


Abstract
Background/Aim: At present, there are no effective drugs for the treatment of insulin resistance. MTH-3, a curcumin derivative, exerts potent anti-cancer effects. The aim of the present study was to explore whether MTH-3 is capable of regulating palmitic acid (PA)-induced insulin resistance in C2C12 cells. Materials and Methods: Cell viability was examined using the MTT assay. C2C12 cells were treated with PA and evaluated for the production of oil droplets using an Oil Red O assay. Glucose uptake was analysed by the 2-NBDG assay. Results: Treatment of cells with up to 500 μM PA for 24 h or with 5 or 10 μM MTH-3 had no effect on cell viability. PA induced production of oil droplets in C2C12 cells. After adding MTH-3, the quantity of oil droplets decreased significantly and glucose uptake recovered. Conclusion: MTH-3 may become an efficient drug for the treatment of insulin resistance and associated diseases.
As the prevalence rate of obesity is increasing on a yearly basis, the incidence of obesity-related complications has also risen sharply. Obesity is regarded as a chronic disease and requires long-term observation and management (1). Thus, there is an urgent need to study potential treatments for obesity-related diseases. Obesity refers to a physical condition that has a negative impact on health due to an excessive increase and accumulation in body fat. The primary causes of obesity are excessive energy intake and a sedentary lifestyle; in a small number of cases, the cause is genetic and/or psychological or due to physical diseases. The current treatment methods are behavioral therapy, drug treatment, and bariatric surgery. Behavior therapy is currently the most important treatment method, including diet control and increased exercise (2). The effects of obesity can be roughly divided into two groups. The first effect of obesity is the mental illness/awareness it can cause due to social obstacles/perceptions (3). The second effect of obesity is the metabolic changes including increased risk of diabetes, insulin resistance, metabolic syndrome, gallbladder disease, hypertension, cardiovascular disease, and cancer (4).
Insulin serves a role in regulating glucose metabolism in tissues such as the fat, liver, heart, kidney and in the central nervous system. It can also inhibit glucose production and increase the speed of glucose absorption by muscle and fat cells. Insulin resistance refers to a situation in which liver cells, fat cells, and muscle cells do not respond to normal concentrations of insulin, thus requiring increased production of insulin (5). In individuals with insulin resistance, a physiological concentration of insulin cannot induce absorption of glucose by muscle and fat cells. To compensate for this situation, the pancreas secretes increased quantities of insulin to stimulate these cells to absorb glucose, which results in hyperinsulinemia. Excessive levels of insulin in the blood triggers obesity, as insulin binds receptors on fat cells to inhibit the activity of lipase, which would normally breakdown triglycerides, resulting in the formation of larger fat cells (6).
Palmitic acid (PA) is one of the most commonly found fatty acids in animals and plants. Butter, cheese, milk, and meat all possess this fatty acid, which is the only fatty acid synthesized in the human body. The liver, adipose tissue, and small intestines in the human body are the primary organs involved in PA synthesis, of which the liver possesses the greatest capacity. Studies have shown that excess lipids affect the levels of free fatty acids, and causes insulin resistance in muscle and liver cells (7, 8). PA is one of the most abundant free fatty acids, accounting for 30-35% of the total free fatty acids in the human blood (9). PA affects the delivery of insulin to hepatocytes and muscle cells in culture (10, 11).
Curcumin is extracted and purified from the root of turmeric. In the past, it was often used in traditional pharmacology to treat biliary diseases, loss of appetite, and cough. Curcumin possesses anti-inflammatory, anti-oxidant, and anti-tumor activities (12-14). Studies have shown that curcumin significantly reduces blood sugar and blood lipid levels in diabetic rats (15, 16), and significantly improves the symptoms of disordered fat metabolism in diabetic rats (17). Although curcumin has been shown to possess a favorable therapeutic effect on several diseases (18), its shortcomings are the poor hydrophilicity and low bioavailability. The curcumin derivative, MTH-3, has thus been developed and studied with the aim of improving hydrophilicity (19). MTH-3 has also been shown to be more effective with regards to anti-cancer effects against triple-negative breast cancer in animal experiments compared with curcumin (20). However, the mechanism by which MTH-3 regulates insulin resistance caused by free fatty acid metabolism remains unclear. The aim of the present study was to explore the effects and mechanism of MTH-3 on insulin resistance in C2C12 cells.
Materials and Methods
Cell differentiation. Cell differentiation was assayed as described previously by Bajaj et al. (21). Briefly, cells were plated at a density of 2×105 cells/ml in each well of a 6-well plate for 24 h. Cells were cultured in DMEM supplemented with 2% horse serum, and the medium was changed every day for 5 days. The formation of a muscular tube represented the completion of differentiation.
MTT assay. Cell viability was assessed using an MTT assay, as previously described (22). Briefly, cells were seeded at a density of 2×105 cells/ml in each well of a 96-well plate (Thermo Fisher Scientific, Inc., Waltham, MA, USA), cultured for 24 h, and then treated with different concentrations (0, 5 and 10 μM) of MTH-3 or different concentrations of PA (0, 100, 200, 300, 400 or 500 μM) for 24 h. MTT was added to each well at a final concentration of 0.5 mg/ml, and incubated at 37°C for 3 h. The supernatant was discarded and DMSO was added to dissolve the formazan crystals. The absorbance at 595 nm was recorded using a microplate reader, and control absorbance was normalized to 100%. A total of six wells per condition were used, across at least four independent experiments.
Oil Red O staining. Cells were cultured in DMEM containing 2% horse serum, and medium was changed every day for 5 days. After 5 days, the culture medium was aspirated, and different concentrations (0, 100, 200, 300, 400 or 500 μM) of PA were added for 24 h. Cells were washed with PBS and then 1 ml of 10% formalin was added to each well for 1 h. Cells were washed with ddH2O and then 1 ml 60% isopropanol was added to each well. After isopropanol was aspirated, cells were stained with Oil-Red-O at room temperature for 2 h in the dark and then observed using an inverted microscope at 100× magnification (23).
Glucose uptake assay. The C2C12 myotubes were treated with 0.4 mM PA and MTH-3 (5 and 10 μM) simultaneously for 24 h. The cells were washed three times with PBS, and subsequently incubated in the presence or absence of 100 nM insulin for 10 min before the addition of 20 μl 2-NBDG (5 mM) for 2 h at 37°C. Subsequently, 30 μl cell suspension was immediately loaded into a 2-chamber slide (NC-Slide A2; ChemoMetec A/S, Allerod, Denmark) for analysis on the NucleoCounter NC-3000 using the built-in caspase assay program (24).
RNA extract. Total RNA was isolated using TRIzol® reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to the manufacturer’s protocol, as described previously (25). Briefly, 2×106 cells for each treatment were harvested and mixed with 1 ml TRIzol® reagent at room temperature for 5 min. To each tube, 0.2 ml chloroform was added, and centrifuged at 12,000×g for 15 min at 4°C. The supernatant was collected and mixed with 0.5 ml isopropanol for 10 min. This was further centrifuged at 12,000×g for 15 min at 4°C. RNA pellets were dissolved in 100 μl DEPC-treated H2O.
Quantitative (q)PCR. qPCR was performed using a TaKaRa One Step SYBR Ex Taq™ qRT-PCR kit (Takara Bio, Inc., Shiga, Japan) as described previously (25) in a QuantStudio™ 3 Real-Time PCR system (Thermo Fisher Scientific, Inc.). Relative expression levels of target genes were normalized to β-actin. The relative expression levels of mRNA are presented as the mean±standard deviation of duplicates. The sequences of the primers used for qPCR were: GAPDH, forward, 5’-gagtgtttcctcgtcccgtaga-3’ and reverse, 5’-gtgaccaggcgcccaata-3’; glucose transporter 4 (GLUT4) forward, 5’-caatggttgggaaggaaaagg-3’ and reverse, 5’-ggcgccccccaaca-3’; Myogenin forward, 5’-accctacagacgcccacaatc-3’ and reverse, 5’-acatatcctccaccgtgatgct-3’; transcription factor EB (TFEB) forward, 5’-gcggcagaagaaagacaatcac-3’ and reverse, 5’-tgatccggtcattgatgttgaa-3’; peroxisome proliferator-activated receptor-γ (PPAR-γ) coactivator 1-α (PGC1α) forward, 5’-ggcacgcagccctattca-3’ and reverse, 5’-tctcgacacggagagttaaagga-3’; and PPARγ forward, 5’-accacagttgatttctccagcatt-3’ and reverse, 5’-ccattgggtcagctcttgtga-3’.
Western blotting. Western blotting was performed as described previously (26). Briefly, 100 μg protein prepared form each sample was used for western blot analysis. The primary antibodies used were: Anti-GLUT4 (cat. no. GTX79317; GeneTex Inc., Irvine, CA, USA) and anti-β-Actin (cat. no. GTX629630; GeneTex Inc.). Protein bands were semi-quantified using ImageJ 1.4 (National Institutes of Health, Bethesda, MD, USA).
Statistical analysis. The experimental data are presented as the mean±standard deviation. A one-way ANOVA was used for multiple comparisons. p<0.05 was considered to indicate a statistically significant difference. Data were analyzed using GraphPad Prism version 7 (GraphPad Software, Inc., San Diego, CA, USA).
Results
Effect of PA on C2C12 cell survival. To establish a cell model for evaluating C2C12 differentiation, C2C12 cells were cultured in a differentiation medium. C2C12 cells differentiated after cells were cultured for 5 days in the differentiation medium (Figure 1A). Myotubes were formed that were observed under an inverted microscope. Myogenin expression, a biological indicator of C2C12 differentiation, was evaluated by reverse transcription (RT)-qPCR (Figure 1B). Myogenin expression was significantly increased after cells were cultured for 5 days in the differentiation medium. To examine whether PA affected cell viability in C2C12 cells, an MTT assay was performed. C2C12 myotubes cells were treated with 0-500 μM PA (Figure 2A) for 24 h to examine cell viability (Figure 2B). Cell viability was decreased when cells were treated with 500 μM PA. PA at a concentration of 100-400 μM had no effect on C2C12 myotube cells. Therefore, PA concentrations <400 μM were used in the present study.
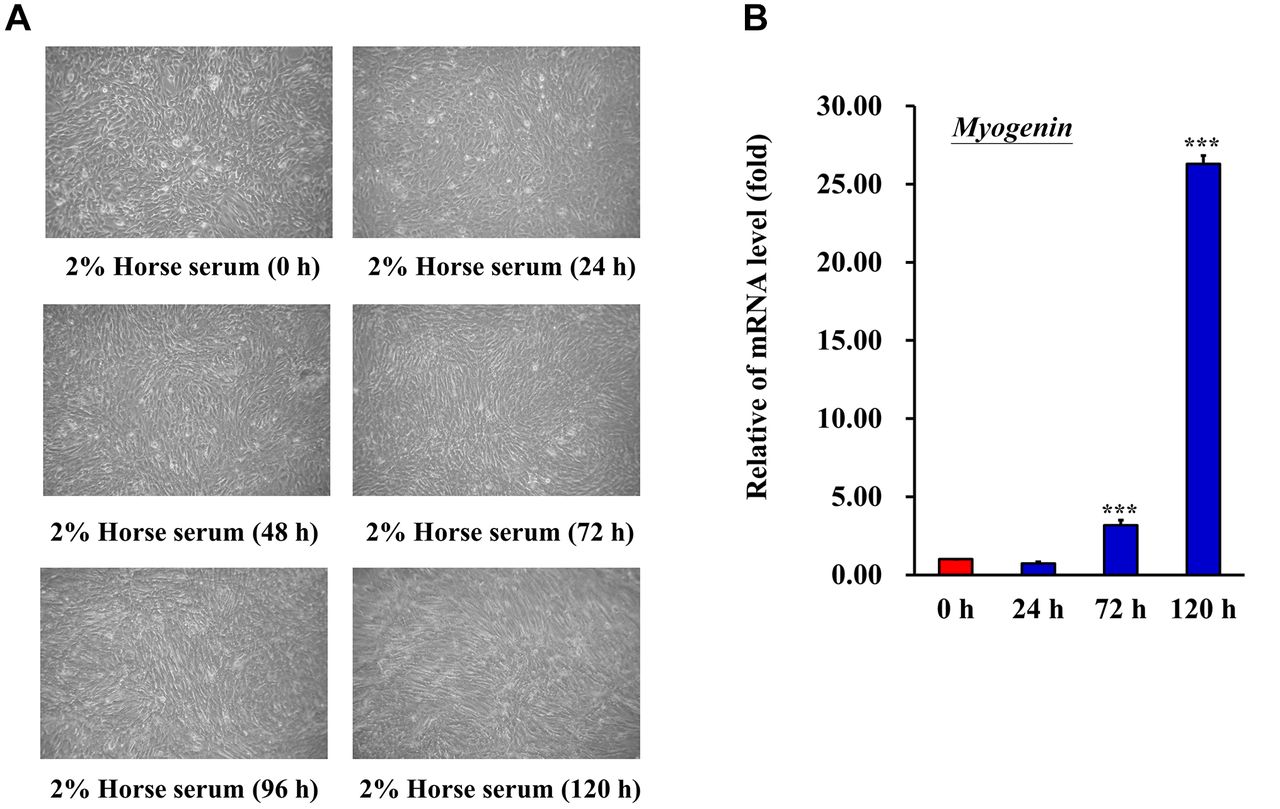
Differentiation of C2C12 cells. (A) Changes in morphology of C2C12 cells. C2C12 were cultured in DMEM supplemented with 2% horse serum to induce differentiation. Culture medium was changed daily. Formation of myotubes was observed under an inverted microscope daily. (B) mRNA expression of myogenin, a biological indicator of C2C12 differentiation, was evaluated. ***p<0.001 vs. control.

Effect of palmitic acid (PA) on C2C12 cell survival. (A) Chemical structure of PA. (B) C2C12 cells were treated with 0, 100, 200, 300, 400, or 500 μM PA for 24 h. Cell viability was determined using an MTT assay. ***p<0.001 vs. control. PA: Palmitic acid.
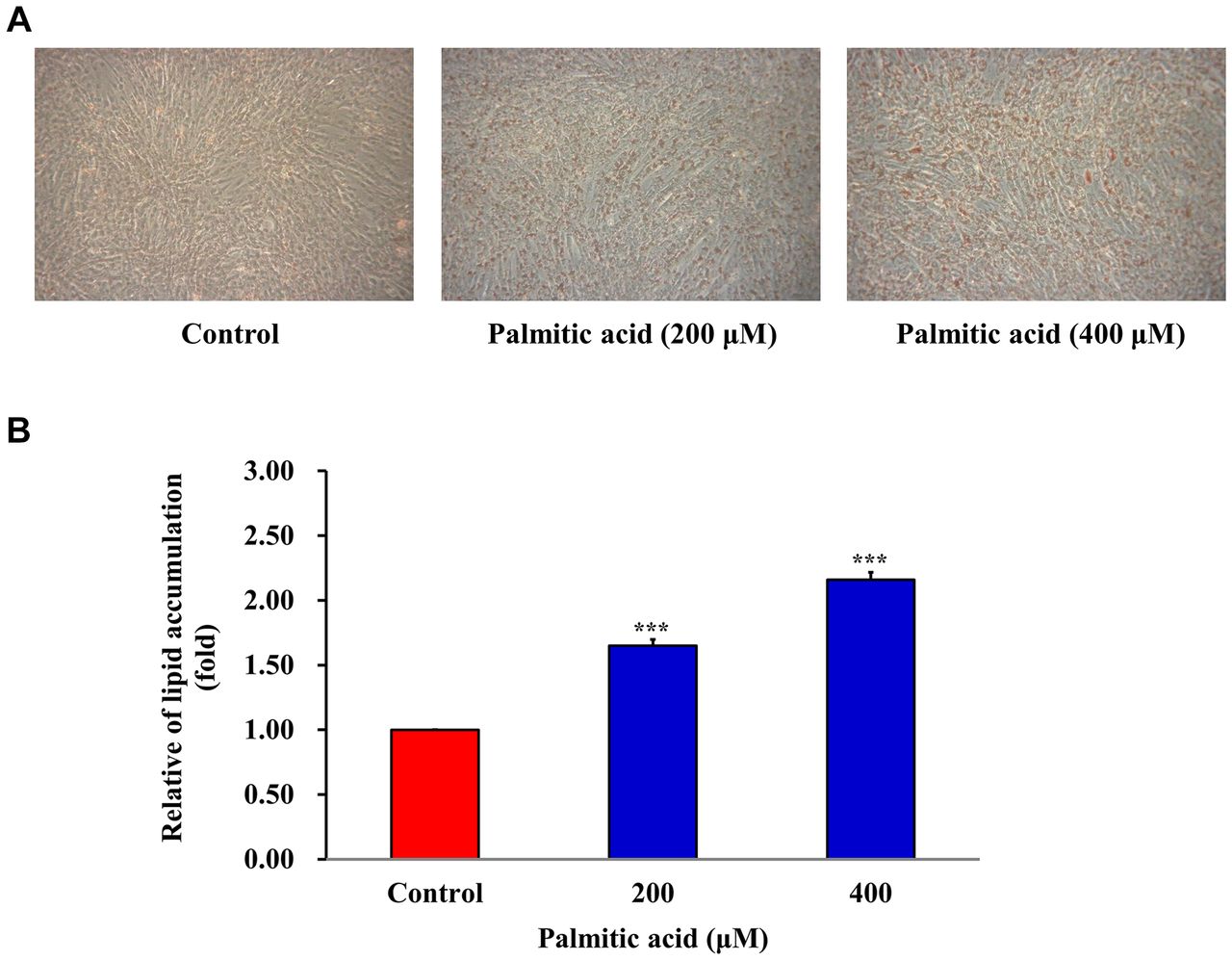
PA induces the production of oil droplets in C2C12 cells. Lipid droplets contain neutral lipids, primarily triglycerides, and cholesterol esters. Excessive accumulation of lipid droplets in cells may be an indicator of metabolic defects or pathogenesis. Oil Red O binds to triglycerides, lipids and lipoproteins, and dyes them red. The quantity of oil droplets was determined using Oil Red O. C2C12 cells were treated with 200 and 400 μM PA for 24 h. PA treated C2C12 cells produced more oil droplets than the control group (Figure 3A and B).

Palmitic acid (PA) induces the production of oil droplets in C2C12 cells. (A) C2C12 cells were treated with 0, 200 or 400 μM PA for 24 h. Production of oil droplets was assayed using Oil Red O staining. (B) Fold change in the treated group compared with the control group was calculated. ***p<0.001 vs. control.
PA down-regulates GLUT4 expression in C2C12 cells. The primary glucose absorption receptor in muscle cells is GLUT4. To examine whether PA affected the function of GLUT4 to alter insulin resistance, the protein and mRNA expression levels of GLUT4 were analyzed using western blotting and RT-qPCR, respectively. The protein and mRNA expression levels of GLUT4 were significantly reduced in PA-treated C2C12 cells (Figure 4A and B). These results highlight a potential mechanism for the insulin resistance in PA-treated C2C12 cells.
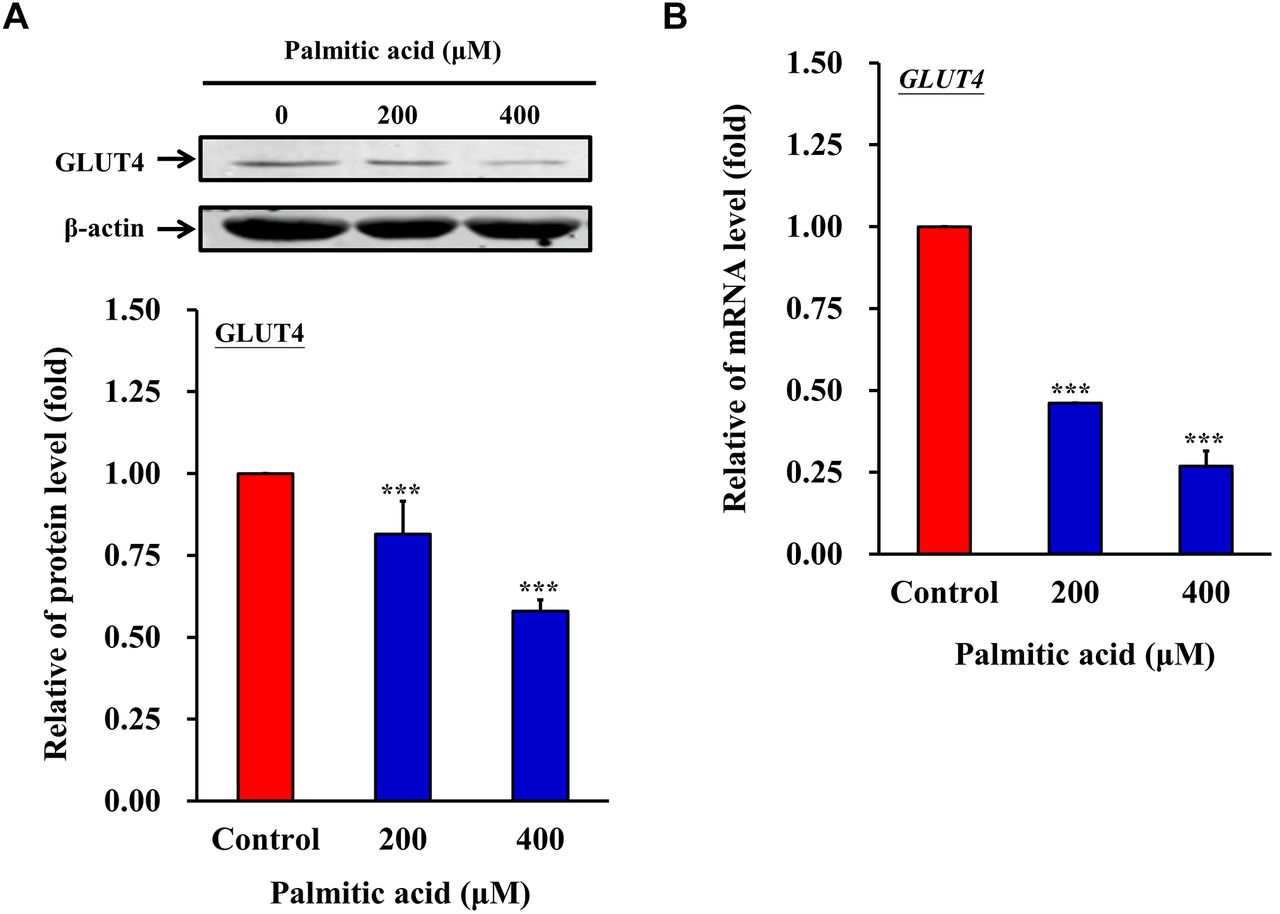
PA down-regulates GLUT4 expression in C2C12 cells. C2C12 cells were treated with 0, 200 or 400 μM PA for 24 h. (A) Protein and (B) RNA expression levels of GLUT4 were analyzed by western blotting and reverse transcription-quantitative PCR, respectively. ***p<0.001 vs. control. GLUT4: Glucose transporter 4.
MTH-3 reduces the effect of PA on the production of oil droplets in C2C12 cells. C2C12 myotubes cells were treated with 5 or 10 μM MTH-3 (Figure 5A) for 24 h to examine cell viability using an MTT assay (Figure 5B). MTH-3 at a concentration of 5 or 10 μM had no effect on C2C12 cells. To investigate whether MTH-3 reduced the production of oil droplets in PA-stimulated C2C12 cells, Oil Red O staining was performed. C2C12 cells were treated with MTH-3 (0, 5 or 10 μM) in the absence or presence of PA (400 μM) for 24 h. The results showed that MTH-3 reduced the production of oil droplets in PA-treated C2C12 cells (Figure 6).

Effect of MTH-3 on C2C12 cell survival. (A) Chemical structure of MTH-3. (B) C2C12 cells were treated with 0, 5 or 10 μM MTH-3 for 24 h. Cell viability was determined using an MTT assay. MTH-3 at a concentration of 5 and 10 μM had no effect on the viability of C2C12 cells.

MTH-3 reduces the effect of palmitic acid (PA) on the production of oil droplets in C2C12 cells. C2C12 cells were treated with MTH-3 (0, 5 and 10 μM) in the absence or presence of PA (400 μM) for 24 h. The production of oil droplets was assayed using Oil Red O staining. Fold change was calculated in the treated groups compared with the control group. ***p<0.001 vs. untreated group.
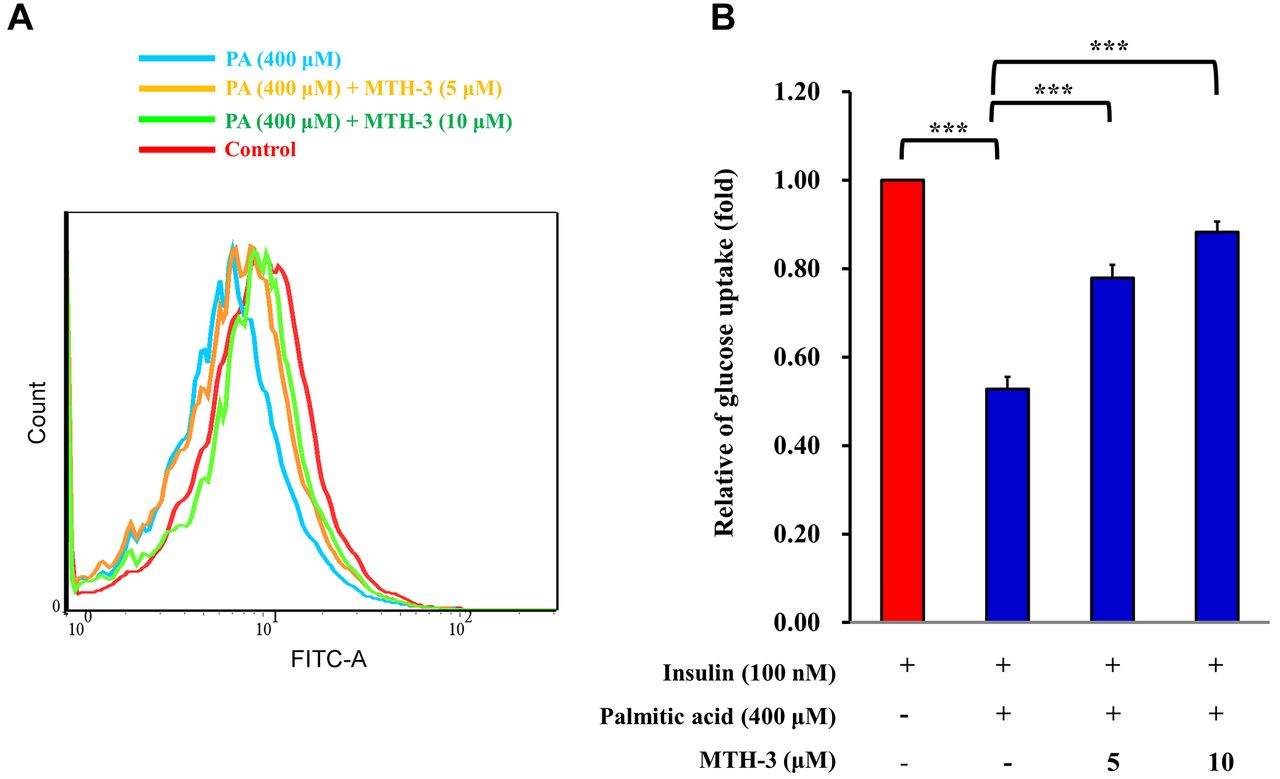
MTH-3 ameliorates glucose uptake in PA-treated C2C12 cells. To investigate whether MTH-3 caused insulin resistance, 2-NBDG was used to detect glucose uptake in PA-stimulated C2C12 cells. C2C12 cells were pre-treated with 100 nM insulin for 10 min, and subsequently treated with MTH-3 (0, 5 or 10 μM) in the absence or presence of PA (400 μM) for 24 h. 2-NBDG was added to cells to analyze glucose uptake. As shown in Figure 7, glucose uptake was significantly increased in the MTH-3 treated C2C12 cells.

MTH-3 ameliorates glucose uptake in palmitic acid (PA)-treated C2C12 cells. (A) C2C12 cells were pre-treated with 100 nM insulin for 10 min. C2C12 cells were subsequently treated with MTH-3 (0, 5 and 10 μM) in the absence or presence of PA (400 μM) for 24 h. 2-NBDG was added to cells to assess glucose uptake. (B) Relative changes were calculated in the treated group compared with the untreated group. ***p<0.001 vs. untreated group.
MTH-3 reverses the expression of TFEB and PPAR-γ in PA-treated C2C12 cells. To characterize the signaling pathways involved, the expression of TFEB and PPAR-γ were determined. TFEB is the primary transcription factor in lysosomal biosynthesis. In a state of starvation, TFEB increases autophagy, and thus increases fat metabolism. In the liver, TFEB regulates genes involved in lipid catabolism, fatty acid oxidation, and ketone synthesis (27). TFEB also plays a key role in mediating insulin sensitivity, glucose stabilization, lipid oxidation, and mitochondrial function (28). C2C12 cells were treated with 5 and 10 μM MTH-3 in the absence or presence of PA (400 μM) for 24 h, and RNA was subsequently extracted. Expression of TFEB was determined by RT-qPCR. TFEB expression was significantly increased in the MTH-3-treated C2C12 cells (Figure 8A). PPARs are a group of nuclear proteins that function as transcription factors to regulate gene expression. PPARs serve a key role in cell differentiation, metabolism and cancer (29). There are three types of PPARs, namely PPARα, β and γ. These three types are expressed in different tissues and reflect different physiological functions. Amongst these, PPAR-γ plays an important regulatory role in insulin sensitivity (30). C2C12 cells were treated with 5 and 10 μM MTH-3 in the absence or presence of PA (400 μM) for 24 h and RNA was subsequently extracted. Expression of PPAR-γ was determined by RT-qPCR. PPAR-γ expression was significantly increased in the MTH-3-treated C2C12 cells (Figure 8B). Taken together, TFEB and PPAR-γ expression was significantly increased in the MTH-3-treated C2C12 cells.
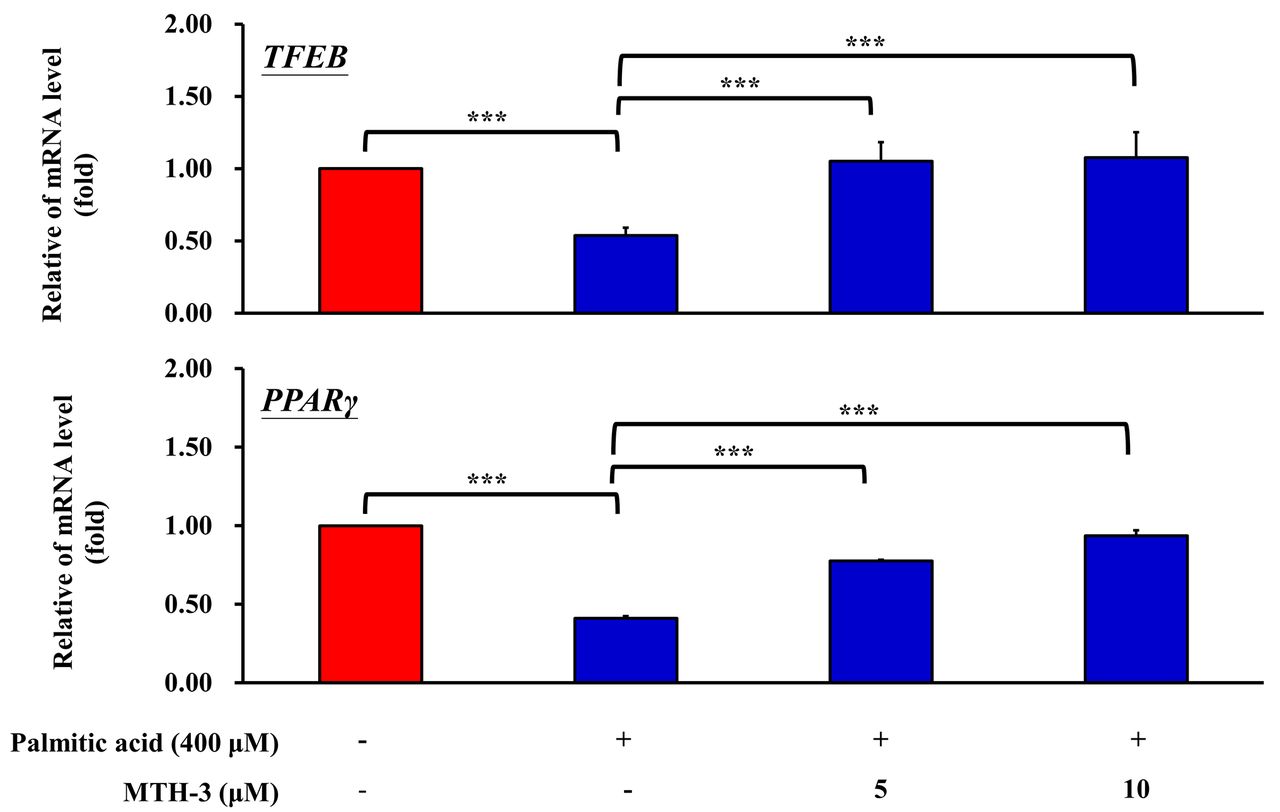
MTH-3 decreases the expression of TFEB and PPAR-γ in PA-treated C2C12 cells. C2C12 cells were treated with MTH-3 (0, 5 and 10 μM) in the absence or presence of PA (400 μM) for 24 h, and RNA was subsequently extracted. Expression of (A) TFEB and (B) PPAR-γ was determined by reverse transcription-quantitative PCR. Relative changes were calculated in the treated group compared with the untreated group. ***p<0.001 vs. control. TFEB: Transcription factor EB; PPARγ: peroxisome proliferator-activated receptor-γ; PA: palmitic acid.
Discussion
Skeletal myoblasts are the optimal cells for studying muscle cell proliferation and differentiation in vitro (31, 32). When skeletal myoblasts are cultured in medium supplemented with 10% fetal calf serum, they continue to proliferate. However, when they are cultured in medium supplemented with 2% horse serum, the cells readily fuse and form myotubes (31, 33). In the present study, the C2C12 cells were an immortalized mouse myoblast cell line, and a well-established in vitro model for studying muscle regeneration and differentiation. In the present study, C2C12 cells were used to study myoblast proliferation and differentiation. During C2C12 cell differentiation, myoblasts undergo remodeling to form mature myotubes, and exhibit increased myogenin mRNA expression (34, 35). As shown in Figure 1A, 2% horse serum medium triggered myogenic differentiation after 72, 96, and 120 h of induction, and the cells that formed the myotubes presented considerable numbers of fused myotubes. Myogenin expression at the mRNA level was increased after 72 and 120 h of culture with 2% horse serum. Thus, it was demonstrated that 2% horse serum induced myogenic differentiation of C2C12 cells.
Insulin resistance is one of the major physiological characteristics of a type II diabetes mellitus (T2D) phenotype (36-38). High levels of free fatty acids in the plasma are associated with insulin resistance in T2D (39, 40). PA is a saturated fatty acid that has been implicated in the development of T2D (41). Excess transpiration of PA to the skeletal muscle is implicated in the development of insulin resistance, resulting from lipid oversupply to skeletal muscles (42-44). In the present study, 100, 200, 300, 400, and 500 μM PA was used to treat C2C12 cells cultured with 2% horse serum supplemented medium for up to 120 h. Treatment for 24 h with 100-400 μM PA did not affect cell viability. PA-induced intracellular lipid accumulation as determined using Oil Red O staining when cells were treated with 200 and 400 μM PA. It has been reported that GLUT4 protein is highly expressed in C2C12 cells, which can be used in the investigation of glucose uptake and insulin resistance mechanisms (45, 46). In the present study, it was shown that GLUT4 mRNA and protein expression levels were reduced in cells treated with 200 and 400 μM PA. It was hypothesized that PA induced intracellular lipid droplet deposition and increased insulin resistance in C2C12 myotubes. Thus, an in vitro model of insulin resistance using C2C12 myotubes was successfully established.
Curcumin, a constituent compound of Curcuma longa plants, has various pharmacological properties, including anti-oxidant, anti-inflammatory, anti-bacterial, anti-viral, anti-cancer, and anti-diabetic properties (47, 48). A clinical study showed that curcumin was safe even at doses of up to 8 g per day (20). However, the pharmacokinetic studies of curcumin on animal and human models have demonstrated poor absorption in the gastrointestinal tract (49, 50). Low systemic bioavailability of curcumin prevents a suitable concentration reaching the target tissues to achieve its pharmacological effects (20). To overcome its poor bioavailability and increase curcumin absorption in the gastrointestinal tract, a novel curcumin derivative, (1E,3Z,6E)-3-hydroxy-5-oxohepta-1,3,6-triene-1,7-diyl)-bis(2-methoxy-4,1-phenylene)-bis(3-hydroxy-2-hydroxymethyl)-2-methyl propanoate, referred to as MTH-3 in the present study, was established. A previous study demonstrated that MTH-3 inhibits proliferation of triple-negative breast adenocarcinoma cells and induces cell death in vitro and in vivo (20). In our previous study, it was also demonstrated that MTH-3 inhibits triple-negative breast adenocarcinoma cell metastasis via the MAPK/ERK/AKT signaling pathway (51). MTH-3 was effective against several different cancer cell lines and exerted only limited toxicity on physiologically healthy cells (19). To the best of our knowledge, the present study is the first to demonstrate that MTH-3 regulates PA-induced insulin resistance in mouse myoblast C2C12 cells. The results showed that 5 and 10 μM MTH-3 treatment for 24 h did not affect cell viability. MTH-3 reduced intracellular lipid accumulation and glucose uptake when 5 and 10 μM MTH-3 was administered in C2C12 cells treated with PA to induce insulin resistance.
TFEB is a transcription factor, and acts as a primary regulator of autophagy, lysosomal genesis, lipid and energy metabolism, and the immune response (52-55). Previous studies demonstrated that TFEB can advance neurodegenerative diseases, such as Parkinson’s disease and Huntington’s disease (56, 57). TFEB also serves an important role in the metabolic response to starvation (58-60). Absence of TFEB results in impairment of lipid catabolism, and TFEB over-expression rescues obesity and the associated metabolic syndrome. It was reported that TFEB regulated insulin resistance and a PPARα agonist (Fenofibrate) reduces hepatic fat accumulation through the upregulation of TFEB-mediated lipophagy (28, 61, 62). In addition, Verma et al. (63) demonstrated that PPARγ expression also regulated insulin resistance in C2C12 skeletal muscle cells. MTH-3 induced TFEB and PPARγ mRNA expression when cells were treated with 5 or 10 μM MTH-3 in PA-induced insulin resistant C2C12 cells. These results suggested that MTH-3 relieves PA-induced intracellular lipid droplet deposition and increases insulin resistance in C2C12 myotubes through upregulating TFEB and PPARγ gene expression.
In conclusion, it was shown that PA induced intracellular lipid droplet deposition and increased insulin resistance in C2C12 myotubes. MTH-3 improved PA-induced-insulin resistance in C2C12 cells through increased TFEB and PPARγ expression at the mRNA level. Based on these results, MTH-3 may be considered a promising therapeutic strategy for the treatment of T2D.
Acknowledgements
The Authors would like to thank the Office of Research & Development at China Medical University, Taichung, Taiwan, R.O.C. for allowing the use of the Medical Research Core Facilities to perform the experiments and data analysis.
Footnotes
This article is freely accessible online.
Authors’ Contributions
YJC, YSL, JSY and SCT conceived and designed the study. YJC, YSL and SCT performed the experiments. YSL, JSY and SCT analyzed the data. YJC, JSY SCK and SCT wrote and modified the manuscript. All the Authors read and approved the manuscript. All Authors confirm the authenticity of all the raw data.
Funding
The present study was supported in part by the China Medical University Hospital (grant no. DMR-110-155), China Medical University (grant no. CMU103-S-16 and MOST 106-2314-B-039-046), Taipei Veterans General hospital (grant no. V110B-038), Yen Tjing Ling Medical Foundation (grant no. CI-110-6) and the Chinese Medicine Research Center, China Medical University from The Featured Areas Research Center Program within the framework of the Higher Education Sprout Project by the Ministry of Education (MOE) in Taiwan, R.O.C.
Conflicts of Interest
All the Authors have declared no conflicts of interest in relation to this study.
- Received July 7, 2021.
- Revision received July 31, 2021.
- Accepted August 5, 2021.
- Copyright © 2021 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.