

Abstract
Mastic is a resinous exudate obtained from the stem and the main leaves of Pistacia lentiscus. Mastic has shown several beneficial pharmaceutical properties such as antibacterial and apoptosis-modulating activities. The aim of this study was to investigate whether mastic affects the function of activated macrophages. Both solid and liquid types of mastic inhibited the production of pro-inflammatory substances such as nitric oxide (NO) and prostaglandin (PG)E2 by lipopolysaccharide (LPS)-activated mouse macrophage-like RAW264.7 cells. This was accompanied by the decline of viable cell number. Western blot and RT-PCR analyses showed that mastic inhibited the expression of inducible NO synthase (iNOS) and cyclooxygenase (COX)-2 at both protein and mRNA levels. ESR spectroscopy revealed that mastic scavenged NO and superoxide radicals very poorly, in contrast to its potent hydroxyl radical scavenging activity. These data demonstrate that mastic inhibits the production of both NO and PGE2 by activated macrophages mostly via its cytotoxic action. The narrow range of effective concentration of mastic due to its cytotoxicity may limit its potential application as an anti-inflammatory agent.
- Mastic
- RAW264.7 cells
- macrophage
- NO
- PGE2
- anti-inflammatory activity
Certain plants used in folk medicine serve as sources of therapeutic substances that have multipotential medicinal effects. Mastic is the extract of sap from Pistacia lentiscus, grown only in Chios island, Greece. Due to its unique shape and diverse efficacy, mastic has been called ‘tear drop of Christ’. This extract has been utilized for as long as 5,000 years as a food for the stimulation of health and in the treatment of digestive diseases (1). Mastic has shown therapeutic effects on stomach and duodenal ulcers (2-4) and has also shown antibacterial activity against Helicobacter pylori (5, 6). Recently, major triterpenic acids in the acid extract have been suggested to be responsible for its anti-H. pylori activity (7). Mastic chewing gum has also shown antiplaque activity in the oral cavity (8). Mastic has been reported to inhibit the growth of colon cancer (9), prostate cancer (10) and erythroleukemia cell line (11). We recently found that mastic induced apoptosis in human leukemia cells without induction of differentiation, whereas it inhibited spontaneous apoptosis of oral polymorphonuclear cells (OPMNs) possibly by its antioxidant activity (12). However, the study of the anti-inflammatory activity of mastic has been limited. We investigated here whether mastic inhibits the production of pro-inflammatory substances such as nitric oxide (NO) and prostaglandin E2 (PGE2) by activated macrophages and if so, whether mastic inhibits inducible NO synthase (iNOS) and cyclooxygenase-2 (COX-2) expression that regulate the amount of NO and PGE2, respectively. For this purpose, we used mouse macrophage-like RAW264.7 cells, established from the peritoneal fluid of BALB/c mice (13). RAW264.7 cells activated by lipopolysaccharide (LPS) have been used by many investigators for the study of the signal transduction pathway during the activation process of macrophages (14, 15) and in the search for substances that modify macrophage function (16).
Materials and Methods
Materials. The following chemicals and reagents were obtained from the indicated companies: mastic [solid form, liquid form (which contains mastic and coconut oil at the ratio of 3:7)] (Nakamura Chiro Association, Tokyo, Japan); Dulbecco's modified Eagles medium (DMEM) (Gibco BRL, Grand Island, NY, USA); fetal bovine serum (FBS) (JRH Bioscience, Lenexa, KS, USA); LPS from Escherichia coli (Serotype 0111:B4), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), hypoxanthine (HX), xanthine oxidase (XOD) (Sigma Chem Co., St Louis, MO, USA); dimethyl sulfoxide (DMSO) (Wako Pure Chem. Ind., Osaka, Japan); 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxy1-3-oxide (carboxy-PTIO; a spin trap agent), 1-hydroxy-2-oxo-3-(N-3-methyl-3-aminopropyl)-3-methyl-1-triazene (NOC-7; an NO generator), diethylenetriaminepentaacetic acid (DETAPAC), 5,5-dimethyl-1-pyrroline-N-oxide (DMPO) (Dojin, Kumamoto, Japan).
Cell culture. RAW264.7 cells (13) (kindly supplied by Professor Ohmori, Meikai University) were cultured as an adherent monolayer culture in DMEM supplemented with 10% heat-inactivated FBS under a humidified 5% CO2 atmosphere (16, 17). When the cells were grown to confluency, they were detached by being splashed with the medium through the pipette, and seeded at lower cell density. Cells were not collected by scraping them off of the plate with a rubber policeman, since the scraping procedure damaged the cells (unpublished data).
Determination of viable cell number. RAW264.7 cells were inoculated at 0.8×106/ml (100 μl) in a 96-microwell plate (Becton Dickinson) and incubated for 1-2 hours. When cells were near-adherent, media were replaced with fresh culture media and cells were treated for 24 hours with the indicated concentrations of mastic solid form (0-100 μg/ml) or liquid form (0-0.5%) in phenol red-free DMEM supplemented with 10% FBS in the presence or absence of LPS (0.1 μg/ml). Viable cell number was then determined by MTT method. In brief, cells were incubated for 60 minutes with 0.2 mg/ml of MTT. After removal of the medium, the cells were lysed with DMSO, and the absorbance at 540 nm was recorded using a microplate reader (16, 17).
Assay for NO production. RAW264.7 cells were inoculated at 0.8×106/ml (100 μl) in a 96-microwell plate and incubated for 1-2 hours. Near confluent cells were treated for 24 hours with the indicated concentrations of mastic: solid form (0-100 μg/ml) or liquid form (0-0.5%) in phenol red-free DMEM supplemented with 10% FBS in the presence or absence of LPS (0.1 μg/ml). The NO production by RAW264.7 cells was quantified by Greiss reagent, using the standard curve of NO2-. To eliminate the interaction between sample and Greiss reagent, the NO concentration in the culture medium without the cells was also measured and this value subtracted from that obtained with the cells. The concentration that inhibited the LPS-stimulated NO production by 50% (50% inhibitory concentration: IC50) was determined from the dose-response curve (16). The efficacy of inhibition of NO production was evaluated by the selectivity index (SI), which was calculated using the following equation: SI=CC50/IC50.
Assay for prostaglandin E2 (PGE2) production. RAW264.7 cells were inoculated at 0.8×106/ml (100 μl) in a 96-microwell plate and incubated for 1-2 hours. Near-adherent cells were treated for 24 hours with different concentrations of mastic: solid form (0-100 μg/ml) or liquid form (0-0.5%) in the presence of LPS (0.1 μg/ml). The culture supernatant was collected by centrifugation and PGE2 concentration determined by EIA kit (Cayman Chemical Co, Ann Arbor, MI, USA) (18).
Assay for iNOS protein expression. RAW264.7 cells were inoculated at 0.8×106/ml (100 μl) in 96-well plates (Becton Dickinson) and incubated for 1-2 hours. Near-adherent cells were treated for 24 hours with different concentrations of mastic: solid form (0-100 μg/ml) or liquid form (0-0.5%). The concentrations were selected by CC50 in the presence or absence of LPS (0.1 μg/ml). The cell pellets were lysed with 50 μl of lysis buffer [10 mM Tris-HCl (pH 7.6), 1% Triton®X-100, 150 mM NaCl, 5 mM EDTA-2Na and 2 mM PMSF] for 10 minutes on ice. The cell lysates were centrifuged at 16,000×g for 20 minutes at 4°C to remove the insoluble materials and the supernatant was collected. The protein concentration of the supernatant was measured by Protein Assay Kit (Bio Rad, Hercules, CA, USA). Equal amounts of the protein from cell lysates (10 μg) were mixed with 2×sodium dodecyl sulfate (SDS)-sample buffer [0.1 M Tris-HCl (pH 6.8), 20% glycerol, 4% SDS, 0.01% bromophenol blue, 1.2% 2-mercaptoethanol], boiled for 10 minutes and subjected to SDS-8% polyacrylamide gel electrophoresis, and then transferred to polyvinylidene difluoride (PVDF) membrane. The membranes were blocked with 5% skim milk in phosphate-buffered saline [PBS (-)] plus 0.05% Tween 20 for 90 minutes and incubated for 90 minutes at room temperature with anti-COX-2 or anti-iNOS (dilution: 1:4,000 and 1:1,000, respectively; Santa Cruz Biotechnology, Delaware, CA, USA) or anti-actin antibody (1:6,000; Sigma), and then incubated with horseradish peroxidase-conjugated anti-goat, anti-rabbit or anti-mouse IgG for 60 minutes at room temperature. Immunoblots were developed with a Western Lighting™ Chemiluminescence Reagent plus (16).
Assay for iNOS and COX-2 mRNA expression. RAW264.7 cells were inoculated at 0.8×106/ml (100 μl) in 96-well plates and incubated for 1-2 hours. Near-confluent cells were treated for 24 hours with different concentrations of mastic: solid form (0-100 μg/ml) or liquid form (0-0.5%) in the presence or absence of LPS (0.1 μg/ml). The expression levels of iNOS and COX-2 mRNA were measured by RT-PCR. The primer sequence and size was: (i) iNOS: PCR product size (496 bp); sense primer (5′-CCCTTCCGAAGTTTCTGGCAGCAGC-3′), antisense primer (5′-GGCTGTCAGAGCCTCGTGGCTTTGG-3′); (ii)COX-2: Product size (371 bp); sense primer (5′-TTTGTTG AGTCATTCACCAGACAGAT-3′), antisense primer (5′-CAGTATTG AGGAGAACAGATGGGATT-3′); (iii)G3PDH: Product size (452 bp), sense primer (5′-ACCACAGTCCATGCCATCAC-3′), antisense primer (5′-TCCACCACCCTGTTGCTGTA-3′) (16).
NO radical-scavenging activity. The radical intensity of mastic was determined at 25°C, using electron spin resonance (ESR) spectroscopy (JEOL JES REIX Tokyo, Japan, X-band, 100 kHz modulation frequency) (16, 19). The radical intensity of NO, produced from the reaction mixture of 20 μM carboxy-PTIO and 60 μM NOC-7, was determined in 0.1 M phosphate buffer, pH 7.4, in the presence of 30% DMSO (microwave power and gain were 5 mW and 400, respectively). When NOC-7 and carboxy-PTIO were mixed, NO was oxidized to NO2 and carboxy-PTIO was reduced to carboxy-PTI, which produces seven-line signals. Samples were added 3 minutes after mixing. The NO radical intensity was defined as the ratio of the signal intensity of the first peak of carboxy-PTI to that of MnO, and expressed as the ratio to the height of MnO an external marker.
The superoxide anion (in the form of DMPO-OOH), produced by the HX-XOD reaction (total volume: 200 μl) (2 mM HX in 0.1 M phosphate buffer [pH 7.4] [PB] 80 μl, 1 mM DETAPAC 20 μl, 10% DMPO 30 μL, sample 40 μl, XOD [0.5 U/ml in PB] 30 μl) was determined (microwave power: 16 mW, gain: 630) (19).
For the determination of hydroxyl radical (in the form of DMPO-OH), produced by the Fenton reaction (200 μl)[1 mM FeSO4 (containing 0.2 mM DETAPAC) 50 μl, 0.1 M phosphate buffer (pH 7.4) 50 μl, 92 mM DMPO 20 μl, sample (in H2O) 50 μl, 1 mM H2O2, 30 μl], the gain was changed to 400 (19).

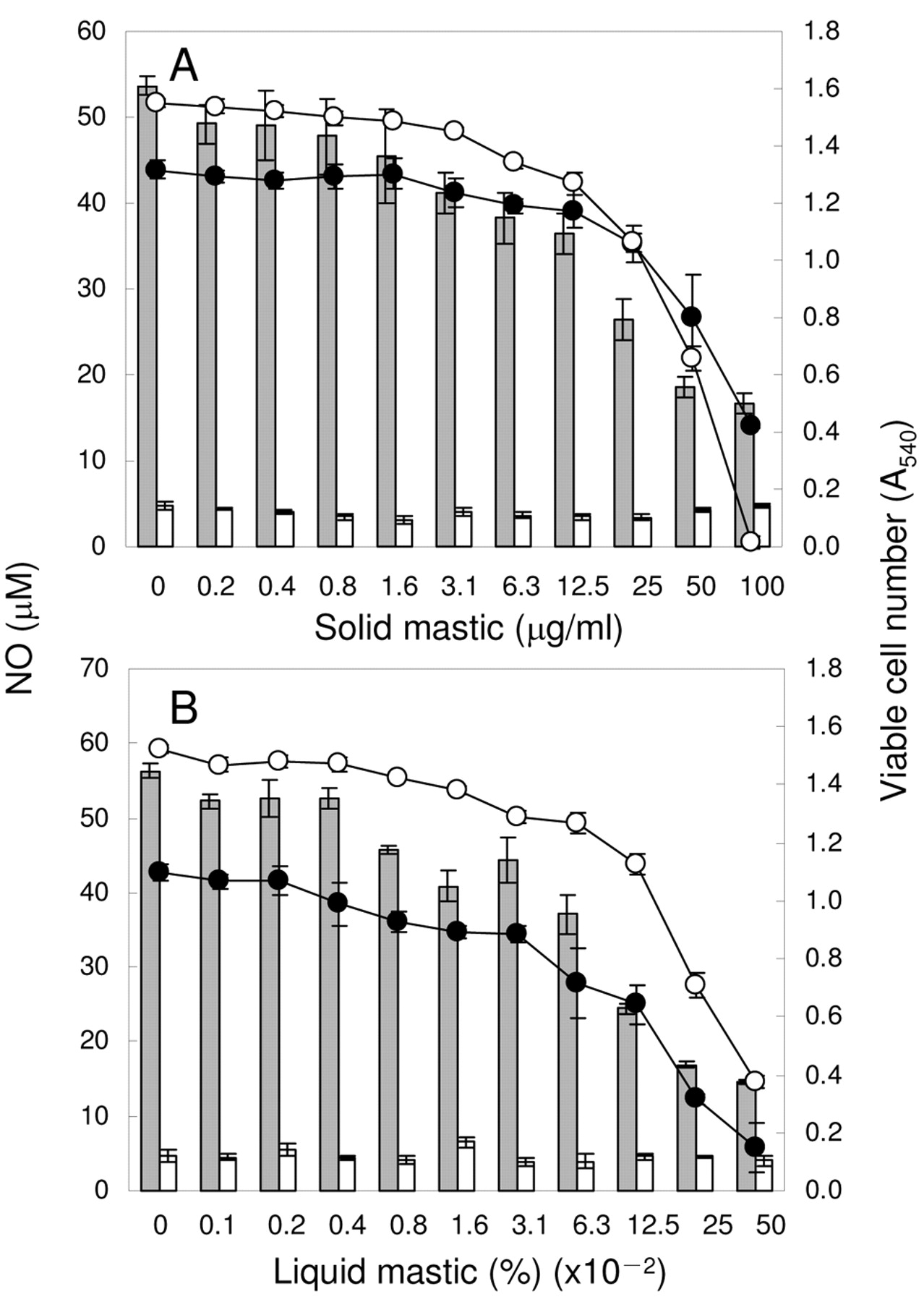
Inhibition by mastic of LPS-stimulated NO production. RAW264.7 cells were incubated for 24 hours with the indicated concentrations of solid (A) or liquid (B) mastic in the presence (gray or black) or absence (white) of 0.1 μg/ml LPS and the viable cell number (circle) and extracellular concentration of NO (bar) were then measured. Each value represents mean±SD from three independent experiments.
Results
Inhibition of NO and PGE2 production. We first confirmed that the optimal concentration of LPS for the stimulation of NO production by RAW264.7 cells was 0.1 μg/ml (data not shown). LSP (0.1 μg/ml) dramatically stimulated NO production [from 4.0±0.6 μM (basal level) to 55.0±1.4 μM] (Figure 1). Both solid and liquid types of mastic dose-dependently inhibited the LPS-stimulated NO production by RAW264.7 cells [IC50=29±2.3 μg/ml (solid), 0.07±0.02% (liquid), respectively] (Figure 1A, B). Although the inhibition of NO production by mastic was coupled with a reduction in the cell viability [CC50=61±4.0 μg/ml (solid), 0.23±0.07% (liquid), respectively], the inhibition of NO production cannot be explained solely by the cytotoxicity of mastic, since the selectivity index (SI=CC50/IC50) was 2.1±0.06 (solid) and 3.0±0.33 (liquid), respectively (Table I).

Inhibition by mastic of LPS-stimulated PGE2 production. RAW264.7 cells were incubated for 24 hours with different concentrations of solid (A) or liquid (B) mastic in the presence or absence of 0.1 μg/ml LPS, and then the viable cell number (circle) and extracellular PGE2 concentration (bar) were determined. Each value represents mean ±SD from three independent experiments.
We found that untreated RAW264.7 cells produced a background level of PGE2 (0.76±0.11 ng/ml), and the addition of 0.1 μg/ml LPS caused a two-order increase of PGE2 production (64.6±0.4 ng/ml) (Figure 2). Mastic dose-dependently inhibited the LPS-stimulated PGE2 production [IC50=49±0.17 μg/ml (solid), 0.31±0.04% (liquid), respectively] (Figure 2A, B), yielding an SI value of 1.9±0.014 (solid), 1.2±0.15 (liquid), respectively] (Table I).
Inhibition of NO and PGE2 production by solid and liquid mastic in the LPS-stimulated RAW264.7 cells. RAW264.7 cells were incubated for 24 hours with various concentrations of solid or liquid mastic in the presence of 0.1 μg/ml LPS, and then the viable cell number and extracellular NO and PGE2 were determined. From the dose-response curve, the 50% cytotoxic concentration (CC50) and the 50% inhibitory concentration of NO and PGE2 production (IC50) were determined. Selectivity index (SI) was determined by dividing the CC50 value by the IC50 value.
Inhibition of iNOS and COX-2 expression. We next investigated whether the inhibition of NO and PGE2 production by both solid and liquid types of mastic is due to the reduced expression of iNOS and COX-2 proteins, respectively. Western blotting analysis showed that this was the case (Figure 3). It is apparent that the dose-dependent inhibitory effect of both mastic samples on the expression of iNOS protein paralleled that of COX-2 protein. At the highest concentration tested (100 μg/ml or 0.5%), both iNOS and COX-2 protein expressions were below the detection limit.
RT-PCR experiments demonstrated that both solid and liquid types of mastic dose-dependently reduced the expression of iNOS and COX-2 mRNA expression, but not so drastically. At the highest concentration tested, iNOS and COX-2 mRNAs were still detectable (Figure 4).
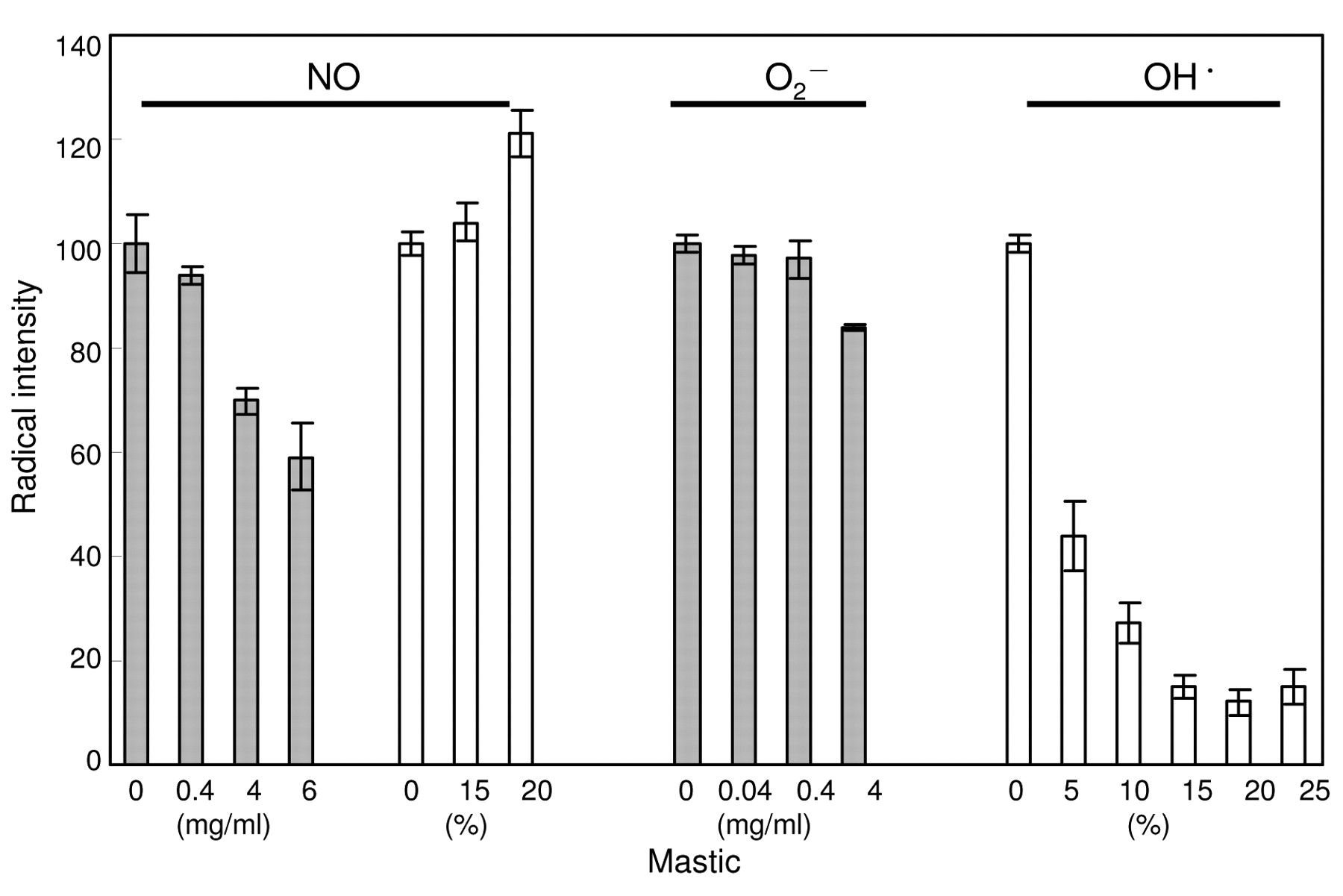
Radical-scavenging activity of mastic. The amount of NO is regulated by both the synthesis and degradation of NO. We next investigated the possibility that the reduced amount of NO recovered from the LPS-stimulated RAW264.7 cells in the presence of mastic might be due to the direct scavenging activity of mastic against NO. In vitro experiments with ESR spectroscopy shows that this was not the case (Figure 5). Solid mastic dose-dependently, but only slightly reduced the radical intensity of NO, generated from NOC-7 in the presence of carboxy-PTIO. However, at 0.4 mg/ml (14 times the concentration of IC50 for the inhibition of NO production), only 6% of NO radical was scavenged. Liquid mastic at 15 or 20% did not inhibit, but rather enhanced the radical intensity of NO. We also found that solid mastic (0.04-0.4 mg/ml) did not scavenge O2- (generated by HX-XOD reaction) that is known to counteract NO. On the other hand, liquid mastic scavenged the hydroxyl radical generated by the Fenton reaction. We did not assay the hydroxyl radical-scavenging activity of solid mastic (dissolved in DMSO), since DMSO reacts with hydroxyl radical to produce a carbon radical.
Discussion
The present study demonstrated for the first time that mastic (both solid and liquid) inhibited the NO and PGE2 production by LPS-activated RAW264.7 cells via the inhibition of iNOS and COX-2 protein expression, rather than by NO scavenging. We also found that mastic inhibited iNOS and COX-2 mRNA expression much less effectively, suggesting the possible post-transcriptional modification of iNOS and COX-2 mRNA. Mastic was highly cytotoxic to RAW264.7 cells, as revealed by a low SI for NO and PGE2 production (Table I). The sensitivity of RAW264.7 cells against both solid and liquid types of mastic (CC50=61-91 μg/ml; 0.23-0.37%) was higher than that of human oral squamous cell carcinoma (HSC-2, HSC-3, HSC-4) (81-126 μg/ml; 0.24-2.0%), human glioblastoma (T98G, U87MG) (127-158 μg/ml; >2.0%), human hepatocellular carcinoma cell lines (48-144 μg/ml; >2.0%) and human normal oral cells (HGF, HPC, HPLF) (93-166 μg/ml; >2.0%) (12). To further characterize the active principle that is responsible for the inhibition of NO and PGE2 production by activated macrophages, it is essential to remove the cytotoxic substances.
Mastic contains essential oils (mostly α-pinene and β-myrcene) at 2-5%, poly β-myrcene at approximately 25%, and acidic and neutral diterpenes and tricyclic, tetracyclic and pentacyclic triterpenes at 75% . By chromatographic separation, the acid fraction gave the major triterpenic acids, while the neutral fraction gave several triterpenic alcohols and aldehydes. The acid fraction reduced H. pylori colonization more effectively than did the neutral fraction, and the most active pure compound was identified as isomasticadienolic acid (7). Gas chromatographic and mass spectroscopic analysis has demonstrated that α-pinene, β-myrcene, β-pinene, limonene, β-caryophyllene are the major constituents of mastic, while the minor components such as verbenone, α-terpineol and linalool were identified as the most potent antibacterial substances against Escherichia coli, Staphylococcus aureus and Bacillus subtilis (20). It remains to be investigated which components in mastic inhibit NO and PGE2 production by activated macrophages.

Inhibition of iNOS and COX-2 protein expression by mastic in LPS-stimulated RAW264.7 cells. RAW264.7 cells were treated for 24 hours with the indicated concentrations of solid (upper) or liquid (lower panel) mastic in the presence or absence of LPS (0.1 μg/ml) and processed for Western blot analysis for iNOS, COX-2 and actin (internal marker) protein expression. Representative pattern of Western blot analysis of three independent experiments are shown.
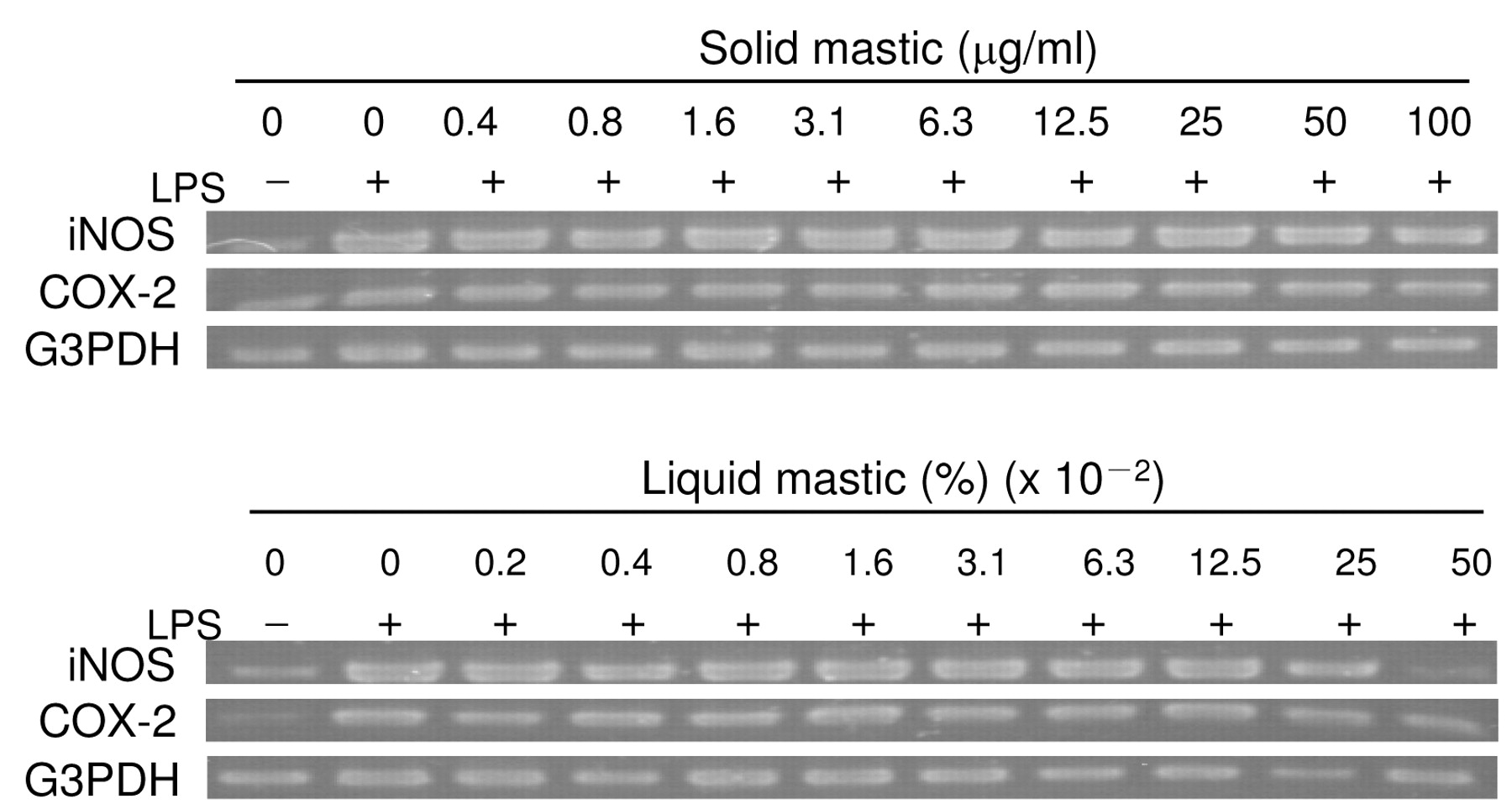
Inhibition of iNOS and COX-2 mRNA expression by mastic in LPS-stimulated RAW264.7 cells. RAW264.7 cells were treated for 24 hours with the indicated concentrations of solid (upper) or liquid (lower panel) mastic in the presence or absence of LPS (0.1 μg/ml) and processed for RT-PCR analysis for iNOS, COX-2 and G3PDH (internal marker) mRNA expression. Similar data were obtained in another two experiments.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Radical scavenging activity of solid (gray bar) and liquid mastic (white bar). The NO, O2- and OH• radical intensity was defined as the ratio of the signal intensity of the first peak of carboxy-PTI, DMPO-OOH or DMPO-OH to that of MnO, respectively, and expressed as a percentage that of the control (without mastic). Each value represents mean±S.D. from triplicate assays.
In contrast to lower molecular compounds in mastic, higher molecular weight substances such as poly β-myrcene have never been analyzed for their biological activity, due to their higher insolubility. We have reported unique biological activity of high molecular weight lignified materials in various plants such as Sasa senanensis Rehder (19), pine cone (21) and cacao husk (22). The biological activity of poly β-myrcene has yet to be investigated.
In conclusion, the present study demonstrated for the first time that mastic inhibits the production of both NO and PGE2 by activated macrophages mostly via its cytotoxic action. The narrow range of effective concentration of mastic due to its cytotoxicity may limit its potential application as an anti-inflammatory agent.
Acknowledgements
This study was supported in part by a Grant-in-Aid from the Ministry of Education, Science, Sports and Culture of Japan (Sakagami, No. 19592156).
- Received February 24, 2009.
- Revision received May 6, 2009.
- Accepted May 15, 2009.
- Copyright © 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved