

Abstract
Polygonum cuspidatum is a traditional Chinese herbal medicine used in the treatment of various diseases. In the present study, we investigated whether the crude extract of Polygonum cuspidatum (CEPC) could affect immune responses of murine leukemia cells in vivo. Normal BALB/c mice were i.p. injected with WEHI-3 cells to generate leukemic mice and then were treated orally with CEPC at 0, 50, 100 and 200 mg/kg for three weeks. Animals were weighed and blood, liver, spleen samples were collected for further analyses. Results indicated that CEPC did not significantly affect the body and liver weight of animals, but reduced the weight of spleen when compared to control groups. Flow cytometric assay demonstrated that CEPC increased the percentage of CD3- (T-cell marker) and CD19- (B-cell marker) positive cells, but reduced that of CD11b-positive ones (monocytes). However, it did not significantly affect the proportion of Mac-3-positive cells (macrophages), compared to control groups. Results indicated that CEPC promoted phagocytosis by macrophages from blood samples at all examined doses but did not affect that of macrophages from the peritoneal cavity. CEPC also promoted natural killer cell activity of splenocytes at 200 mg/kg of CEPC. CEPC promoted B-cell proliferation at 200 mg/kg treatment when cells were stimulated with lipopolysaccharides but did not promote T-cell proliferation at three doses of CEPC treatment on concanavalin A stimulation.
- Crude extract of Polygonum cuspidatum
- leukemia
- Balb/c mice
- phagocytosis
- macrophage
- natural killer cells
Cancer remains a chronic disease causing major morbidity and mortality worldwide. Leukemia is a type of cancer caused by the unregulated proliferation of immature blood cells that are derived from mutant hematopoietic stem cells (1). It was reported that the malignant cells can be effectively trapped at their early stage of differentiation (2). In patients with leukemia, the main curative treatment is chemotherapy but the characteristics for this treatment are relatively low efficacy and high toxicity for the patient (3). Many reports have shown that increased consumption of fruit and vegetables may reduce the risk of cancer (4) and cardiovascular diseases and atherosclerosis (5). It was reported that antioxidant nutrients, dietary fiber and phytochemicals present in fruits and vegetables may potentially protect against cancer (6).
It is well-documented that after introduction of antigens into the human body, several white blood cell types are involved in interactions with each other to produce immune responses against invading foreign agents (7). It is well-known that the responses in leukemia to therapeutic agents differ from one type or subtype to another. In our earlier studies, we found some natural compounds and crude extracts of natural plants which can promote immune responses in leukemic mice (8-11). Thus, we are further investigating whether such compounds or crude extract from natural plants can promote immune responses in vivo.
The root of the plant Polygonum cuspidatum (Polygonaceae), widely distributed in southern China and Japan, has been used to treat inflammation, infection and hyperlipidemia (12). Emodin isolated from Polygonum cuspidatum has been demonstrated to have an inhibitory function against Coxsackievirus B4 in vitro and in vivo (13). Furthermore, emodin has been shown to have anticancer function against several human cancer cell lines (14-16). Our earlier studies also showed that emodin induced apoptosis of C6 cells after a 12-h treatment, but C6 cells survived a 72-h drug treatment, indicating resistance to emodin (17). We also found that crude extract of Polygonum cuspidatum (CEPC) induced immune responses in normal mice in vivo (18), however, there is no report to demonstrate the effect of CEPC on the immune responses in leukemic mice. Thus, in the present study, we investigated the immune responses of leukemic BALB/c mice in vivo when treated with CEPC.
Materials and Methods
Materials and reagents. Dimethyl sulfoxide (DMSO) was purchased from Sigma-Aldrich Corp. (St. Louis, MO, USA). Tissue culture plastic wares were obtained from BD Bioscience (Pharmingen Inc., San Diego, CA, USA). All products used for cell culture (RPMI-1640 medium, fetal bovine serum, L-glutamine and penicillin-streptomycin) were obtained from Gibco Life Technologies (Carlsbad, CA, USA). CEPC was kindly provided by Dr. Chao-Lin Kuo (Department of Chinese Pharmaceutical Sciences and Chinese Medicine Resources, China Medical University, Taichung, Taiwan, ROC) and was immediately dissolved in DMSO at 1% stock solution and kept at −20°C in a 50 ml tube covered with aluminum paper to avoid light before use and diluted as desired directly in medium.
Murine WEHI-3 leukemia cells. WEHI-3 cell line (murine myelomonocytic leukemia cells) was purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan, ROC). Cells were cultured in RPMI-1640 medium supplemented with 10% fetal bovine serum, 100 units/ml penicillin, 100 μg/ml streptomycin and 2 mM L-glutamine and maintained at 37°C with 5% CO2 in a humidified incubator (19, 20).
In order to maintain exponential growth, cells were split every four days before the experiments.
Male BALB/c mice. Fifty male Balb/c mice, aged 8-weeks-old (weighing from 20-25 g) were used in this study, obtained from the National Laboratory Animal Center (Taipei, Taiwan, ROC). All animals were housed in groups in stainless steel mesh-bottomed cages. They were kept under specified pathogen-free conditions in the animal center of China Medical University (Taichung, Taiwan, ROC). The mice were fed with clean water and normal nutrient food under standard conditions based on the institutional guidelines (Affidavit of Approval of Animal Use Protocol) that have been reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of China Medical University (Taichung, Taiwan, ROC) as described previously, approval number is 98-74-N (19).
In vivo treatment of animals with CEPC. Ten Balb/c mice received normal diet as control (group I). Forty Balb/c mice were randomly divided into four groups with 10 mice in each group. One day before dosing, every mouse was peritoneally inoculated with 1×106 WEHI-3 leukemia cells. Group II mice received normal diet as positive control. Group III mice received CEPC at 50 mg/kg. Group IV mice received CEPC at 100 mg/kg. Group V mice received CEPC at 200 mg/kg. CEPC in olive oil was administered by oral gavage to animals in each group for 16 days and then all mice were weighed and sacrificed by euthanasia with CO2 as described previously (21).
Immunofluorescence staining for surface markers of cells from each mouse in each group. At the end of treatment, each mouse was individually weighed and then blood and spleen was individually collected and splenocytes were isolated from each spleen for measuring natural killer (NK) cell activity as described previously (22). Leukocyte cell markers were measured in each blood sample. In brief, 1 ml blood from each mouse was collected and lysed to destroying the red blood cells with 1× Pharm Lyse™ lysing buffer (BD Biosciences) as per guidelines from BD Biosciences. All blood samples were centrifuged at 1500 × g for 15 min at 4°C to collect white blood cells and then were stained by Phycoerythrin (PE)-labeled anti-mouse CD3, PE-labeled anti-mouse CD19, Fluorescein isothiocyanate (FITC)-labeled anti-mouse CD11b and Mac-3 antibodies (BD Biosciences) for 30 min. All cells were washed with Phosphate-buffer saline (PBS) then were stained with secondary antibody and analyzed by flow cytometry for determining the percentage of cell markers as previously described (22).
Determinations of macrophage phagocytosis. Macrophages were isolated from peripheral blood mononucleated cells (PBMCs) and peritoneum of each mouse from each group as described previously (9, 22). Isolated cells (macrophages) were placed in plates and 50 μl of Escherichia coli-FITC added according to PHAGOTEST® kit manufacturer's instructions (ORPEGEN Pharma Gesellschaft für biotechnologische, Heidelberg, Germany). All samples were analyzed by flow cytometery, and quantified by CellQuest software (Becton Dickinson Immunocytometry System, Mountain View, CA) as described previously (9, 22).
Determination of NK cell cytotoxic activity. Isolated splenocytes from each mouse were placed in 96-well plate atadensity of 1×105 cells/well with each well containing 1 ml of RPMI-1640 medium. YAC-1 cells (a murine T-lymphoma cell line sensitive to NK-cells) (2.5×107 cells) in serum-free RPMI-1640 medium and PKH-67/Dil.C buffer (Sigma-Aldrich Corp.) based on the manufacturer' instructions were added to the cells in each well then mixed thoroughly for 2 min at 25°C. Then 2 ml PBS was added to each well for 1 min, 4 ml medium was added to the well and plants were incubated for 10 min. After incubation, cells were centrifuged at 453 rcf for 2 min at 25°C then NK cell cytotoxic activity was assayed by flow cytometry as described elsewhere (9, 22).
Determination of T- and B-cell proliferation. Isolated splenocytes at a density of 1×105 cells/well were placed in 96-well plates and 100 μl of RPMI-1640 medium was added to each well. For T-cell proliferation, concanavalin A (Con A, 5 μg/ml) (Sigma Chemical Co., St. Louis, MO) was added to the cells to stimulate for three days. For B-cell proliferation, lipopolysaccharide (LPS, 5 μg/ml) (Sigma Chemical Co., St. Louis, MO) was added to the cells to stimulate for five days. All samples were assayed by using CellTiter 96 AQueous One Solution Cell Proliferation Assay kit (Promega, Madison, WI, USA) as previously described (9, 22).

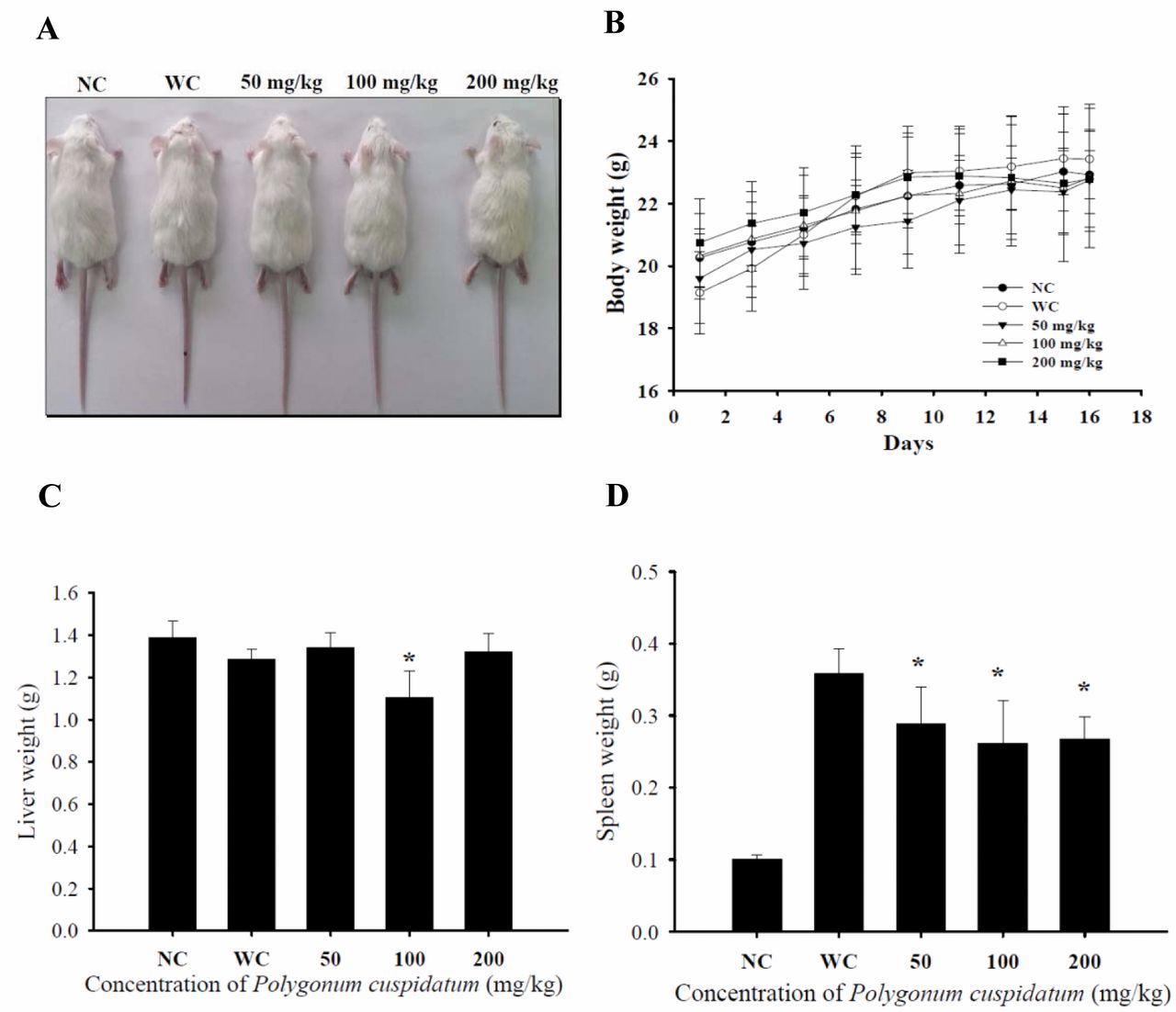
Crude extract of Polygonum cuspidatum (CEPC) effect on the body, liver and spleen weights of leukemic Balb/c mice. Group I was normal control (NC). Group II-V mice were injected WEHI-3 cells i.p. then were divided into four groups. Group II was treated with normal diet (WC). Group III was treated with 50 mg/kg of CEPC. Group IV was treated with 100 mg/kg of CEPC. Group V was treated with 200 mg/kg of CEPC. All animals were treated for 16 days. A representative image of one animal from each group (A), body (B), liver (C) and spleen (D) weights are presented. The total body weights were measured every two days. *Significant difference at p<0.05 between WC and CEPC-treated groups.
Statistical analysis. All data are expressed as mean±S.D. Comparisons between control and CEPC-treated groups were analyzed by the Student's t-test. For all comparisons, differences were considered statistically significant at p<0.05.
Results
CEPC affected the body weight of leukemic Balb/c mice. From the in vivo protocol, 10 Balb/c mice received normal diet as did the control, and another 40 WEHI-3-generated leukemic mice were divided into groups, one was the control and other three groups were treated with CEPC (50, 100 and 200 mg/kg) for 16 days. During the experimental period, mice were individually weighed every two days. A representative animal is presented in Figure 1A and its body weight, liver and spleen weigh are showing in Figure 1B, C and D, respectively. These results indicate that CEPC did not significantly affect body weight and liver weights (Figure 1B and C) but did reduce the spleen weight when compared to leukemic mice without CEPC treatment as shown in Figure 1D.
CEPC affected markers of white blood cells from leukemic Balb/c mice. All animals were sacrificed and blood samples were collected from each mouse from each group then the levels of cell markers CD3, CD19, CD11b and Mac-3 were measured by using flow cytometry (Figure 2). The results indicate that CEPC promoted cells carrying CD3 (Figure 2A) and CD19 (Figure 2B) but reduced the proportion of those carrying CD11b (Figure 2C). However, it did not significantly affect the proportion of cells bearing Mac-3 (Figure 2D).

Crude extract of Polygonum cuspidatum (CEPC) affected the levels of cell markers in white blood cells from leukemic Balb/c mice. Group I was normal control (NC). Group II-V mice were injected WEHI-3 cells i.p. then were divided into four groups. Group II was treated with normal diet (WC). Group III was treated with 50 mg/kg of CEPC. Group IV was treated with 100 mg/kg of CEPC. Group V was treated with 200 mg/kg of CEPC. All animals were treated for 16 days. Blood was collected from animals in each group and then analyzed for cell markers by flow cytometry as described in Materials and Methods. A: CD3; B: CD19; C: CD11b and D: Mac-3. The data are expressed as the mean±S.D. of three experiments (n=10). *Significant difference at p<0.05 between WC and CEPC-treated groups.
CEPC affected phagocytosis by macrophages from PBMCs and peritoneal cavity of leukemic BALB/c mice. After treatment, cells were collected from PBMCs and the peritoneal cavity of each animal from each group for measuring the levels of phagocytosis. CEPC treatment at both high doses significantly reduced macrophage phagocytosis from PBMCs (Figure 3A) but no dose of CEPC treatment affected phagocytotic activity of macrophages which were obtained from the peritoneal cavity (Figure 3B).
CEPC affected the cytotoxic activity of NK cells from leukemic BALB/c mice. For NK cell activity measurement, splenocytes from each mouse from each group were used with YAC-1 target cells and then were assayed by flow cytometry. The results indicate that YAC-1 target cells were killed by NK cells isolated from splenocytes of mice treated with the three dose levels of CEPC, when compared to normal mice; however, compared to leukemic mice, NK cell activity was not significantly affected.

Crude extract of Polygonum cuspidatum (CEPC) promoted phagocytosis of macrophages from peripheral blood mononucleated cells (PBMCs) and peritoneal cavity of leukemia Balb/c mice. Group I was normal control (NC). Group II-V mice were injected WEHI-3 cells i.p. then were divided into four groups. Group II was treated with normal diet (WC). Group III was treated with 50 mg/kg of CEPC. Group IV was treated with 100 mg/kg of CEPC. Group V was treated with 200 mg/kg of CEPC. All animals were treated for 16 days. Blood samples were collected from CEPC-treated and control animals then macrophages were isolated from PBMCs (A) and peritoneum (B) of each mouse. Phagocytosis of isolated macrophages was measured by flow cytometery and quantified by CellQuest as described in the Materials and Methods. *Significant difference at p<0.05 between WC and CEPC-treated groups.
CEPC affected proportion of B- and T-cells from leukemic BALB/c mice. Results from T- and B-cell proliferation assays indicate that CEPC at 50 and 200 mg/kg treatment promoted B-cell (Figure 5A) proliferation, however, CEPC treatment had no affect on T-cell proliferation (Figure 5B).
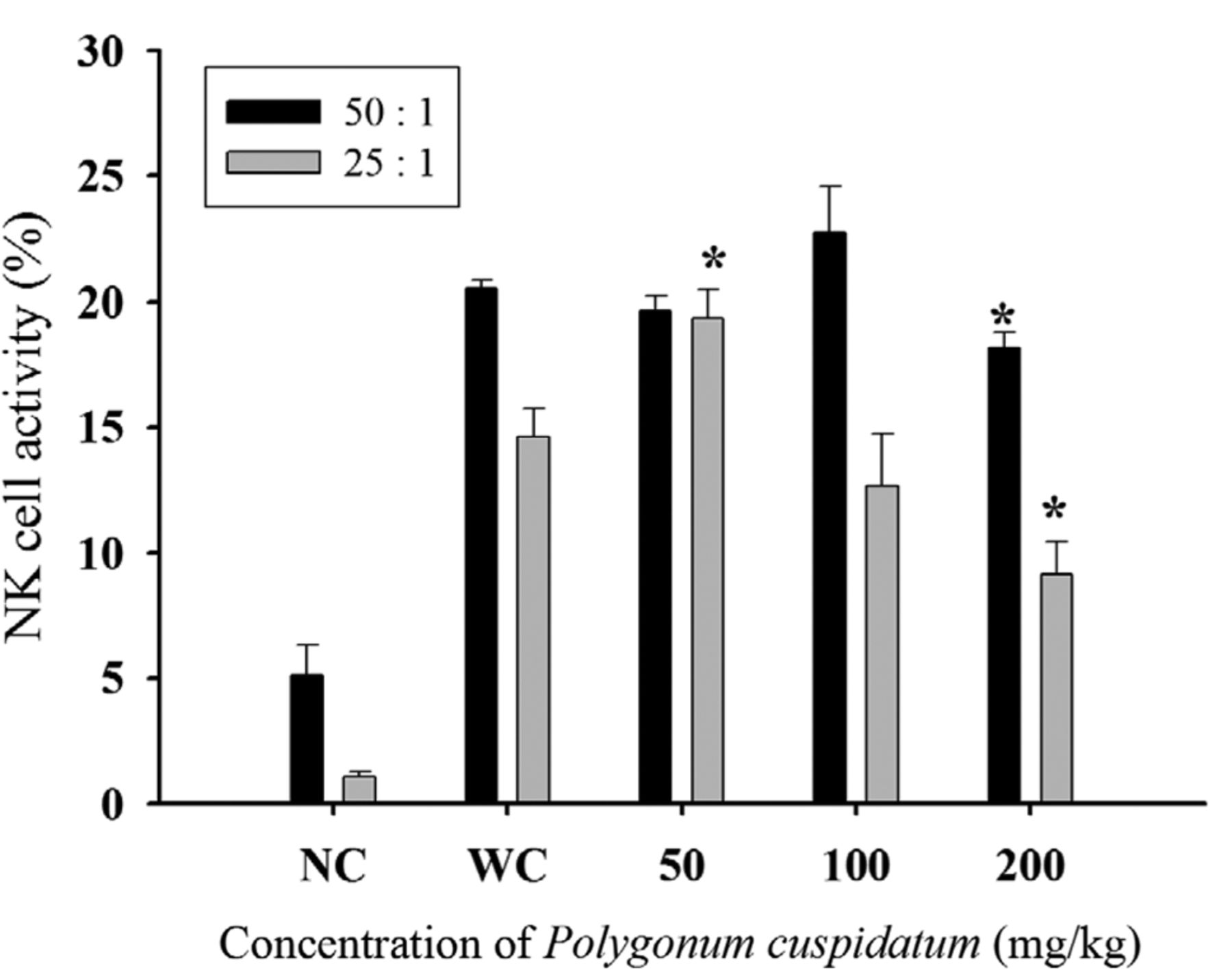
Crude extract of Polygonum cuspidatum (CEPC) affected the cytotoxic activity of natural killer (NK) cells in leukemic Balb/c mice. Group I was normal control (NC). Group II-V mice were injected WEHI-3 cells i.p. then were divided into four groups. Group II was treated with normal diet (WC). Group III was treated with 50 mg/kg of CEPC. Group IV was treated with 100 mg/kg of CEPC. Group V was treated with 200 mg/kg of CEPC. All animals were treated for 16 days. Isolated splenocytes (1×105 cells/well) were placed in 1 ml of RPMI-1640 medium in 96-well plates. Target YAC-1 cells (2.5×107 cells) with serum-free RPMI-1640 medium and PKH-67/Dil.C buffer was added to the cells for the determination of the NK cell cytotoxic activity by flow cytometry, as described in Materials and Methods. *Significant difference at p<0.05 between WC and CEPC-treated groups.
Discussion
Many plant-derived bioactive compounds, such as paclitaxel from Taxus brevifolia and camptothecin from Camptotheca acuminata, have been used as anticancer drugs in patients with cancer (23) and numerous studies have also focused on the extracts of traditional herbs as alternative and complementary medications for cancer therapy. There is not sufficient and reliable data regarding the biological activities (genotoxic and cytotoxic effects) of CEPC on cultured human cancer cells and animal studies. Previous studies on the immune responses from normal mice after exposure to various doses of CEPC show they promoted immune responses, in particular promoting macrophage phagocytosis (18). However, there are no available to show CEPC effects on immune responses in leukemic mice in vivo. Thus, in the present study, we investigated the effects of CEPC on immune responses of leukemic Balb/c mice in vivo.
Normal animals were injected i.p. with WEHI-3 cells and then treated with or without CEPC at various doses. The results indicated that CEPC at concentrations of 50, 100 and 200 mg/kg did not lead to significant differences of body and liver weight compared to the control groups (Figure 1B and C) but did reduce the weight of spleen when compared to leukemic mice without treatment of CEPC (Figure 1D). Our earlier studies showed that CEPC did not induce cytotoxic effects on normal animals (18). Herein, we found that CEPC enhanced cellular populations such as CD3-positive cells (T-cells) and CD19-positive cells (B-cells) and reduced CD11-positive cells (monocytes), but did not significantly affect the Mac-3-positive (macrophage) population in leukemic mice. These finding indicated that oral treatment of CEPC can increase T and B cell population. Numerous studies have demonstrated that macrophages suppress intracellular bacterial growth and can even destroy an infection in vivo (24-26). Because foreign bacteria entry to human body has been known to be associated with cancer development. However, Figure 3A indicates that CEPC significantly promoted macrophage phagocytosis in leukemic mice, thus, further studies on the effect of CEPC on macrophage function are needed.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Crude extract of Polygonum cuspidatum (CEPC) effects on T- and B-cell proliferation in leukemic Balb/c mice. Group I was normal control (NC). Group II-V mice were injected WEHI-3 cells i.p. then were divided into four groups. Group II was treated with normal diet (WC). Group III was treated with 50 mg/kg of CEPC. Group IV was treated with 100 mg/kg of CEPC. Group V was treated with 200 mg/kg of CEPC. All animals were treated for 16 days. A: Isolated B-cells were pretreated with lipopolysaccharide (LPS) and B-cell proliferation was analyzed by flow cytometry. B: T-Cells were pretreated with concanavalin A (Con A) and then cell proliferation was analyzed by flow cytometry. *Significant difference at p<0.05 between WC and CEPC-treated groups.
Results from Figure 4 indicate that CEPC at 50 mg/kg promoted NK cell activity of splenocytes but other doses of CEPC treatment did not have any significant effect. Furthermore, CEPC treatment did not significantly affect T-cell proliferation after Con A stimulation (Figure 5B), however, 50 and 200 mg/kg of CEPC treatment led to B-cell proliferation after LPS stimulation (Figure 5A). B cells can produce antibodies for connecting with antigens then macrophague can destroy them.
Acknowledgements
This work was supported by the grant CMU102-ASIA-20 from China Medical University, Taichung, Taiwan, ROC.
Footnotes
-
* These Authors contributed equally to this study.
- Received November 5, 2014.
- Revision received November 25, 2014.
- Accepted November 26, 2014.
- Copyright © 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved