


Abstract
Background/Aim: Asian Traditional medicines are renowned for their antitumor properties and are efficacious in the clinical treatment of various cancer types. ERM210 is a Korean traditional medicine comprising nine types of medicinal plants. In the present study, we examined the pro-apoptotic effect and molecular mechanisms of the effects of ERM210 on HepG2 liver cancer cells. Materials and Methods: The cytotoxicity of ERM210 on HepG2 cells was investigated using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide and wound-healing assays, and apoptosis and signaling pathways by fluorescence microscopy flow cytometry and western blotting. Results: ERM210 significantly impaired HepG2 cell viability and enhanced mitochondria-dependent cellular apoptosis in a time- and dose-dependent manner by up-regulating the expression of caspases 3, 7 and 9, and of BCL2 apoptosis regulator (BCL2)-associated X, apoptosis regulator (BAX) proteins, whilst down-regulating that of BCL2 protein. Furthermore, ERM210 treatment increased accumulation of cellular and mitochondrial reactive oxygen species (ROS) and significantly inhibited cell migration. Additionally, all these phenomena were reversed by treating with the ROS scavenger N-acetylcysteine. The analysis of signaling proteins revealed that ERM210 significantly up-regulated the phosphorylation of ROS-dependent mitogen-activated protein kinases (p38, extracellular-regulated kinase, and c-Jun N-terminal kinase in HepG2 liver cancer cells. Conclusion: ERM210 exerts anticancer effects in HepG2 liver cancer cells by up-regulating ROS/mitochondria-dependent apoptosis signaling, providing new insight into the possibility of employing this traditional medicine for the clinical treatment of liver cancer.
A wide variety of unique traditional herbal medicines abound in Asia. These medicines have numerous advantages over conventional chemotherapy drugs, such as low toxicity, easy absorption, and multi-target action (1, 2). According to the World Health Organization, traditional medicine is “used in the maintenance of health as well as in the prevention, diagnosis, improvement or treatment of physical and mental illness” (3). This underlies the long-established traditional use of herbal-based medicines. It includes herbal medicines and herbal finished products, which contain plants and other combination as active ingredients (4). In recent years, numerous studies have shown that herbal medicine is effective in treating insomnia (3), Alzheimer’s disease (5), cardiovascular diseases (6), polycystic ovary syndrome (7), esophageal cancer (8), and other diseases (9, 10). Herbal medicines contain abundant anticancer compounds that directly affect cancer cells and have been utilized as anticancer agents far longer than any of the chemotherapy drugs.
Cancer is an intractable human disease. Liver cancer is for the third leading cause of mortality (11). Hepatocellular carcinoma constitutes the primary and most common liver cancer subtype, with contributing risk factors including chronic viral hepatitis B and C, aflatoxin exposure, alcohol, and non-alcohol fatty liver disease or steatohepatitis. Surgical resection, radiotherapy, and chemotherapy comprise the commonly applied treatment methods for afflicted patients. However, surgical resection is more effective for patients diagnosed in the preliminary stages of the disease. For patients with highly invasive and metastatic advanced-stage hepatocellular carcinoma, chemotherapy remains the most viable option. Although remarkable milestones and considerable progress have been achieved in cancer therapeutics, first-line anticancer drugs, such as sorafenib, 5-fluorouracil, and cisplatin, produce highly toxic side-effects in patients receiving them. These include vomiting, dizziness, and hair loss. This necessitates the development of novel anticancer drugs.
Reactive oxygen species (ROS) are molecules that are not only associated with deleterious effects but also act as signaling molecules in multiple cellular signaling pathways that decide the fate of both normal cells and tumor cells. Research shows that ROS exert multifunctional regulatory actions over proteins, nucleic acids, lipids, and other biological macromolecules. When present in excess, they adversely affect the normal physiology of the cell (12-14). ROS have been implicated in tumor initiation, tumor progression, and metastasis (15). It is established that the mitochondria play multiple essential roles in intracellular ROS generation (16). However, the ROS level in tumor cells was found to be higher than those in normal cells due to their enhanced glucose metabolism, leading to mitochondrial dysfunction and oncogenic activity (17). The resulting oxidative stress may also generate potential antitumor effects. Oxidative stress can cause damage to mitochondria that is disruptive to normal function, including significantly reducing the mitochondrial membrane potential and elevating the ROS level in mitochondria, thereby inducing cell death.
Mitogen-activated protein kinase (MAPK) signaling is an important signal transduction pathway in cells, and is primarily involved in the regulation of apoptosis, cell metabolism, cell differentiation and tumorigenesis, in addition to other essential life processes (18, 19). MAPKs are an extensive family of protein kinases, including three main subfamilies: Extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and P38 MAPK. The MAPK family plays a crucial role in ROS generation-induced apoptosis (20).
An increasing number of clinical studies have reported that Asian traditional herbal medicine can relieve disease symptoms, improve the quality of life, and prolong the survival of patients with cancer. Some Asian traditional herbal medicines are considered to be uniquely advantageous in the treatment of certain tumors (21); however, their underlying mechanisms of action remain elusive. Numerous active compounds in Asian traditional herbal medicine exert a variety of antitumor effects, likely through the enhancement of the ROS level in treating patients with cancer (22, 23). Therefore, the suitable manipulation of ROS as anticancer agents is immensely challenging.
ERM210 is a formulation of extracts from nine Asian plants (Picrasma quassioides (D. Don) Benn, Polygonum cuspidatum, Sophora flavescens Solander ex Aiton, Gossypium indicum Lamarck, Picrorhiza kurroa Bentham, Chelidonium majus Linne, Cinnamomum cassia, Achyranthes bidentata, Zingiber officinale Rosc). In the present study, we demonstrated that the MAPK signaling pathway is significantly altered during ERM210-induced apoptosis of HepG2 cells.
Materials and Methods
ERM210 formulation. ERM210 is composed of nine oriental medicinal herbs (Table I). The batch used in the present study comprised the following by dry weight: 19% Picrasma quassioides (D. Don) Benn (Korea), 13% Polygonum cuspidatum (Korea), 8% S. flavescens Solander ex Aiton (Korea), 11% G. indicum Lamarck (Korea), 11% Picrorhiza kurroa Bentham (Korea), 10% Chelidonium majus Linne (Korea), 10% Cinnamomum cassia (Korea), 5% A. bidentata (Korea), and 2% Z. officinale Rosc (Korea). The herbal components were obtained from the Haeam Convalescence Hospital (Gapyeong-gun, Gyeonggi-do, Republic of Korea) and kindly authenticated by Dr. Jaihyung Lee, KMD, Ph.D. (Haeam Convalescence Hospital, Republic of Korea).
The composition of ERM210 by dry weight.
Extraction of ERM210 from the constituent herbs. The extract was prepared as follows: The dried and pulverized medicinal herbs were mixed and batches weighing 31 g each were soaked in distilled water and shaken at 37°C at a speed of 200 rpm/min (5 l). The extract was concentrated with a rotary evaporator, lyophilized and 5 g extract was obtained which was reconstituted in 1× Dulbecco’s modified Eagle’s medium (DMEM) for the in vitro studies.
Cell culture. The HepG2 hepatoblastoma cell line (ATCC®HB-8065; American Type Culture Collection Manassas, VA, USA) and QSG-7701 non-transformed hepatocyte cell lines (ATCC®CL-0419; American Type Culture Collection) were cultured in DMEM (Invitrogen, Carlsbad, CA, USA) containing 10% (v/v) fetal bovine serum (Hyclone, Logan, UT, USA), and 100 U/ml penicillin and 100 mg/ml streptomycin (Solarbio Life Sciences, Beijing, P.R. China) at 37°C in a humidified incubator under 5% CO2.
Cell viability assay. The cell viability was analyzed with the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Sigma-Aldrich, St. Louis, MO, USA) assay. Both HepG2 and QSG-7701 cells were seeded into 96-well plates at a density of 5×105 cells/ml. After culturing in DMEM for 24 h, the cells were treated for 24 h with ERM210 at different concentrations (0, 5, 10, 15, and 20%; drug concentration being 5% v/v of DMEM). Thereafter, 10 μl (0.5 mg/ml) of MTT solution was added into each well of the 96-well plates and cells were incubated for a further 2 h at 37°C under 5% CO2. Subsequently, the supernatant was removed, and formazan was solubilized in 100 μl dimethyl sulfoxide. The absorbance was measured at 490 nm using a UV MAX kinetic microplate reader (Molecular Devices, LLC, Sunnyvale, CA, USA).
Apoptosis detection. Flow cytometry was used to detect cell apoptosis with ERM210 at concentrations of 0, 10, 15, and 20%. HepG2 cells were seeded into 6-well plates at a density of 9×104 cells/ml, cultured for 12 h, and then treated with ERM210 at concentrations of 0, 10, 15, and 20% for 24 h, then cells were stained using annexin V-fluorescein isothiocyanate/propidium iodide (Solarbio Life Sciences, Beijing, PR China), according to the manufacturer’s protocol. Cells were recovered and analyzed by flow cytometry (FACSCalibur, BD Biosciences, Franklin Lakes, NJ, USA). The results were analyzed using WinMDI software (version 2. 9; BD Biosciences).
Cellular ROS detection. HepG2 cells were seeded in 6-well plates, at a density of 105 cells/ml, and then pre-treated with the ROS scavenger N-acetylcysteine (NAC; 5 μM) for 30 min or treated with ERM210 at concentrations of 0, 10, 15, and 20%. This was followed by incubation for 24 h. To evaluate the changes in HepG2 cellular ROS levels, the cells were processed using 1 μM dihydroethidium (DHE; Beyotime Biotechnology, Shanghai, PR China) and MitoSOX (Thermo Fisher Scientific, Waltham, MA, USA) staining. Cell nuclei were visualized using Hoechst 32258 (Thermo Fisher Scientific) staining (2 μg/ml) for a 20-min incubation period. Subsequently, the ROS levels in different treatment groups were qualitatively assessed under a microscope.
Wound-healing assay. Cells treated with/without ERM210, alone and pretreated with 5 μM NAC for 30 min, were seeded into 12-well plates at a density of 25×104 cells/well and then cultured in DMEM for 12 h. Subsequently, linear wounds of equal width were created using 10 μl pipette tips. The culture medium was changed to that DMEM containing 1% fetal bovine serum to be cultured for another 24 h. The cells were observed and images were captured at 0 h and 24 h using fluorescence microscopy.
Western blot assay. HepG2 cells were treated with 15% ERM210 for 0, 1, 3, 6, 12, and 24 h, following which the cells were recovered and lysed. Subsequently, the cell protein lysates were separated on 12% sodium dodecyl sulfate-polyacrylamide gels and transferred onto nitrocellulose membranes (Millipore, Bedford, MA, USA). The membranes were incubated with primary antibodies against vimentin (#sc-373717; Santa Cruz Biotechnology, Dallas, TX, USA), p-P38 (#sc-7973; Santa Cruz Biotechnology), P38 (#LF-MA0126; AbFrontier, Seoul, Republic of Korea), p-ERK (#sc-7383; Santa Cruz Biotechnology), ERK (#sc-135900; Santa Cruz Biotechnology), p-JNK (#sc-6254; Santa Cruz Biotechnology), JNK (#sc-7345; Santa Cruz Biotechnology), BCL2 apoptosis regulator (BCL2; #sc-7382; Santa Cruz Biotechnology), BCL2-associated X, apoptosis regulator (BAX; #ab90435; Abcam Cambridge, MA, USA), cleaved caspase-9 (#9505; Cell Signaling Technology, Beverly, MA, USA), cleaved caspase-7 (#sc-56063; Santa Cruz Biotechnology), cleaved caspase-3 (#9661; Cell Signaling Technology, Beverly, MA, USA), β-actin (#ab7291, dilution 1:2,000; Abcam) at 4°C for 12 h. The membranes were washed five times with tris-buffered saline containing Tween-20 [TBST;10 mM Tris HCl (pH 7.5), 150 mM NaCl, and 0.2% Tween-20] and subsequently incubated with horseradish peroxidase-conjugated goat anti-rabbit IgG or anti-mouse IgG for 1 h at room temperature. Following the removal of excess antibodies by washing with TBST, specific binding was detected using a chemiluminescence detection system (GE Healthcare Life Sciences, Little Chalfont, UK) according to the manufacturer’s protocol.
Statistical analysis. All the data from at least three independent experiments are reported as the mean±standard deviation. Repeated measures of the two-way analysis of variance were used to analyze the period and differences between groups in each experiment. Differences between groups in each experiment with values of p<0.05 were considered indicative of significant difference.
Results
ERM210 reduced cell viability by inducing apoptosis of HepG2 liver cancer cells. To investigate the cytotoxic effects of ERM210 on liver cells, HepG2 liver cancer cells and QSG-7701 normal liver cells were treated with ERM210 at different concentrations (0-20%) for 24 h. Cell viability was then detected using MTT assay as described in the Materials and Methods. The obtained results showed that ERM210 treatment significantly reduced HepG2 liver cancer cell viability at concentrations of 15% and 20% while generating weak cytotoxicity in QSG-7701 normal liver cells (Figure 1A). To clarify the effects of ERM210 on cell apoptosis, HepG2 cells were treated with ERM210 at various concentrations (0-20%) for 24 h, following which the apoptosis of HepG2 cells was detected by flow cytometry. As shown in Figure 1B and C, ERM210 treatment significantly enhanced cellular apoptosis in a dose- and time-dependent manner in HepG2 cells. To understand the effect of ERM210 on apoptosis-related protein expression, HepG2 cells were treated with ERM210 at a concentration of 15% for 0, 1, 3, 6, 12, and 24 h. The levels of apoptosis-related proteins were determined using western blot assay. The obtained results showed that after treating HepG2 cells with ERM210, the expression of pro-apoptotic proteins, such as cleaved caspase-3, -7 and -9, and BAX, was significantly up-regulated; conversely, the expression of anti-apoptotic BCL2 protein was down-regulated (Figure 1D).
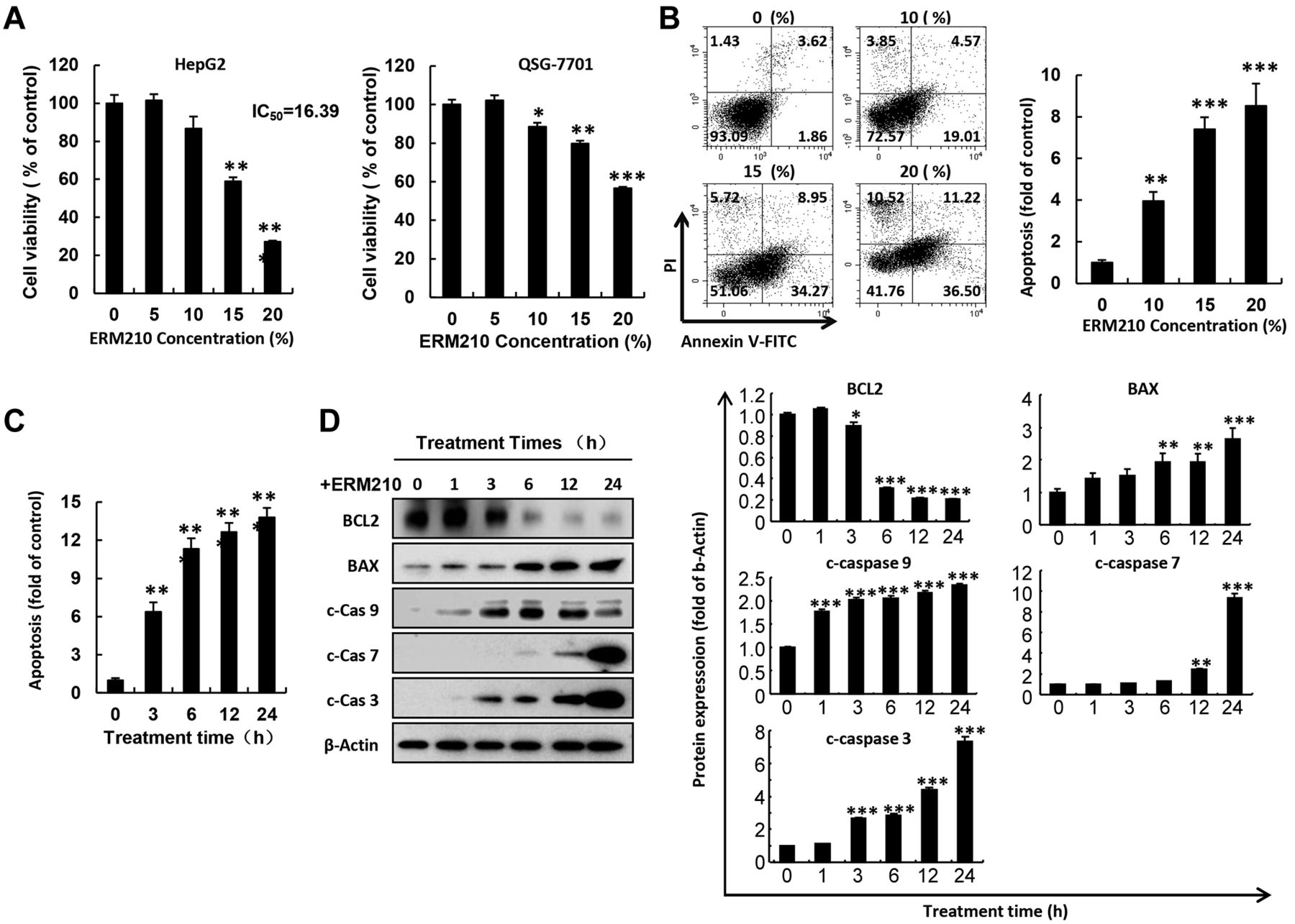
ERM210 reduces cell viability and induces concentration-dependent apoptosis of HepG2 cells. A: HepG2 liver cancer cells and QSG-7701 normal liver cells were treated with ERM210, following which their cell viability was evaluated by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. B: Concentration-dependent apoptosis was determined in HepG2 cells treated with ERM210 for 24 h, after which annexin V-fluorescein isothiocyanate (FITC) and propidium iodide (PI) staining was performed for 25 min followed by flow cytometry. A representative cytogram and quantitative analysis are shown. C: HepG2 cells were treated with 15% ERM210 for different times followed by annexin V-FITC staining. Apoptosis of HepG2 cells was quantified using flow cytometry. D: Western blotting was used to detect pro-apoptotic BCL2-associated X, apoptosis regulator (BAX), cleared caspases (c-Cas), and anti-apoptotic proteins BCL2 apoptosis regulator (BCL2) in HepG2 cells treated with ERM210 for different times. Proteins were quantitatively normalized by β-actin protein. A representative blot and quantitative analysis are shown. Significantly different at: *p<0.05, **p<0.01, and ***p<0.001.
ERM210 induced the accumulation of ROS, inhibiting HepG2 cell migration. To determine the effect of ERM210 on intracellular and mitochondrial ROS, HepG2 cells were treated with 0, 10, 15, and 20% ERM210 for 24 h. The cells were stained with DHE (whole-cell ROS detector) and MitoSOX (a detector of mitochondrial ROS) and observed using fluorescence microscopy and flow cytometry. The results showed that after treatment of HepG2 cells with different concentrations of ERM210 (0, 10, 15, and 20%) for 24 h, the intracellular and mitochondrial ROS levels significantly increased in a dose-dependent manner (Figure 2A and B). In addition, to determine the effects of ERM210 on cell-migration ability, the wound-healing assay was also performed after ERM210 treatment of HepG2 cells. As shown in Figure 2C and D, ERM210 treatment significantly inhibited cell migration of HepG2 cells by down-regulating the expression of vimentin protein (a marker of cell migration).

ERM210 induced the accumulation of reactive oxygen species (ROS) and reduced the migration of HepG2 cells. Cells were treated with different concentrations of ERM210 for 24 h. A: Intracellular ROS levels were determined by staining with dihydroethidium and Hoechst using fluorescence microscopy (left) and flow cytometry (right). B: Mitochondrial ROS levels in HepG2 cells were detected by MitoSOX staining and fluorescence microscopy (left) and flow cytometry (right). C: Wound-healing assay examined the migration ability of HepG2 cells. A representative image and quantitative analysis are shown. D: The expression of the migration marker vimentin was analyzed by western blot. A representative blot and quantitative analysis are shown. Significantly different at ***p<0.001.
ROS were found to play a crucial role in ERM210-induced cell apoptosis and migration of HepG2 cells. To verify that ROS is the principal factor modulating ERM210-induced cell apoptosis, HepG2 cells were pre-treated with NAC for 30 min and then treated with ERM210 (15%) for 24 h. The intracellular and mitochondrial ROS levels were determined by staining with DHE and MitoSOX. The obtained results indicated that NAC pre-treatment significantly reduced the intracellular (Figure 3A) and mitochondrial (Figure 3B) ROS levels in HepG2 cells. We simultaneously examined the effect of NAC on cell migration and apoptosis upon ERM210 stimulation. As shown in Figure 3C and D, NAC treatment significantly suppressed ERM210-induced cell migration and apoptosis, delineating the important regulatory role of ERM210 on ROS. Compared with untreated HepG2 cells pretreated with NAC alone as a control, there was no difference in the intracellular ROS levels of the two groups (data not shown).

Inhibition of intracellular reactive oxygen species (ROS) reduced cell apoptosis and migration of HepG2 cells. Cells were pre-treated with reactive oxygen species scavenger N-acetylcysteine (NAC) followed by treatment with 15% ERM210 for 24 h. HepG2 cells were stained with dihydroethidium, MitoSOX, and Hoechst. They were then subjected to fluorescence microscopy (upper panels) and flow cytometry (lower panels) to demonstrate the impact on the cellular (A) and mitochondrial (B) ROS levels in HepG2 cells. Representative images and quantitative analyses are shown. C: Wound-healing assays examined the ability of HepG2 cells to migrate after pre-treatment with NAC then ERM210. D: After pretreating HepG2 cells with NAC, the apoptotic effect of ERM210 on HepG2 cells was detected by flow cytometry. Significantly different at: *p<0.01, and ***p<0.001.
ERM210 induced apoptosis of HepG2 cells via MAPK signaling pathways. To clarify the molecular mechanisms underlying the effects of ROS on ERM210-induced apoptosis and cell migration, we also analyzed the expression of apoptosis- and migration-related proteins following NAC treatment. The obtained results showed that the addition of NAC reduced the expression of cleaved caspase-3, -7 and -9, and of BAX (pro-apoptotic) and vimentin (migration marker) proteins in HepG2 cells induced by ERM210 and restored BCL2 (anti-apoptotic) protein expression (Figure 4A). In order to exclude the influence of NAC on cells during the ERM210-induced initiation of the MAPK signaling pathway of HepG2 cells, we set up a separate NAC treatment group which showed that treatment with NAC alone had no effect on cells (data not shown). Furthermore, the effect of ERM210 on ROS-dependent MAPK signaling pathways was also detected by the western blot assay. As shown in Figure 4B, after treating HepG2 cells with ERM210, p38 and ERK protein phosphorylation was significantly up-regulated while that of JNK phosphorylation was down-regulated.

ERM210 induced apoptosis via mitogen-activated protein kinase (MAPK) signaling pathways in HepG2 cells. A: Western blot assays were used to detect the apoptotic effect of ERM210 on HepG2 cells. HepG2 cells were pretreated with reactive oxygen species scavenger N-acetylcysteine (NAC) for 30 min and then treated with ERM210 for 24 h. The expression of apoptosis-related proteins namely cleaved caspases (c-Cas), BCL2 apoptosis regulator (BCL2), BCL2-associated X apoptosis regulator (BAX), and the migration-related protein vimentin were analyzed by western blotting (left) and quantified (right). B: The levels of native and phosphorylated (p) p38, extracellular-regulated kinase (ERK), and c-Jun N-terminal kinase (JNK) proteins in the MAPK signaling pathway were determined after treating HepG2 cells with ERM210 for 24 h by western blotting (left) and quantified (right). Significantly different at: *p<0.05, and ***p<0.001.
Discussion
In recent years, oxidative stress has been found to play a pivotal role in the progression of several cancer types. Mitochondria are the main source of ROS, large amounts of which are generated within the electron transport chain (24). The excessive production of ROS leads to intracellular mitochondrial damage (such as a reduction of mitochondrial membrane potential, increased mitochondrial ROS and changes in the morphology of the mitochondrial ridge), which has emerged as a major focus area for current cancer treatment (25-27). Therefore, ROS play an indispensable role in cell growth, differentiation, metabolism, and proliferation.
The occurrence of liver cancer is mostly attributable to infection with hepatitis B virus, liver cirrhosis, alcoholism, smoking, and other detrimental habits (28). Studies have shown that Asian traditional herbal medicine exerts anti-inflammatory (29, 30), antibacterial (31), antitumor (32-34), and other physiological actions (35-37). Its antitumor effect is relatively significant. Asian traditional herbal medicine has demonstrated potent anticancer activity, with low toxicity and minimal side-effects; the prognosis of patients is favorable, and patient lifespan is enhanced [reviewed in (38)], Therefore, we further explored the potential of Asian traditional herbal medicines as an innovative avenue for using novel therapeutic drugs in liver cancer treatment.
In the present study, we showed that ERM210, composed of a variety of Chinese herbal medicines, induced apoptosis of HepG2 liver cancer cells. Since ERM210 contains a variety of components from Asian traditional herbal medicine, all of its main components have anticancer effects and may have toxic and side-effects on normal cells. ERM210 was significantly more toxic towards HepG2 cells than QSG-7701 cells (normal liver cells). We speculate that the reason for this may be that ERM210 has a certain targeting effect on liver cancer cells.
We optimized effective ingredients into ERM210 with the aim of producing a Chinese herbal medicine that is more effective on cancer cells and has lower toxic and side-effects on normal cells to achieve better therapeutic effects. We found that ERM210 induced a rise in the ROS level and caused mitochondrial dysfunction in HepG2 cells treated with ERM210, which in turn triggered cell apoptosis. When NAC (ROS scavenger) was added, there was a steep decline in the intracellular and mitochondrial ROS leveIs, and cell apoptosis reduced significantly. These results indicate that ERM210 induces HepG2 cell apoptosis by facilitating an increase of ROS in HepG2 cells. The MAPK signaling pathway plays an important role in cells, controlling a series of physiological activities such as cell growth, differentiation, and proliferation (39). Numerous studies have shown that MAPKs play an indispensable role in the regulation of cell physiological activity by ROS (40-42). The results of the present study showed that the MAPK signaling pathway is particularly important in the process of ERM210-induced apoptosis of HepG2 cells.
Apoptosis can be classified into the endogenous and exogenous pathways (43). When a cell is externally stimulated, mitochondrial damage within the cell releases cytochrome c and activates caspase-9. The cleavage of caspase-9 in turn cleaves caspase-3 and induces cell apoptosis (44). In the process of cell apoptosis, the BCL2 family is critical because it regulates various physiological activities of mitochondria in cells [reviewed in (45)]. The BCL2 family is divided into anti-apoptotic and pro-apoptotic proteins. When mitochondria are damaged, the levels of anti-apoptotic proteins reduces while there is a rise in the pro-apoptotic protein level. This is accompanied by a significant increase in the expression of cleaved caspase-3. Our study provides confirmation that ROS are key to ERM210-induced apoptosis of HepG2 cells as after pretreatment of the cell with NAC, intracellular and mitochondria ROS were significantly reduced in HepG2 cells. ERM210-induced cell apoptosis was also significantly reduced by the addition of NAC, with a concomitant increase in the expression of anti-apoptotic BCL2 and reduction in the expression of pro-apoptotic BAX. Additionally, the extent of mitochondrial damage, as well as the cleavage of caspases 3, 7, and 9 proteins was significantly reduced. These results all indicated that ROS may be a key factor in ERM210-induced apoptosis of HepG2 cells.
With our results, we demonstrate that the MAPK signaling pathway is considerably altered during ERM210-induced apoptosis of HepG2 cells, and plays an important role. The increase in cell survival rate upon inhibition of the intracellular and mitochondrial ROS further reiterates the important role of ROS in the process of ERM210-induced apoptosis.
ERM210 may also induce apoptosis of liver cancer cells through other methods such as changes in cell metabolism, cell membrane composition or other signaling pathways. However, since ERM210 has not yet been analyzed by liquid chromatography, the precise compounds responsible for such effects remain unknown. Nevertheless, we demonstrate that ERM210 indeed induces apoptosis of HepG2 cells through a ROS-mediated mechanism. Our results, therefore, establish the prospect of developing ERM210 as a new therapeutic drug for clinically treating liver cancer and lays a suitable foundation for the development of subsequent liver cancer treatment approaches in future studies.
Acknowledgements
The present study was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2020R1I1A2052417) and The Korean Research Institute of Bioscience and Biotechnology Research Information System (RBM0112112).
Footnotes
This article is freely accessible online.
↵* These Authors contributed equally to this study.
Authors’ Contributions
Conceptualization: JL, YXG, DPX, HNS and TK. Methodology: HNS and TK. Software: TK and HNS. Validation: TK and HNS. Formal analysis: JL, YXG, DPX, HJ, HS, JK, YHP, HNS and TK. Investigation: JL, YXG, DPX, HNS and TK. Resources: TK and HNS. Data curation: TK and HNS. Writing – original draft: JL, YXG, DPX, HNS and TK. Writing – review & editing: TK and HNS. Visualization: TK and HNS. Supervision: TK and HNS. Project administration: TK and HNS. Funding acquisition: TK. All Authors read and approved the final article.
Conflicts of Interest
The Authors declare that they have no competing interests in regard to this study.
- Received May 3, 2021.
- Revision received May 29, 2021.
- Accepted June 1, 2021.
- Copyright © 2021 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.