


Abstract
Background/Aim: Chloride intracellular channel 4 (CLIC4) is associated with the progression of colorectal cancer (CRC). However, quantitative differences in CLIC4 expression in epithelial and stromal cells of normal mucosal tissue (NT), cancer adjacent to normal colorectal mucosal tissue (NAT), and CRC tissue remain unclear. Materials and Methods: We investigated the number of CLIC4 high-expressing (CLIC4high) cells in colorectal tissue of CRC patients and healthy individuals. Results: The number of CLIC4high cells in malignant epithelial cells at early cancerous lesions was significantly higher than that in NAT, but was significantly lower or tended to become low corresponding to the progression of colorectal carcinogenesis. Meanwhile, the number of CLIC4high cells in the stromal tissue remained low in NAT compared to late lesions. Conclusion: The number of CLIC4high cells is a useful predictor in determining the pathological condition in both malignant epithelial and stromal tissues of CRC patients.
Chloride intracellular channel 4 (CLIC4), a small globular protein (28 kDa), that belongs to the CLIC family (CLIC1, CLIC2, CLIC3, CLIC4, CLIC5A, CLIC5B, and CLIC6), and is known to function as a putative chloride channel that regulates cellular redox (1-3). CLIC4 is ubiquitously distributed in the cytoplasm, subcellular mitochondria, endoplasmic reticulum, and nuclear membrane (3, 4), and plays an important role in the regulation of development, such as endothelial morphogenesis and angiogenesis (5, 6). Recently, CLIC4 has gained attention as a significant cancer-regulating molecule. CLIC4 is broadly expressed in cancer tissues of patients with various types of cancers (7). Previously, several studies had demonstrated that CLIC4 expression is regulated by factors such as tumor suppressor p53, c-MYC, tumor necrosis factor (TNF)-α, and transforming growth factor (TGF)-β, and is expressed during cell differentiation and DNA damage (4, 8-10). CLIC4 positively or negatively affects the signals of Kirsten-ras (KRAS) and p38 mitogen-activated protein kinase (MAPK), induction of inducible nitric oxide synthase, apoptosis, cell growth, cell migration, invasion, epithelial mesenchymal transition (EMT), immune function, loss of mitochondrial membrane potential, endoplasmic reticulum stress, microRNAs, hypermethylation and tumorigenesis, as well as through differentiation of myofibroblasts and formation of the tumor microenvironment (TME) (7, 11-18). However, the function of CLIC4 depends on the type of cancer, and its variation in each cell remains elusive.
Worldwide, colorectal cancer (CRC) is a disease that exhibits a high incidence as well as mortality rate (19). Many aberrations of genes, proteins, and other molecules, such as gene mutations (e.g., KRAS and APC), gene fusions, hypermethylation, transcriptomes, microRNAs, core signal transduction, immune functions and TME formation, have contributed to initiation and promotion of CRC (20-28). CLIC4 is assumed to play an important role in these complex aberrations that contribute toward the onset of CRC. Two reports have demonstrated that the expression of human CLIC4 is negatively associated with pathological grades of CRC and with poor prognosis (7, 29). In addition, a clear association has been shown between CLIC4 expression and metastatic potential of CRC stem-like cells (29). Furthermore, CLIC4 expression in malignant stromal cells is elevated corresponding to an increase in the histopathological grade of CRC (7). The high CLIC4-expressing stromal cells are often related to myofibroblasts, which largely correspond to cancer-associated fibroblasts (CAFs) characterized by high levels of α-smooth muscle actin (α-SMA). CAFs are essential cells that assemble the TME and play a role in cancer stemness, invasion, metastasis, EMT, tumorigenesis, angiogenesis, hyperplasia of extracellular matrix, and immune response (7, 30-35). Thus, CLIC4 could be used as a biological marker representing the progression of histopathological grade of CRC and accumulation of CAFs in CRC tissue. However, differences in CLIC4 expression in colorectal epithelial cells and stromal cells among normal tissue (NT), cancer adjacent to normal colorectal tissue (NAT), and CRC tissue corresponding to the classification of cancer stages and tumor-node-metastasis (TNM) remains unclear.
In the present study, we elucidated quantitative differences in CLIC4 expression in the colorectal epithelial and stromal cells among NT, NAT, and CRC tissue at various pathological stages of cancer.
Materials and Methods
Chemicals. Anti-CLIC4-Alexa Fluor 488 mouse monoclonal antibody was obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Bovine serum albumin (BSA) was obtained from Nacalai Tesque (Kyoto, Japan). ProLong Gold Antifade reagent with 4’,6-diamidino-2-phenylindole (DAPI) was purchased from Invitrogen (Carlsbad, CA, USA). All other solvents and chemicals were of analytical grade.
Human tissues. Human CRC tissue arrays (catalog nos. CO242b, CO243a, CO243b, and CO483b) were purchased from US Biomax (Rockville, MD, USA). The arrays included 79 CRC cases, which had been characterized by age, sex, anatomic site, pathology diagnosis, TNM grading, grade, and stage. Eighteen NTs and 15 NATs were used as control samples. The company conserves the informed written consent obtained from CRC patients (or their relatives) whose tissues were used to prepare the tissue arrays. Detailed information on all cancers, NTs, and NATs can be viewed at http://www.biomax.us/tissue-arrays/Colon.
Fluorescence immunohistochemistry. Paraffin-embedded tissue arrays were de-waxed by immersion in xylene, and then washed by immersion in anhydrous ethanol, 95% ethanol in aqueous solution, and distilled water. Antigens were retrieved from the tissue sections by treating them with 1 mM EDTA buffer (pH 9.0) at 95°C for 20 min and washing with distilled water and Tris-buffer saline containing 0.1% polyoxyethylene (20) sorbitan monolaurate (TBS-T). The sections were incubated with 5% bovine serum albumin (BSA)/TBS-T (w/v) at room temperature for 1 h and later incubated at 4°C for 20 min. Subsequently, the sections were incubated with an anti-CLIC4-Alexa Fluor 488 mouse monoclonal antibody diluted 1:50 in 5% BSA/TBS-T overnight at 4°C. Next, the sections were washed with TBS-T and PBS, and finally covered with ProLong Gold Antifade reagent containing DAPI. Immunofluorescence images were acquired using a Nikon TE2000 confocal microscope equipped with EZ-C1 software (Nikon, Tokyo, Japan). The number of high-intensity CLIC4 (CLIC4high) cells in the cancer or stromal tissue per tissue area (mm2) was measured by panel photography using the confocal microscope.
Statistical analysis. All data are expressed as the mean±standard error (SE). Significant differences between two groups were assessed using Student’s t-test. Multiple comparisons were conducted using one-way analysis of variance (ANOVA) followed by post-hoc Tukey-Kramer test. Significant differences are expressed as *p<0.05; **p<0.01.
Results
Characteristics of patients with colorectal cancer. Table I shows the basic and pathological characteristics of patients with CRC and healthy individuals. Median age of CRC patients was 55.9 years (range=47.0-67.0 years). Median age of sample donors of NAT and NT was 61.4 years (range=50.5-70.5 years) and 30.2 years (a small range was observed; 29.5-30.5 years), respectively. The percentages of female participants were 29.1%, 27.0%, and 0% in CRC, NAT, and NT, respectively. Pathological stages, such as grade 2 (44.9%), stage IIIb (43.0%), T3 (46.8%), N0 (50.6%), and M0 (100%) were found in the majority of CRC patients.
Characteristics of patients with colon cancer.
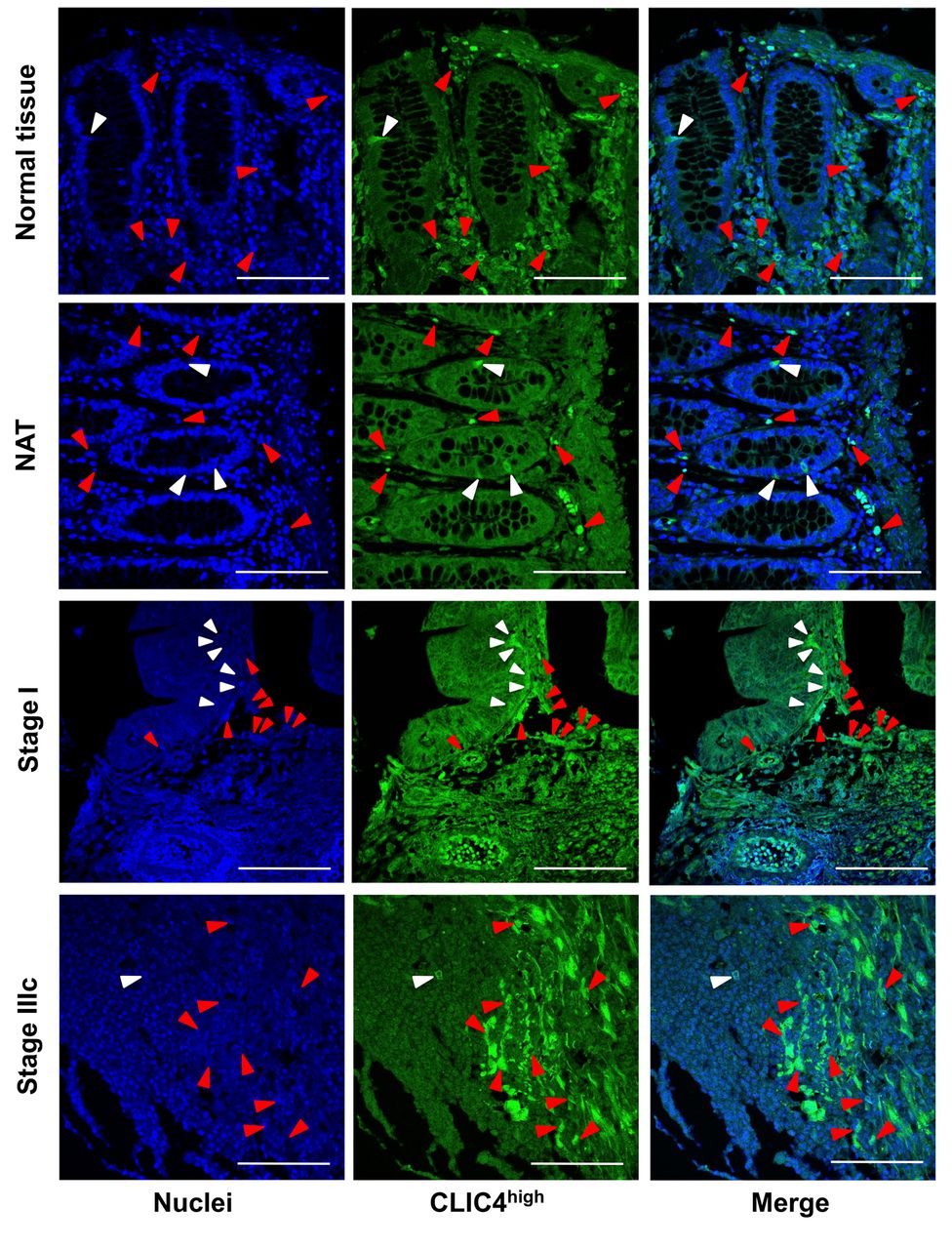
Protein expression levels of CLIC4 in malignant epithelial and stromal cells of patients with colorectal cancer. To evaluate differences in protein expression levels of CLIC4 in NT, NAT, and colorectal adenocarcinoma in patients and/or healthy individuals, the number of CLIC4high cells in normal crypt area, malignant epithelial area, and/or stromal area were determined by fluorescent immunohistochemical analysis. CLIC4high cells were detected in the all the epithelial and stromal tissues of NT, NAT, and colorectal adenocarcinoma (Figure 1). The number of CLIC4high cells in normal crypt area of NT and NAT or in malignant epithelial tissue area were significantly lower compared to those in the corresponding peripheral stroma area: normal crypt area of NT, 46.7±14.2; normal crypt area of NAT, 17.5±3.9; malignant epithelial area of adenocarcinoma, 38.6±4.0; stromal area of NT, 357.3±39.0; stromal area of NAT, 441.3±41.4; stromal area of adenocarcinoma, 308.0±18.0. No significant difference in the number of CLIC4high cells was observed among normal crypt areas of NT and NAT and malignant epithelial area, or among areas of the corresponding peripheral stroma. However, differences were observed between the stroma areas of NAT and adenocarcinoma. The numbers of CLIC4high cells in the stromal areas of NT, NAT, and adenocarcinoma were 7.6-, 25.3-, and 8.0-fold higher than that in the corresponding epithelial areas, respectively (Figure 2).

Detection of CLIC4high cells in the human colorectal tissue. The nuclei (blue fluorescence) and CLIC4 (green fluorescence) were observed by confocal microscopy. White arrow heads in normal crypt cells or malignant epithelial cells, and red arrow heads in stromal cells show the cells with high expression of CLIC4 (CLIC4high). White bars are 100 μm. NAT, Cancer adjacent normal colorectal tissue.

The different number of CLIC4high cells in normal and malignant tissue. The number of CLIC4high cells per tissue area (mm2) were measured in normal colorectal tissue (NT), cancer adjacent normal colorectal tissue (NAT) and adenocarcinoma, and shown in both epithelial area (normal crypt or malignant area) and stroma area. Mean±SE (n=15-79). **p<0.01 by Student’s t-test.
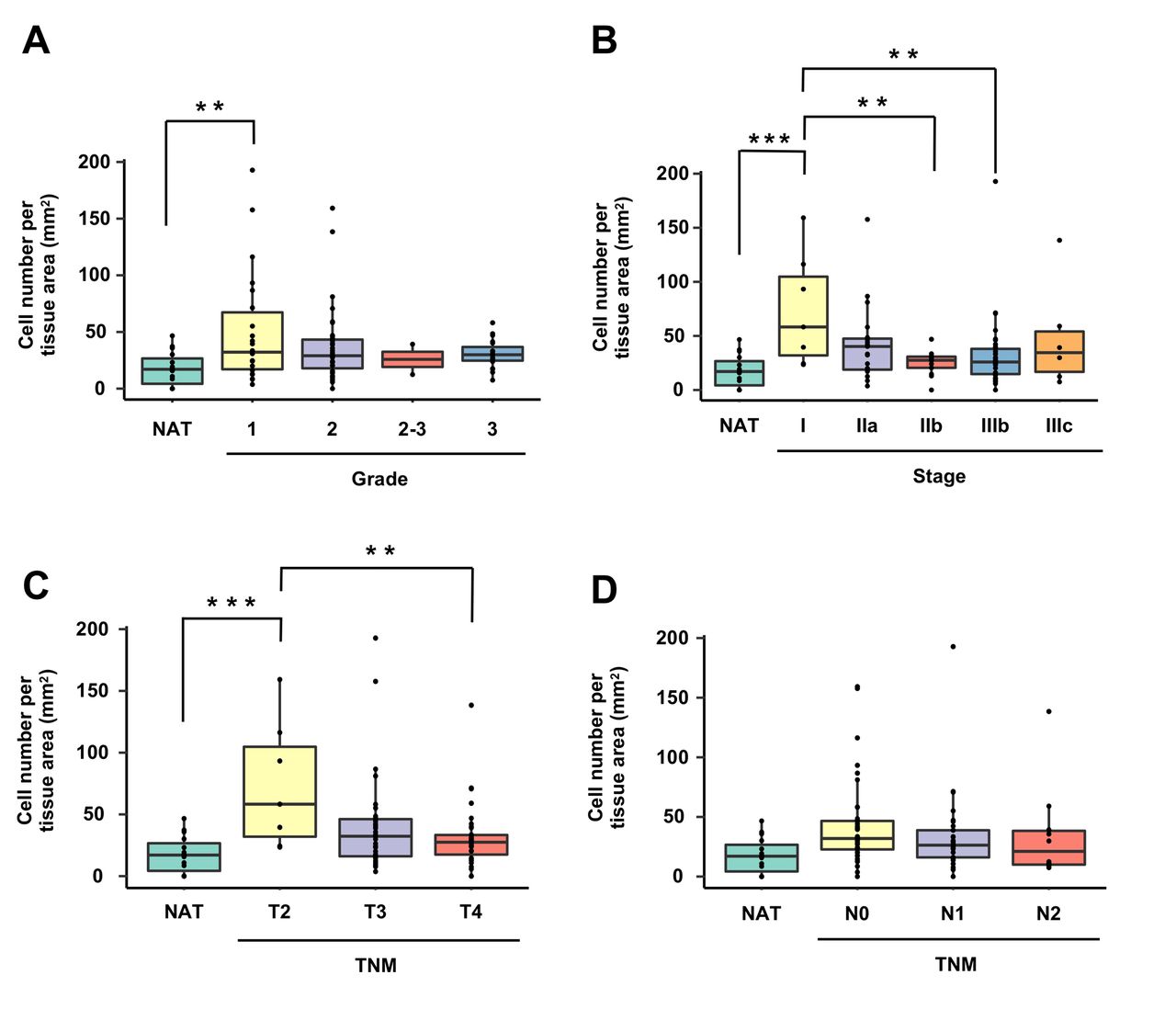
Next, protein expression of CLIC4 at each pathological classification stage was quantitatively evaluated between malignant epithelial areas (Figure 3) and stromal areas (Figure 4). The number of CLIC4high cells in the malignant epithelial area at grade 1, stage I and T2 were significantly increased by 2.9-, 4.2-, and 4.2-fold, respectively, than that in normal crypt area of NAT (Figure 3A, B, and C). However, no significant difference was observed in the number of CLIC4high cells in the malignant epithelial area among grades 1, 2, 2-3 and 4 (Figure 3A). The number of CLIC4high cells in the malignant epithelial area at stages IIb and IIIb was significantly lower than that at stage I (Figure 3B). Likewise, the number of CLIC4high cells in the malignant epithelial area at T4 was significantly lower than that at T2 (Figure 3C). No significant difference was observed in the number of CLIC4high cells in the malignant epithelial area among N0, N1 and N2 TNM classifications (Figure 3D).

The different number of CLIC4high cells in epithelial area. The number of CLIC4high cells per malignant epithelial tissue area (mm2) were estimated by each (A) cancer grade (1, 2, 2-3 and 3), (B) cancer stage (I, IIa, IIb, IIIB and IIIc), (C) tumor stage (T2, T3 and T4) and (D) node stage (N0, N1 and N2) using tumor, node, metastasis (TNM) classification. NAT, cancer adjacent normal colorectal tissue. Mean±SE (n=2-40). **p<0.01 by one-way ANOVA with a post hoc Tukey-kramer test.
The number of CLIC4high cells in the stromal area at grades 1 and 3, stage IIIb, T4, and N1 was significantly decreased compared with that in the stromal area of NAT (Figure 4A-D).

The different number of CLIC4high cells in stromal area. The number of CLIC4high cells per stromal tissue area (mm2) were estimated by each (A) cancer grade (1, 2, 2-3 and 3), (B) cancer stage (I, IIa, IIb, IIIB and IIIc), (C) tumor stage (T2, T3 and T4) and (D) node stage (N0, N1 and N2) using tumor, node, metastasis (TNM) classification. NAT, cancer adjacent normal colorectal tissue. Mean±SE (n=2-40). **p<0.01 by one-way ANOVA with a post hoc Tukey-kramer test.
Discussion
The present study demonstrated that the number of CLIC4high cells in malignant epithelial tissue of CRC patients at early cancerous lesions (grade 1, stage I, and T2) was significantly higher than that in NAT, and then significantly decreased or tended to be low along with the progression of colorectal carcinogenesis. In addition, the number of CLIC4high cells in the stromal tissue at grades 1 and 3, stage IIIb, T4, and N1 was lower than that in the stromal tissue of the corresponding NATs. This is the first report that reveals quantitative differences in CLIC4 expression in colorectal cancer epithelial tissue and stromal tissue at three pathological classifications (grade, stage, and TNM) and NAT in CRC patients and in NT of healthy individuals.
First, we quantitatively evaluated CLIC4high cells in epithelial and stromal tissues of NAT and adenocarcinoma in CRC patients and NT of healthy individuals. A negligible difference in the number of CLIC4high cells was observed among NT, NAT, and adenocarcinoma in both the epithelial and stromal tissues. However, there was a difference between the stromal tissue of NAT and adenocarcinoma; the number of CLIC4high cells in each stromal tissue was 7.6- to 25.3-fold higher than in normal crypts of NT, NAT, and malignant epithelial cells (Figure 2). Interestingly, a large number of CLIC4high cells were detected in NT and NAT. Generally, it has been reported that CLIC4 protein expression is very low in stromal cells, except for vascular endothelial cells (6, 7). However, we speculate that previous researchers might not have measured CLIC4-positive cells per tissue area in colorectal NT and NAT and therefore failed to clarify the role of CLIC4-expressing cells in colorectal stromal tissue.
Overexpression of CLIC4 in fibroblasts augments the expression of αSMA, an activated fibroblast marker in CAFs, and induces myofibroblast transformation in the stroma of tumors (7, 13). A majority of myofibroblasts covering tumor tissue is CAFs. The CAFs present in the stroma of tumors play an important role in the regulation of epithelial mesenchymal transition, angiogenesis, extracellular matrix (ECM) remodeling, tumorigenesis, drug accessibility, drug sensitivity and immune function, and production of growth and inflammation factors (26, 27, 35). Inflammatory factors such as TGF-β and TNF can induce the transformation of normal fibroblasts into CAFs (27, 35). Notably, CLIC4 is up-regulated by TGF-β and TNF-α (4, 9). Therefore, we speculate that the function of stromal CLIC4high cells in normal-looking tissue may have potential close to CAFs.
Subsequently, we quantitatively evaluated the number of CLIC4high cells in malignant epithelial and stromal tissues at three pathological classifications of cancer (i.e., grade, stage, and TNM) in CRC patients. The results demonstrated that the number of CLIC4high cells in malignant epithelial tissue of CRC patients at grade 1, stage I, and T2 was significantly higher than that in NAT, and it became significantly lower or tended to be lower according to progression of colorectal lesions. Moreover, the cell number in the stromal tissue at grades 1 and 3, stage IIIb, T4, and N1 was lower than that in the corresponding NATs (Figure 3). By contrast, a negligible difference was observed in the number of CLIC4high cells among colorectal lesions in the stromal tissue at all the pathological classifications (Figure 4). Suh et al. demonstrated that CLIC4 expression is down-regulated in tumor cells and up-regulated in tumor stroma corresponding to the progression of malignancy in CRC patients (7). Thus, our results that showed a tendency of the occurrence of lower numbers of CLIC4high cells in malignant epithelial tissue, were consistent with the previous results described above (7). However, our data on stromal tissue did not agree with those of Suh et al. This discrepancy between the two studies may be due to a difference in the methods of observation; in our research, CLIC4high cells were counted, whereas in that of Suh et al. CLIC4 positive areas were observed macroscopically. Comprehensively, our results suggested that CLIC4 expressions in malignant epithelial and stromal tissues of colorectal cancer were a useful predictor for the pathological condition in CRC, as well as that of Suh et al. (7). Another CLIC molecule, CLIC1, is also suggested to be a potential marker in patients with invasive urinary bladder cancer (36).
Nevertheless, it is not clear as to why the number of CLIC4high cells was higher in early cancerous lesions (grade 1, stage I, and T2) compared to that of NAT and why they were decreased in the later cancerous lesions, and why the cell number in the stromal tissue was lower in various degrees of pathological classifications (grades 1 and 3, stage IIIb, T4, and N1) than that of NAT. One possibility for this phenomenon may be explained by studies on microRNAs. Carofino et al. demonstrated that CLIC4 expression in tumor tissue was attenuated due to the production of microRNA-142-3p from infiltrating immune cells in a squamous cell carcinoma xenograft model mouse (37). Moreover, Huang et al. suggested that CLIC4 is regulated by microRNAs, transcriptomes, and signal pathways in human acute myeloid leukemia (17). Both malignant epithelial and stromal tissues may be consecutively exposed to CLIC4-targeting microRNAs during the progression of colorectal lesions.
In conclusion, the number of CLIC4high cells in epithelial and peripheral stromal tissues of patients with CRC was significantly suppressed corresponding to an increase in the degree of pathological classification. Further investigation is needed to clarify the molecular mechanisms underlying CLIC4 expression in malignant epithelial and stromal tissues of CRC.
Acknowledgements
This work was supported, in part, by JSPS KAKENHI Grant Number 20K05879.
Footnotes
This article is freely accessible online.
Authors’ Contributions
Reo Yokoyama and Masaru Terasaki conceived, designed the study and wrote the paper. Reo Yokoyama, Masaru Terasaki and Takuji Tanaka performed the experiments. Atsuhito Kubota, Hiroyuki Kojima, and Michihiro Mutoh reviewed and edited the manuscript. All Authors read and approved the manuscript and agree to be accounted for in all aspects of the research in ensuring that the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Conflicts of Interest
The Authors declare no conflicts of interest.
- Received August 10, 2021.
- Revision received September 10, 2021.
- Accepted September 30, 2021.
- Copyright © 2021 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}