological features and distribution of immune cells of triple-negative breast cancer with abundant tumor-infiltrating lymphocytes (TILs) and tertiary lymphoid structure (TLS) formation. A: High-grade tumor cells with TILs and TLS (upper left) are shown (H&E-stained, ×100). B-D: Distribution of cluster of differentiation (CD)4+ T-cells (B), CD8+ T-cells (C) and dendritic cells (D) throughout TILs and TLS are highlighted by immunohistochemistry (×100). E, F: Dendritic cells showing CD11c expression are predominantly located in areas with large numbers of TILs (E, ×200) and TLS (F, ×200).

Distribution of cluster of differentiation (CD)4+ and CD8+ T-cells and CD11c positivity. The number of CD4+ T-cells (A) and CD8+ T-cells (B) in tissue microarray cores were automatically counted and are shown in a histogram. The mean number of CD4+ T-cells was larger than that of CD8+ T-cells, and both their distributions are left-skewed. The optical density of CD11c expression (C) measured instead of counting dendritic cells suggests that the distribution of dendritic cells is also skewed to the left.
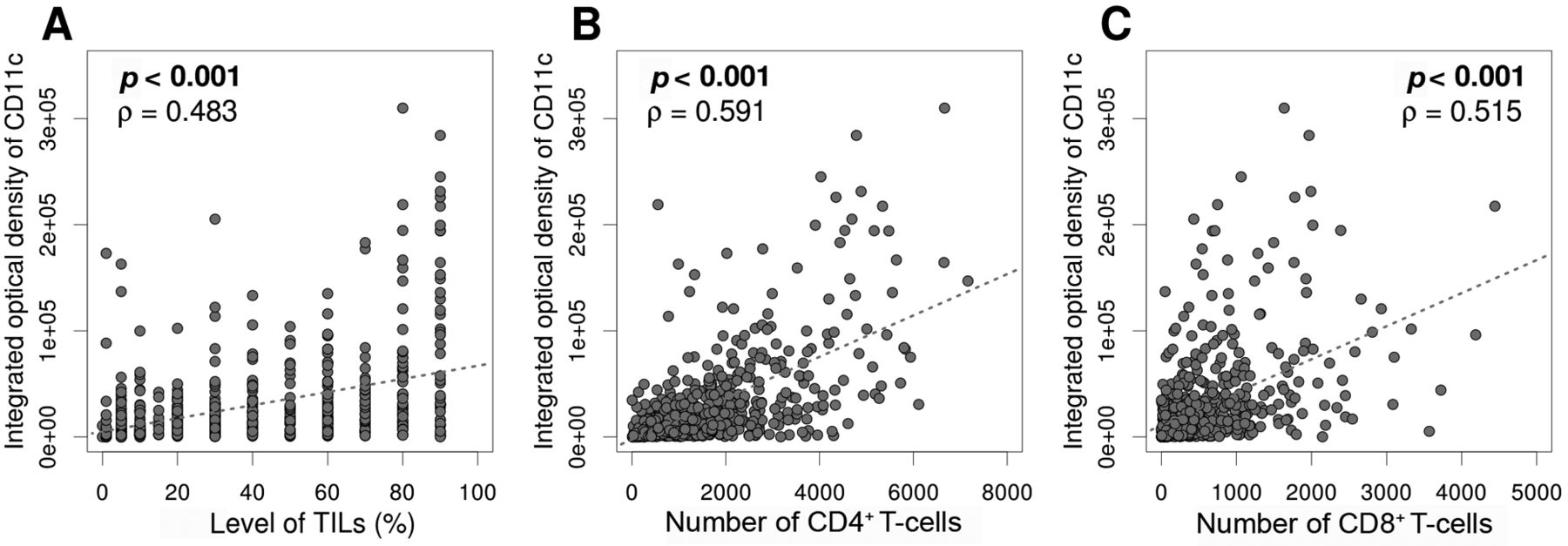
Correlation between cluster of differentiation (CD)11c expression and the abundance of tumor-infiltrating lymphocytes (TILs) (A) and CD4+ (B) and CD8+ (C) T-cells. Integrated optical density of CD11c positivity was significantly positively correlated with the abundance of TILs, the number of CD4+ T-cells, and that of CD8+ T-cells.
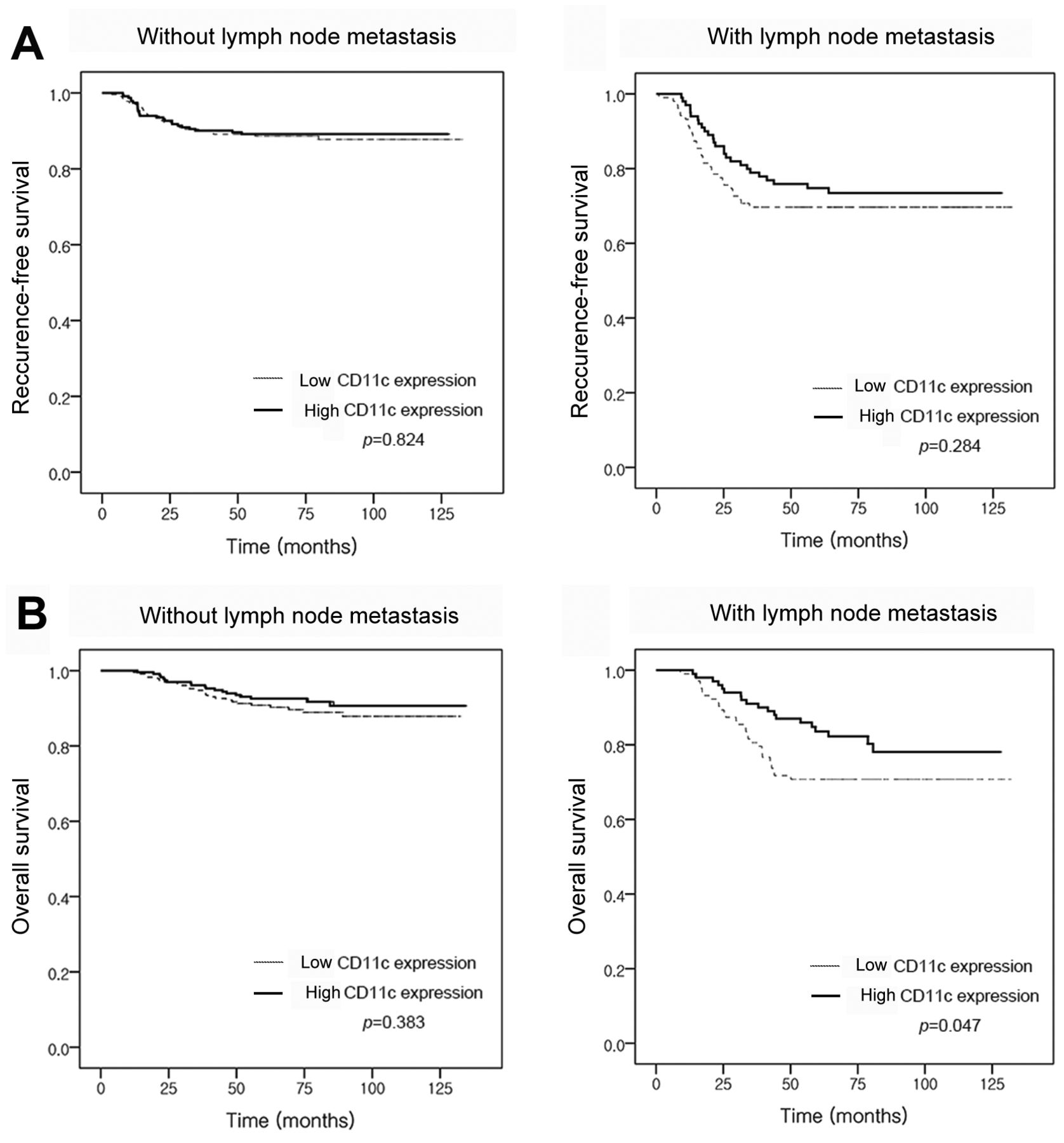
Prognostic significance of CD11c expression in patients with TNBC. Chemotherapy regimens and TNBC prognosis differed according to lymph node metastasis status; survival analyses were, therefore, performed on patients according to lymph node metastasis. Among patients without lymph node metastasis, there was no difference in survival between those with high or low CD11c expression (recurrence-free survival, p=0.824; overall survival, p=0.383, Figure 4). On the other hand, in the group with lymph node metastasis, high CD11c expression showed a trend for being associated with better recurrence-free survival (p=0.284) and significantly better overall survival (p=0.047) compared to low CD11c expression. However, in multivariate analysis (including pathological T stage, lymphovascular invasion, TIL level and CD11c expression), CD11c expression was not independently associated with better overall survival in patients with lymph node metastasis [pathologicaI T stage, hazard ratio (HR)=1.147, 95% confidence interval (CI)=0.720-1.827, p=0.565; lymphovascular invasion, HR=2.006, 95% CI=1.080-3.726, p=0.028; TIL, HR=0.978, 95% CI=0.965-0.991, p=0.001; CD11c, HR=1.0, 95% CI=1.0-1.0, p=0.394].
Clinicopathological correlation of CD11c positivity in consecutive triple-negative breast cancer cohorts.
Gene expression analysis with two different TNBC cohorts. In order to confirm the close relationship of CD11c with T-cells and TLSs, we analyzed the associated gene expression in two different TNBC cohorts. The first set consisted of 55 TNBC cases with NanoString-based gene expression data, covering genes associated with TLSs (CXCL12 and CXCL13) (15, 19). The second set consisted of RNA sequencing data of 189 TNBC cases from TCGA breast cancer data, which included genes associated with TLSs (CCL21, CXCL12, CXCL13, and LTB). In the first set, CD11c gene expression was positively associated with CD4, CD8A, CD8B, CXCL12, and CXCL13 expression (rho >0.5, p<0.001, Table II). Likewise, CD11C gene expression was also positively correlated with CD4, CD8, and other genes related to TLSs, including LTB in the second dataset (rho>0.4, p<0.001, Table III).
Discussion
CD11c, a member of the integrin β2 adhesion molecule family, is highly expressed in monocytic and myeloid DCs, which are representative antigen-presenting cells (20). Antigen-presenting cells are important for the induction of antitumor immunity by priming naïve T-cells and leading them to activate and differentiate into effector cells (21). Chemokines, including CCL3, CCL4, CCR5L, CCL17, and CCR4 (which are responsible for recruiting naïve cytotoxic T-lymphocytes) are secreted by DCs. In the present study, we demonstrated that CD11c positivity was significantly correlated with CD4+ and CD8+ T-cells counts and TIL levels in surgical specimens of patients with TNBC. In addition, we analyzed the distribution of immunohistochemical CD11c expression and found that CD11c+ cells were located in areas with high numbers of TILs. Therefore, we confirm there to be a close association between CD11c+ DCs and T-cells in TNBC.
Correlation between genes associated with T-cells and tertiary lymphoid structures and CD11C from patients with triple-negative breast cancer using NanoString-based gene expression data (n=55).
We also identified a positive correlation between CD11c positivity and the presence of TLSs in the TNBC microenvironment. The positive relationship of CD11C expression with CD4, CD8, and genes related to TLSs was also confirmed in gene-expression analysis using NanoString-based gene expression data and TCGA breast cancer RNA sequencing data. In the chronic inflammatory environment (including in the context of tumor growth), lymphoid neogenesis is induced, and TLSs structurally and functionally resembling secondary lymphoid organs are formed (22). B-Cell zones with germinal centers and distinct T-cell-rich areas with DCs can be found in TLSs. We previously demonstrated that TLSs in breast cancer are usually present in areas surrounding invasive tumors and are closely associated with terminal duct lobular unit and ductal carcinoma in situ (2, 23, 24). Inflammatory cytokines and chemokines, such as LTa, LTa1b2, CXCL13, CCL21, CCL19, interleukin (IL)17, IL22, IL23, IL7, and IL27 produced by various cell types (including DCs and macrophages in the tumors) are involved in initiation, formation, and maintenance of TLSs (25). High endothelial venules and lymphatic vessels in TLSs enable recruitment of peripheral immune cells into the tumor microenvironment, and accordingly activated immune cells generate local antitumor immunity. Thus, the presence and abundance of TLSs in the tumor are associated with the number of TILs and beneficial clinical outcomes. We previously reported that TLSs are abundant in TNBC and that the abundance of TLSs is associated with the TIL level (2). Patients with moderate or abundant TLSs had better survival than those with few or no TLSs at higher pathological stage regarding tumor, lymph node, and distant metastasis. Moreover, the presence of TLSs at a site of breast cancer metastasis exhibited was significantly correlated with the abundance of TILs in metastatic sites and better overall survival (unpublished data).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Kaplan–Meier survival analysis of recurrence-free (A) and overall (B) survival for patients with triple-negative breast cancer according to cluster of differentiation (CD)11c expression and lymph node status. In patients without lymph node metastasis, no difference recurrence-free or overall survival according to CD11c expression was observed (left panel). On the other hand, high expression of CD11c was related to better overall survival in patients with lymph node metastasis (right panel).
We showed that, in the context of lymph node metastasis, patients with high CD11c expression had a better overall survival. However, in multivariate analysis, CD11c expression was not an independent prognostic factor for better overall survival. This result might be caused by the strong association between CD11c positivity and TIL level, which is a significantly important prognostic factor in TNBC.
Correlation between genes associated with T-cells and tertiary lymphoid structures and CD11C from patients with triple-negative breast cancer [The Cancer Genome Atlas data (n=189)].
CD11c can also be expressed by activated macrophages, natural killer cells, and T-cells (26-29). Better clinical outcomes in colorectal cancer, gastric cancer, and malignant melanoma have been associated with CD208 (dendritic cell-lysosomal-associated membrane glycoprotein, DC-LAMP) expressed on mature DCs located in T-cell areas of lymphoid tissues, as well as CD86 (B7.2, a ligand of CD28/CTLA-4) expressing mature DCs (30-32). Another study showed that tumor-infiltrating CD1a+ immature DCs were associated with an unfavorable prognosis in colorectal cancer (33). It is possible that opposing actions of antigen-presenting cells can be exerted on tumor progression, depending on the polarization and activation status of DCs or macrophages. The DC phenotype can also be switched from immunoenhancing to immunosuppressive by factors secreted in the tumor microenvironment (34). Immunosuppressive DCs are associated with low expression of co-stimulatory molecules, blunting antigen presentation, and up-regulation of regulatory molecules such as programmed death 1 within tumors. Thus, further studies elucidating subsets and characteristics of antigen-presenting cells using different markers together with CD11c are warranted.
In conclusion, this study demonstrated a close correlation of CD11c expression with TILs and TLSs in TNBC. Therapeutic modalities enhancing the number and activation of DCs in the tumor microenvironment could facilitate infiltration of TILs and yield better clinical outcomes.
Acknowledgements
This study was supported by Basic Science Research Programs, through the National Research Foundation of Korea (NRF), funded by the Ministry of Science, ICT & Future Planning, Republic of Korea (2017R1D1A1B03035491 and 2017R1D1A1B03033104).
Footnotes
This article is freely accessible online.
Conflicts of Interest
The Authors declare no conflicts of interest in regard to this study.
- Received July 24, 2018.
- Revision received September 14, 2018.
- Accepted September 19, 2018.
- Copyright© 2018, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved