


Abstract
Background/Aim: Although taurolidine is known to exert a wide spectrum of biological actions, its effects on immune cells have not been characterized in detail. In this study, we investigated the ex vivo effects of taurolidine on relevant innate and adaptive immune cell functions. Materials and Methods: Leukocyte functions in whole blood were assessed following treatment with various taurolidine concentrations. Viability of peripheral blood mononuclear cells (PBMCs) and granulocytes was measured using the WST-1 assay. PBMC function was assessed by measuring TNFα and IFNγ production after stimulation with lipopolysaccharide (LPS) or Candida, respectively. Reactive oxygen species (ROS) production by granulocytes was measured in whole blood using luminol-enhanced chemiluminescence. Granulocyte degranulation and activation were evaluated by membrane expression of degranulation (CD63, CD66B) and adhesion markers (CD62L, CD11b) using immunofluorescent staining followed by flow-cytometric analysis. Results: Taurolidine decreased viability of PBMCs and granulocytes: after 2 h, IC50 concentrations were 500 and 520 μg/ml, respectively. Following prolonged exposure (≥24 h) of PBMCs, the IC50 concentrations declined to 40 μg/ml. PBMC cytokine production significantly decreased at taurolidine concentrations below the cytotoxic threshold, whereas no changes in ROS production were observed. The expression of all granulocyte adhesion and degranulation markers increased at concentrations higher than 500 μg/ml (the cytotoxic level of taurolidine). Conclusion: Taurolidine exhibits a dose- and time-dependent cytotoxicity toward PBMCs and granulocytes. The effects on PBMCs, as exemplified by a decrease in cytokine production, occurred below the toxic threshold, whereas granulocyte function (ROS production) remained unchanged at these taurolidine concentrations. Granulocyte activation and degranulation markers only increased at cytotoxic taurolidine concentrations.
- Taurolidine
- leukocytes
- reactive oxygen species
- cell markers
- CD63
- CD66B
- CD62L
- CD11b
- cytokines
- cytotoxicity
- granulocyte
- PBMC
- neutrophil
Taurolidine [bis-(1,1-dioxoperhydro-1,2,4-thiadiazinyl-4)methane] is a chemical consisting of two cross-linked taurinamide rings, derived from the naturally occurring amino acid taurine. Hydrolysis of taurolidine produces the active metabolites taurultam and methylol-taurultam, which is finally metabolized from taurinamide to taurine, CO2, and H2O (1). In the mid-1970s, the therapeutic use of taurolidine was first described in the form of a topical rinsing solution because of its broad-spectrum bactericidal activity to treat intraperitoneal infections (2, 3). Two decades later, its efficacy as a catheter locking solution to prevent intravenous catheter-related bloodstream infections was recognized in the fields of oncology, hemodialysis, and long-term home parenteral nutrition (4-7).
The underlying mechanisms behind the antimicrobial effects of taurolidine include the inhibition of microbial adhesion to epithelial cell or endoluminal catheter surfaces, followed by the destruction of microbial cell membranes (1, 8-11). These effects are caused by downstream metabolites such as N-methylol derivatives of taurultam and taurinamide. Taurolidine has also been shown to modulate immunoregulatory properties of immune cells through its taurine moiety. It inhibits the production of the pro-inflammatory cytokines Interleukin (IL)-1 and Tumor Necrosis Factor α (TNFα) by peripheral blood mononuclear cells (PBMCs), and IL-6 and IL-8 by peritoneal and hepatic cells, respectively (12-16). Decreased toxicity through the binding of taurolidine to microbial endotoxins has also been reported (1, 17).
In the late 1990s, in a completely different field, antineoplastic properties of taurolidine in murine and human tumor cell lines were discovered. Taurolidine appeared to induce cancer cell death through a variety of mechanisms, including enhancement of apoptosis, inhibition of angiogenesis and tumor adherence, inhibition of pro-inflammatory cytokine and endotoxin levels, and stimulation of immune cell response to surgically induced trauma (18).
Whereas most effects of taurolidine are attributed to its antimicrobial properties, others clearly seem to be related to altered immune cell functions. Since these effects have not been characterized in much detail in immune cells, we embarked on the present investigation, given that the spectrum for the therapeutic use of taurolidine is rapidly expanding, ranging from surgery and oncology to hemodialysis and home parenteral nutrition.
The aim of this study was to investigate the effects of 2% taurolidine in 5% polyvinylpyrrolidone (PVP), a frequently used dosage in these clinical settings, on the modulation of ex vivo granulocyte and PBMC functions, and cytotoxicity following short and prolonged treatment periods.
Materials and Methods
Materials. Taurolidine was kindly provided by Geistlich Pharma AG (Wolhusen, Switzerland). Taurolidine’s vehicle PVP was purchased from Sigma-Aldrich (St Louis, MO, USA). The research ethics committee of the Radboudumc in Nijmegen, the Netherlands approved this study (reference number 2021-8310). Blood samples were drawn from twelve healthy volunteers after informed consent was obtained and processed as described below. These volunteers comprised an equal number of men and women between the age of 20 and 55 years.
Methods
Cell isolation. Cells were isolated from venous whole blood in 10 ml Monoject tubes containing 170 IU of lithium heparin (Belliver Industrial Estate, Plymouth, UK). Blood, diluted with an equal volume of phosphate-buffered saline (PBS, B. Braun, Melsungen, Germany), was layered onto FicollPaque™ Plus (ρ 1.077 g/ml, GE Healthcare, Uppsala, Sweden) and centrifuged for 20 min at 700×g. The lymphocyte-containing interphase (PBMCs) was washed twice and suspended in RPMI medium, supplemented with 5% heat-inactivated human plasma, 1% pen/strep, 1% pyruvate and 1% glutamax (stable glutamine) to the desired final concentration. The granulocyte-containing pellet was suspended in 50 ml ice-cold lysis solution (8.3 g/l NH4CL, 105 g/l KHCO3, pH=7.4) for 10 min. After centrifugation (400×g, 5 min), the remaining erythrocytes were lysed in fresh lysis solution for another 5 min. The granulocytes were washed twice and suspended in RPMI medium to the desired final concentration.
Cell survival assay. The WST-1 assay (a tetrazolium reduction assay) was used to assess cell viability and toxic effects of taurolidine and/or PVP on PBMCs and granulocytes. Per well, 25×104 PBMCs and 5×104 granulocytes were seeded in a 96-well flat bottom plate. Cells were exposed to three dilution series, comprising of two-fold dilution series of 5.000-5 μg/ml taurolidine in RPMI, two-fold dilution series of 25.000-25 μg/ml PVP in RPMI, and a two-fold dilution series of 2.000-2 μg/ml taurolidine/5% PVP in RPMI (10% v/v). The effect of each concentration was assayed in triplicate. After 2 h and 4 h (all cell types), and 24 h and 48 h (PBMCs only), cells were incubated with 10% (v/v) WST-1 reagent (Roche Diagnostics, Mannheim, Germany). After 4 h of incubation, the absorbance was measured at a wavelength of 435 nm, with a background correction read at 620 nm, according to the manufacturer’s instructions. The extinction was measured using the Tecan Infinite M200Pro and analyzed with Magellan™ software (Tecan Trading AG, Männedorf, Switzerland). Granulocytes were not assessed at 24 and 48 h due to their short viability ex vivo.
Cytokine production in PBMCs
Cell stimulation. PBMCs (5×105 cells per 90 μl) were seeded into a flat bottom 96-well plate and stimulated with either LPS (10 ng/ml, for 4 and 24 h) or heat-killed Candida albicans (CA, 10 microorganisms/ml, for 48 h). Simultaneously, PBMCs were exposed (v/v 10%) to different concentrations of taurolidine and/or the vehicle PVP. The taurolidine concentration series in which taurolidine was dissolved in RPMI medium comprised concentrations of 0, 10, 50, and 100 μg/ml w/v. The second concentration series comprised taurolidine dissolved in 5% PVP in water (w/v). These concentration series were acquired from the cell survival assay. Last, cells were treated with 0, 2.5, and 5 mg/ml PVP dissolved in water. For the treatment series containing PVP, a control for the quantity of medium was included, containing cells in medium and PBS (10% v/v). Each measurement was performed in duplicate.
LPS and CA were chosen as a model for bacterial and fungal infections, respectively. In addition, LPS and CA have different mechanisms of action; LPS stimulated cytokine production is endotoxin-induced, whereas heat-killed CA does not produce toxins and cytokine production is related to phagocytosis.
Cytokine measurements. Short and long-term effects of taurolidine were tested by measuring the levels of the cytokines TNFα and IFNγ in the supernatant of PBMCs treated with LPS and CA, respectively. Cytokine production was determined using a specific enzyme-linked immune sorbent assay (ELISA) kit for TNFα and IFNγ (R&D DuoSet®, Minneapolis, MN, USA) according to instructions of the manufacturer.
Oxygen radical production. Stimulus-induced oxygen radical production was evaluated by means of luminol-enhanced chemiluminescence using the automated TriStar2 Multimode Reader LB942 (Berthold Technologies, Bad Wilbad, Germany), as described in detail previously (19). Whole blood samples were incubated at 37°C for two h by gentle head-over-head turning with 0, 100, and 500 μg/ml taurolidine in 5% PVP solution (v/v 10%). This concentration series was acquired from the cell survival assay. Subsequently, the samples were diluted 100 times with Hanks’ Balanced Salt Solution (HBSS, Gibco, Paisley, UK). In a 96-well microplate, 200 μl of each sample was added to 20 μl phorbol 12-myristate13-acetate (PMA, receptor independent stimulus; a 0.1 mg/ml stock diluted 1:20 in HBSS) or serum-treated zymosan (STZ, receptor dependent stimulus; 10 mg/ml stock). To each well, 20 μl luminol (a 10 mmol/l stock, diluted 1:10 in HBSS/0.5% bovine serum albumin) was added. Each measurement was performed in quintuple. Luminescence was expressed as relative light units per second (RLU/s) and measured on a 425 nm at 37°C during 90 min. The area under the curve (AUC), representing the overall oxygen radical production, was analyzed with MikroWin 2000 software (Mikrotek Laborsysteme GmbH, Overath, Germany).
Leukocyte membrane surface antigen expression. The expression of activation molecules on the membrane surface of leukocytes was determined using immunofluorescent staining followed by flow-cytometric analysis, as previously described (19). Granulocyte surface marker characterization was performed using antibodies (purchased from Beckman Coulter, Miami, FL, USA) directed against an adhesion molecule of the β-integrin family (CD11b), degranulation markers (CD66b and CD63), and L-selectin (CD62-L). Blood samples were incubated at 37°C for two h by gentle head-over-head turning with 0, 10, and 500 μg/ml taurolidine in 5% PVP solution (v/v 10%). This concentration series was acquired from the cell survival assay. In total, 100 μl of each treated blood sample was incubated with 5 μl of each directly fluorochrome-labeled antibody for 15 min. Antibodies against the following proteins were used: CD66b and CD62L, labeled with fluorescein isothiocyanate (FITC); CD11b and CD63 labeled with phycoerythrin (PE); CD45, a leukocyte marker, labeled with phycoerythrin cyanin 7 (PC7); and CD14, a monocyte marker, labeled with PE-cytochrome 5 (PE-CY5). Subsequently, erythrocytes were lysed using 4 ml of BD Pharm Lyse™ solution (BD Biosciences, San Jose, CA, USA). The cell suspension was centrifuged at 400×g at room temperature for 5 min. The cells were washed three times with 0.5% BSA/HBSS and resuspended in 0.5 ml 0.5% BSA/HBSS. Flow cytometry analyses were carried out using a Beckman Coulter FC500 (Fullerton, CA, USA).
Statistical analysis. All values are expressed as means with standard deviations. The effects of different taurolidine and/or PVP concentrations were compared relative to untreated controls (set as 1). Differences between taurolidine concentrations were analyzed using the One-Way ANOVA test. The post-hoc tests Tukey or Games-Howell were applied to correct for multiple testing. A p-value lower than 0.05 was considered statistically significant. All statistical analyses were performed using SPSS software (version 22.0; IBM SPSS, Chicago, IL, USA).
Results
Cell survival. In Figure 1, the cytotoxicity effects of different concentrations of taurolidine and/or PVP at different time intervals on PBMCs and granulocytes is shown. Taurolidine/PVP decreased viability of PMBCs and granulocytes. After two h of incubation, the IC50 concentrations for PBMCs and granulocytes were 500 and 520 μg/ml, respectively (Figure 1A and D). In general, IC50 concentrations were lower for PBMCs than for granulocytes. Prolonged exposure of PBMCs to taurolidine/PVP resulted in decreased IC50 concentrations. After 24 and 48 h, the IC50 concentrations were 40 and 43 μg/ml, respectively. Similar IC50 concentrations were observed for taurolidine dissolved in RPMI (Figure 1B and E). A remarkable increase in PBMC survival was observed for the taurolidine/PVP and PVP dissolved in RPMI, in particular after prolonged exposure, but not for taurolidine dissolved in RPMI.
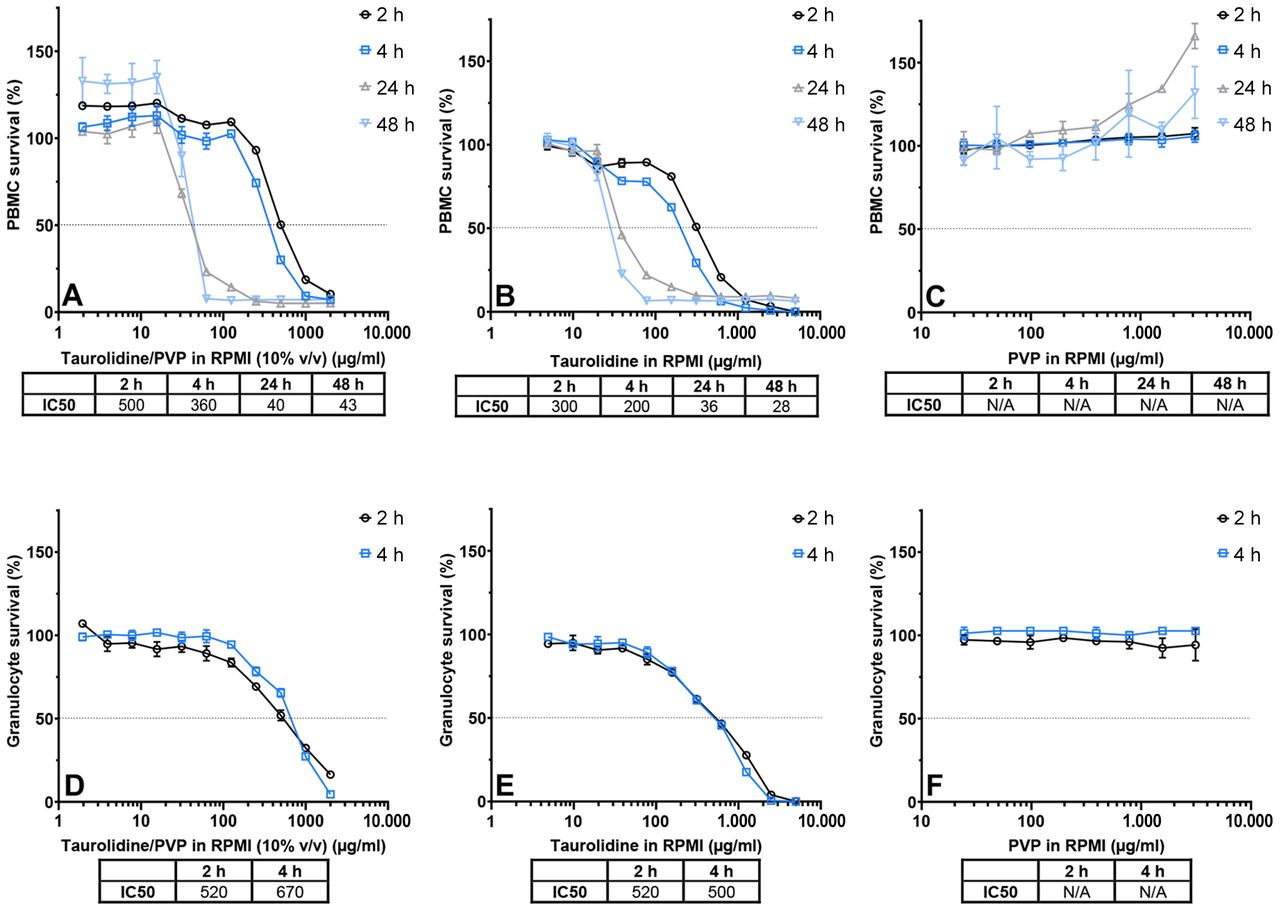
Survival of peripheral blood mononuclear cells (PBMCs) and granulocytes after exposure to various taurolidine and/or polyvinylpyrrolidone (PVP) concentrations. Blood samples from one volunteer were exposed to various taurolidine and/or PVP concentrations. Each measurement was performed in triplicate. Dotted lines indicate 50% survival. A, B, C) PBMC survival after 2, 4, 24, and 48 h incubation with two-fold dilution series of A) Taurolidine/PVP in RPMI (10% v/v), B) Taurolidine in RPMI and C) PVP in RPMI. D, E, F) Granulocyte survival after 2 and 4 h incubation in two-fold dilution series of D) Taurolidine/PVP in RPMI (10% v/v), E) Taurolidine in RPMI and F) PVP in RPMI. IC50: Half maximal inhibitory concentration.
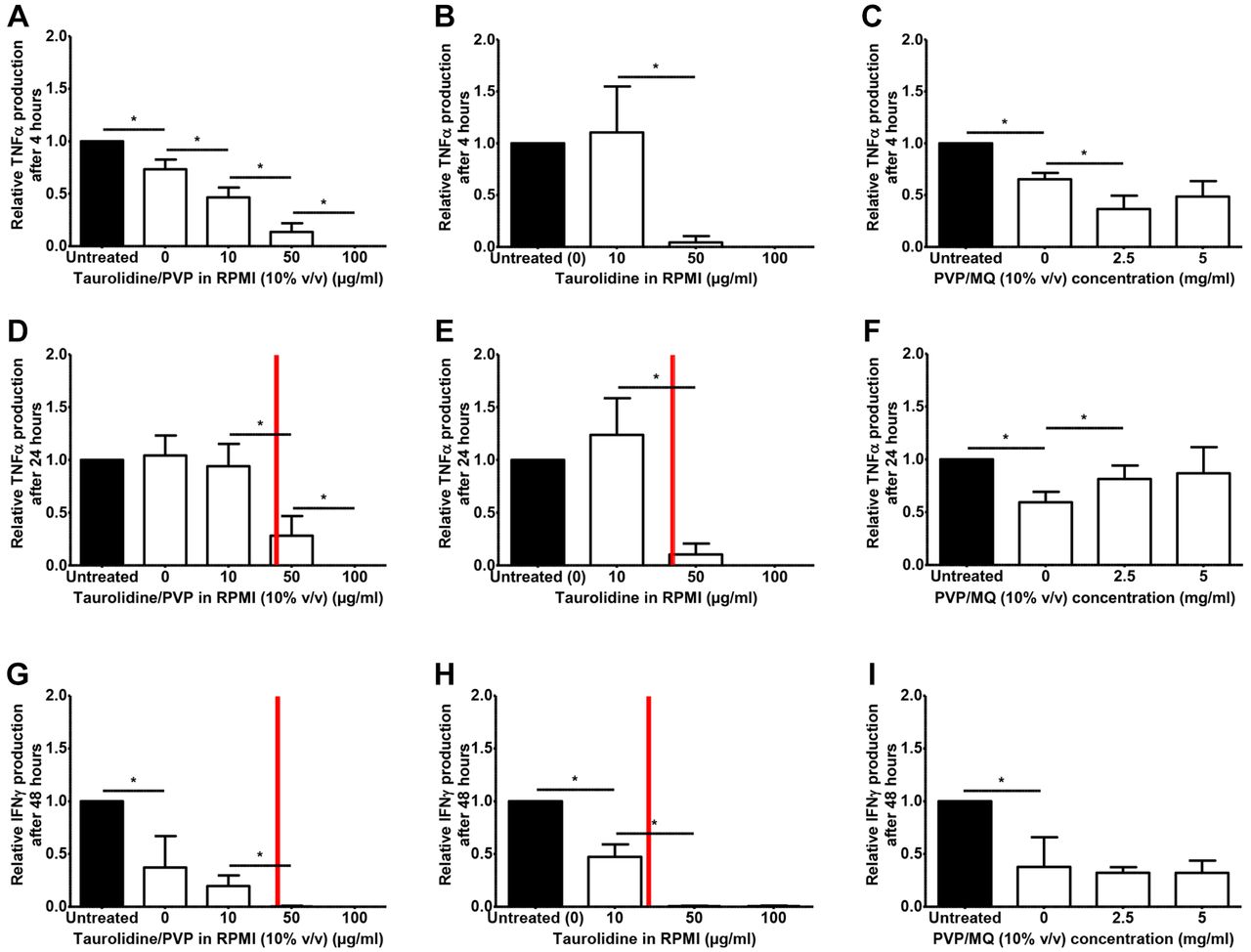
Functions of the adaptive immune defense. Figure 2 demonstrates the relative cytokine production after stimulating PBMCs with LPS (4 and 24 h) or Candida (48 h) at various taurolidine and/or PVP concentrations. Cytokine levels significantly decreased at non-toxic concentrations of taurolidine/PVP (Figure 2A, D, G). After 4 h, TNFα decreased in a dose-dependent pattern. Similar patterns were observed for TNFα and IFNγ at 24 and 48 h of incubation. Notably at these latter time intervals, taurolidine concentrations higher than 40 μg/ml were toxic for PBMCs. Treatment with taurolidine dissolved in RPMI resulted in a similar decrease in TNFα and IFNγ levels (Figure 2B, E and H). The vehicle PVP did not show any relevant effects on cytokine production (Figure 2C, F and I).

Relative cytokine production of peripheral blood mononuclear cells (PBMCs) exposed to taurolidine and/or polyvinylpyrrolidone (PVP). Blood samples from eight volunteers were exposed to various taurolidine and/or PVP concentrations. Each measurement was performed in duplicate. PBMCs were incubated with taurolidine/PVP in RPMI (10% v/v) (A, D, G); taurolidine in RPMI (B, E, H); or PVP/MQ (10% v/v) (C, F, I). PBMCs were stimulated with LPS (A-F) or Candida (G-I), and tumor necrosis factor α (TNFα) (after 4 and 24 h) and interferon-gamma (IFNγ) (after 48 h) were assessed, respectively. Cytokine production for each taurolidine concentration is shown relative to stimulated untreated controls (set at 1). Red bars indicate half maximal inhibitory concentrations (IC50) of taurolidine/PVP or taurolidine, as illustrated in Figure 1. *p<0.05 compared to other taurolidine concentrations or control.
Functions of the innate immune defense. Figure 3 shows the mean ROS production of whole blood for each taurolidine/PVP concentration after stimulation with PMA or STZ. After stimulation with PMA, no change in ROS production was observed at up to 100 μg/ml taurolidine/PVP concentrations. However, treatment with 500 μg/ml taurolidine/PVP decreased the mean ROS production by 19%. After stimulation with STZ, the mean ROS production remained stable at all taurolidine/PVP concentrations. The vehicle PVP did not show any effects on ROS production.

Stimulus-induced reactive oxygen species (ROS) at different taurolidine concentrations in whole blood. Blood samples from twelve volunteers were exposed to taurolidine/polyvinylpyrrolidone (PVP) in RPMI (10% v/v) and stimulated with A) phorbol 12-myristate13-acetate (PMA) or B) serum-treated zymosan (STZ). Each measurement was performed in quintuple. Mean ROS production was expressed as relative light units per granulocyte. The ROS production for each taurolidine/PVP concentration is shown relative to stimulated untreated controls (set at 1). Red bars indicate half maximal inhibitory concentrations (IC50) of taurolidine/PVP or taurolidine, as illustrated in Figure 1. *p<0.05 compared to other taurolidine concentration or control.
In Figure 4, the relative expression of membrane surface markers for adhesion and degranulation are demonstrated. The adhesion markers CD11b and CD62L (the latter of which is downregulated in case of cellular activation) showed an activation trend following treatment of granulocytes with the highest taurolidine/PVP concentration. Similar patterns were observed for the granulocyte-specific degranulation markers CD63 and CD66B.The vehicle PVP did not have an effect on the expression of membrane surface markers.

Relative expression of cell membrane markers on granulocytes (CD11b, CD62L, CD63, CD66B). Graphs show the mean activation of cell membrane markers from whole blood of eight volunteers exposed to 0, 100, or 500 μg/ml taurolidine/polyvinylpyrrolidone (PVP) in RPMI (10% v/v). The expression of membrane surface antigens in viable granulocytes at each taurolidine concentration is shown relative to stimulated untreated controls (set at 1). A) activation of CD11b, B) activation of CD62L, which becomes downregulated during cell activation due to receptor shedding, C) activation of CD63, and D) activation of CD66B. *p<0.05 compared to other taurolidine/PVP concentration or control.
Discussion
Since the 1970s, taurolidine has emerged in several fields because of its therapeutic potential in various clinical settings. However, there has been limited insight on taurolidine’s antimicrobial, immunoregulatory and antineoplastic characteristics. Nonetheless, the successful prevention of infections and the treatment of neoplasms with taurolidine requires a deep knowledge of the mechanisms that underlie the intricate trade-off between antimicrobial/antineoplastic effects on one hand, and subtle interactions with the host immune system on the other.
In the present study, we investigated how crucial PBMC and granulocyte functions are modulated by a broad concentration range of taurolidine. As a most important issue that was lacking in previous studies, cell viability was assessed to rule out that any cytotoxic effects of taurolidine underlie these effects. We found that taurolidine, depending on its concentration, is cytotoxic in both PBMCs and granulocytes, which seems not surprising given its antiseptic and antineoplastic properties. Indeed, several studies have reported on taurolidine’s effect to inhibit microbial growth, with minimum inhibitory concentrations (MICs), ranging between 256 to 2,048 μg/ml. Other studies have shown dose response effects in numerous cultured (24-72 h) tumor cell lines, with half maximal effective concentrations ranging between 40 to 80 μg/ml (20, 21). So far, however, adequate studies on the cytotoxic effects of various taurolidine concentrations in PBMCs and granulocytes were lacking. Bedrosian et al. investigated viability in PBMCs after 24 h incubation, but did not find any evidence for cytotoxic effects up to 100 μg/ml (12). Their use of lactic acid dehydrogenase release, a known non-sensitive measure of cytotoxicity, is a likely explanation for the apparent conflict with our findings.
PBMCs appeared to be more sensitive to taurolidine when compared to granulocytes, and cell viability decreased in both cell types during longer treatment periods, i.e., MICs decreased, and immune cells appeared to be increasingly sensitive to this compound after longer exposure. These results seem of importance in clinical practice when considering new treatment strategies.
Stendel et al. investigated the pharmacokinetics of 2% taurolidine following repeated intravenous infusions in glioblastoma patients (22). The authors found a mean plasma concentration of 83 μg/ml and showed that the treatment regimen used in their study was within the antineoplastic effective concentration range. Treatment regimens differ, however, per tumor type or setting. For example, taurolidine has been infused in large volumes (>1 liter per 12 h) in cancer treatment, and only a few milliliters for the prevention of catheter-related bloodstream infections (22-24). Given taurolidine’s limited solubility of up to 2% in water and its low mean plasma concentration due to its short half-life of 1.7 h despite high dose intravenous volumes in vivo, we expect that IC50 concentrations of 500 μg/ml found after short incubation may be less relevant in clinical practice (22).
After 4, 24 and 48 h incubation with taurolidine/PVP, a dose response effect was observed for TNFα and IFNγ production. The dose response pattern of TNFα following incubation for 24 h is in accordance with Bedrosian et al. (12) However, it is again important to note that the toxicity of taurolidine/PVP is time dependent and increases during longer incubation periods. Therefore, for the 24- and 48 h time intervals, the toxic concentrations of 50 and 100 μg/ml probably decreased the production of TNFα and IFNγ. To date, the exact underlying mechanisms by which taurolidine induces its anti-inflammatory response remains elusive. Some studies have proposed that taurolidine has antitoxin properties. According to this hypothesis, taurolidine neutralizes cytokine inducing agents, such as LPS, by the release of active methylol groups, which cross-link the amino groups of these (endo)toxins (25, 26). Bedrosian et al., however, found that the reduction of cytokine production was due to taurolidine but not to a reduction of LPS by taurolidine. In addition, taurolidine in combination with non-toxin-inducing agents, such as heat-killed S. epidermidis or C. albicans, as was observed in this study, have been also shown to reduce cytokine production (12). This suggests that taurolidine reduces cytokine production independent of its antitoxin property. Taurolidine’s mode of action possibly involves mechanisms independent of the presence of methylol groups. According to this hypothesis, suppression takes place at the level of the translation of cytokines or other components of protein kinase C-dependent signal transduction pathways in affected immune cells (1, 12, 27).
ROS production remained unaffected by taurolidine at <500 μg/ml concentrations for PMA and STZ. A higher concentration of 500 μg/ml resulted in a significant decrease in ROS production after stimulation with PMA, but not after stimulation with STZ. This may be related to the different modes of action of PMA and STZ. Whereas PMA can influence ROS production directly by stimulating protein kinase-C within the cell, STZ acts in a receptor-dependent, and –hence– slower, manner. In addition, the cytotoxic taurolidine concentration of 500 μg/ml might have influenced the ROS production as well. Several studies have reported various effects of taurolidine on ROS production in different settings. Van Westreenen et al. observed similar results and found a 50% decrease in ROS production in granulocytes treated with 500 μg/ml taurolidine after stimulation with PMA (13). However, Watson et al. found an increase in superoxide anion release and nitric oxide generation in peritoneal macrophages from CD1 mice (1). In addition, several studies have reported increased levels of ROS in various tumor cell types (20).
Treatment with 500 μg/ml taurolidine increased the expression of all evaluated cell membrane markers. Whereas the increased expression of activity cell markers might indicate that taurolidine primes granulocytes to exhibit various defense functions, it must be kept in mind that, although all markers were tested in viable cells, stress due to toxic taurolidine concentrations (500 μg/ml) may have played a role. Whether the observed cell activation enables a more potent response to invading pathogens in vivo remains a question.
Taurolidine’s vehicle PVP is used in various taurolidine solutions in clinical practice and has been reported to stabilize decomposition products of taurolidine, such as its primary degrading product taurultam (28). Indeed, taurolidine’s metabolites were stable over prolonged incubation periods (>48 h) in both whole blood and PVP with RPMI or PBS (data not shown). In the present study, we investigated PVP’s role as it may have potential side effects on immune cell functions and cell viability. PVP did not have any relevant effects on cytokine production in PBMCs, or ROS production and expression of cell membrane markers in granulocytes, but a remarkable finding was the increase in PBMC viability at higher PVP concentrations in RPMI. At first glance, these results suggest a proliferative effect of PVP on PBMCs. However, PBMCs do not proliferate ex vivo. After incubation of PVP in RPMI with WST-1, in the absence of PBMCs, a similar increase in absorbance was measured for higher PVP concentrations, suggesting an interaction of PVP and WST-1, rather than an effect on cell proliferation (data not shown).
The strengths of this study include its broad experimental set-up in which we explored the effect of a broad range of taurolidine concentrations on several relevant human innate and adaptive immune cell functions and related these to the effects of the vehicle PVP. Furthermore, we included both prolonged incubation periods (24 and 48 h) to compare our results with the current literature, and short incubation periods (2 and 4 h) as this may be more relevant in clinical practice given taurolidine’s rapid clearance (22).
The implementation of the higher cytotoxic taurolidine concentration (500 μg/ml), although methodologically valid, obviously limited the range of our conclusions in some of the experiments. Another limitation of this study is its ex vivo design, as the results may not (entirely) apply in vivo, and the relevance for clinical practice needs to be further established. In addition, since we did not investigate taurolidine’s mechanism of action to decrease cytokine production, additional studies need to be conducted on the specific intracellular (synthesis) pathways that are affected by taurolidine.
In conclusion, this study showed that taurolidine displays dose- and time-dependent cytotoxicity toward PBMCs and granulocytes ex vivo, and that the sensitivity to taurolidine can be observed at lower concentrations in PBMCs compared to granulocytes.
Footnotes
Authors’ Contributions
YW, GM, RTM, HR, GW designed the study: conceptualization, and methodology. YW, GM, and HR gathered data: resources, and data curation. YW and GM analyzed the data and wrote the manuscript: formal analysis, investigation, visualization, and writing – original draft; which was reviewed and approved by all authors: writing – review & editing. All Authors jointly decided to publish.
Conflicts of Interest
GW reports grants from Geistlich Pharma AG, Fresenius Kabi, Baxter international and BBraun Medical outside the submitted work. GW is a consultant for Shire. YW reports grants from Geistlich Pharma AG outside the submitted work.
- Received June 2, 2022.
- Revision received July 8, 2022.
- Accepted July 12, 2022.
- Copyright © 2022, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY-NC-ND) 4.0 international license (https://creativecommons.org/licenses/by-nc-nd/4.0).
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.