


Abstract
Background/Aim: Hydrogen peroxide (H2O2) is one of the reactive oxygen species (ROS), which can induce apoptotic cell death in numerous cancer cells. Pterostilbene (PTE), a natural polyphenolic compound, induces cell apoptosis in many human cancer cells. Materials and Methods: We investigated whether PTE could enhance H2O2-induced cell apoptosis in human keratinocyte HaCaT cells in vitro. The morphological change of HaCaT cells was observed and photographed under a contrast-phase microscope. The percentage of cell viability was measured by propidium iodide exclusion assay. Cell apoptosis was performed by Annexin V/PI double staining and assayed by flow cytometer. DNA condensation was measured by DAPI staining. The protein expression was determined by western blotting. ROS production-associated proteins were also assayed by confocal laser scanning microscopy. Results: PTE pre-treatment enhanced H2O2 (600 μM)-induced cell morphological changes and reduced the total cell number (cell viability). The decreased cell viability in HaCaT cells was through induction of apoptotic cell death, which was confirmed by Annexin V/PI double staining and DAPI staining. Western blotting studies indicated that HaCaT cells which were pre-treated with PTE (100 μM) and then co-treated with H2O2 (600 μM) for 12 h showed significantly increased levels of SOD (Cu/Zn), SOD (Mn), Bax, caspase-3, caspase-8, caspase-9, PARP, p53, p-p53, and p-H2A.X but decreased levels Bcl-2 and catalase. Results also showed that HaCaT cells pre-treated with PTE and then co-treated with H2O2 had increased expression of SOD (Cu/Zn) and glutathione but decreased catalase. Conclusion: These observations suggest that PTE pre-treatment can enhance the H2O2-induced apoptotic cell death in keratinocyte cells and may be an effective candidate for the treatment of proliferative keratinocytes.
Human skin covers the human body, and is exposed to various stressors (1, 2), including environmental factors such as UV-light (3, 4). Actually, skin plays a vital role in the body against exposure to environmental factors (temperature, humidity, and sun exposure) (5). Skin aging is the most common human aging aspect and one of its main reasons is oxidation injury of the skin (6). Reactive oxygen species (ROS) caused by UV light plays an essential role in skin aging (7). Furthermore, UVB radiation-induced DNA lesions may lead to DNA mutation during cell division and then induce the initiation of carcinogenesis (7). Keratinocyte hyperproliferation is highly associated with skin pathogenesis, such as skin cancer, psoriasis, keratosis, and verrucae (8, 9). Therefore, dynamic regulation of apoptosis and cell proliferation in keratinocytes is important for the function of the skin.
Cellular ROS, primarily arising from cell oxidative metabolism, plays an important role in cell signaling and homeostasis and in both chronological aging and photoaging of skin (7). Ozone (O3), hydroxy radical (OH•), singlet oxygen (1O2), hydrogen peroxide (H2O2), and superoxide anion radical (O2•−) belong to ROS (10). Hydrogen peroxide (H2O2), one type of ROS, is produced under normal circumstances and is increased due to external stressors. In cells, the formation of H2O2 is linked to other forms of ROS and has been associated with cellular senescence (11). H2O2 has been reported to induce ROS in cells, which caused oxidative stress (12) and can directly or indirectly damage cells for inducing apoptosis and necrosis (13, 14). The majority of the epidermis consists of keratinocytes (15), which are very susceptible to oxidative stress (16, 17). Earlier reports have shown the importance of antioxidants on attenuating oxidative stress, which is associated with chronic diseases (18, 19). Thus, H2O2 was recognized to be used to induce oxidative stress in vitro models.
Natural compounds can be effective candidates for various skin diseases. Pterostilbene (PTE; trans-3,5-dimethoxy-40-hydroxystilbene), a natural polyphenolic compound, is also a natural dimethylated analog of resveratrol found in Pterocarpus marsupium Roxb. (Fabaceae) (20) and is also present in fruits such as blueberries, grapes, and tree wood (21). Numerous studies have shown that PTE has anticancer effects on a wide range of human cancers in vitro, including breast (22), cervical (23), leukemic cells (24), colorectal cancer (25), lung squamous cell carcinoma cells (26), hepatocellular carcinoma cells (27), multiple myeloma (28), and oral cancer (29). PTE has been the focus of several studies due to its potent antioxidant and anti-inflammatory properties (30), particularly in neuronal cells. PTE exerts neuroprotection against oxidative toxicity via oestrogen receptor α signaling pathways in human neuronal cells (31) and in hypoxia-induced brain injuries (32). It also inhibits ROS production and apoptosis by the rapamycin signaling pathway in primary spinal cord neurons (33). Besides, PTE also attenuates mitochondrial oxidative damage induced by cerebral ischemia-reperfusion injury (34).
Apoptosis, a type of programmed cell death, which constitutes multiple molecular mechanisms to maintain tissue homeostasis (35), is classified into the extrinsic and intrinsic apoptosis pathways (36). The extrinsic apoptosis pathway is triggered by apoptotic ligands binding to death receptors and TRAIL receptors (37). The intrinsic apoptosis pathway which is the response to cellular stress (38), leads to a decrease in mitochondrial membrane potential and cytochrome c release from mitochondria. Then activated caspase-9 and caspase-3 induce cell apoptosis (39). Thus, the induction of cancer or abnormal cell apoptosis is one of the strategies for cancer therapy.
PTE has been demonstrated to present cell apoptotic biological activity. No reports have shown results of PTE co-treated with H2O2 in skin cells; thus, herein, PTE was evaluated as to whether or not it can enhance H2O2-induced cell apoptosis in human keratinocyte HaCaT cells in vitro. We investigated the molecular mechanisms in pre-treatment with PTE and H2O2 treatment in HaCaT cells.
Materials and Methods
Chemicals and reagents. Pterostilbene (PTE), dimethyl sulfoxide (DMSO), propidium iodide (PI), and trypsin-EDTA were obtained from Sigma Chemical Co. (St. Louis, MO, USA). For HaCaT cell culture, Dulbecco’s Modified Eagle Medium (DMEM) medium, fetal bovine serum (FBS), L-glutamine, and antibiotic (penicillin/streptomycin) were purchased from GIBCO®/Invitrogen Life Technologies (Carlsbad, CA, USA). Primary antibodies included anti-SOD (Cu/Zn), -SOD (Mn), -catalase, -β-actin, -p53, and -p-p53 were obtained from Santa Cruz Biotechnology, and anti-Bax, -Bcl-2, -caspase-3, -caspase-8, and -caspase-9 from Cell Signaling Technology, Inc. (Beverly, MA, USA). Anti-MGMT, -PARP, -p-ATM, and -p-ATR antibodies were purchased from Calbiochem (San Diego, CA, USA), and anti-p-H2A.X from GeneTex Inc. (Irvine, CA, USA). The HRP-conjugated secondary antibody was purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA). DMSO was used to dissolve PTE for further diluted in the culture medium.
Cell culture. Human keratinocyte HaCaT cells were kindly provided by Professor Huey-Chun Huang (China Medical University, Taiwan). HaCaT cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM) medium supplemented with 1% antibiotic (100 units/ml penicillin and 100 μg/ml streptomycin), 10% fetal bovine serum (FBS), and 2 mM L-glutamine at 37°C in a 5% CO2 humidified incubator as described previously (40).
Cell viability and morphology observation. HaCaT cells (1×105 cells/well) were cultured in 12-well plates with DMEM medium overnight and cells were treated with H2O2 (600 μM), PTE (100 μM), or pre-treated with PTE at 100 μM for 1 h and then co-treated with H2O2 (600 μM) for 3, 6, and 12 h. After treatment, cells were observed and photographed under contrast phase microscopy and then cells were mixed with PI solution (4 μg/ml) and analyzed by flow cytometry as described previously (41).
Annexin V/PI staining for apoptotic cell death. Apoptotic cell death was performed by using Annexin V/PI double staining assay according to the manufacturer’s instructions, as described previously (41). In brief, HaCaT cells (1×105 cells/well) in 12-well plates were treated with H2O2 (600 μM), or pre-treated with PTE at 100 μM for 1 h and then co-treated with H2O2 (600 μM) for 12 h. At the end of incubation, cells were harvested and resuspended in Annexin V binding buffer and incubated with Annexin V/PI in the dark for 15 min. After incubation, cells were analyzed using BD FACSCalibur (BD Biosciences) for apoptotic cell numbers. Experiments were performed in triplicate.
DAPI assay. HaCaT cells (1×105 cells/well) were seeded on the 12-well plates and treated with H2O2 (600 μM), or pre-treated with PTE at 100 μM for 1 h and then co-treated with H2O2 (600 μM) for 12 h, and cells were examined and photographed under fluorescent microscopy at ×200. Cells were collected and fixed in 4% paraformaldehyde in PBS for 20 min at room temperature. Cells were stained with DAPI solution (2 μg/ml) and examined and photographed using a fluorescence microscope as described previously (42).
Western blotting analysis. HaCaT cells (2×106 cells) were cultured onto 10-cm dishes overnight and treated with H2O2 (600 μM), or pre-treated with PTE at 100 μM for 1 h and then co-treated with H2O2 (600 μM) for 12 h. Cells were harvested and were lysed using a lysis buffer [120 mM NaCl, 40 mM Tris (pH 8), and 0.1% NP-40] on ice. Each protein concentration was detected and quantified by the Protein Assay Reagent Kit (Bio-Rad, Hercules, CA, USA). Aliquots of the protein solutions (30 μg) from each treatment were electrophoresed on a 10% SDS-PAGE and transferred onto polyvinylidene difluoride (PVDF) membranes (Millipore, Belford, MA, USA). The blot was then incubated overnight at 4°C with a primary antibody [anti-SOD (Cu/Zn), - SOD (Mn), -catalase, -β-actin, -caspase-3, -caspase-8, -caspase-9, -Bax, -Bcl-2, -MGMT, -PARP, -p-ATM, -p-ATR, -p-H2A.X, -p53, and -p-p53] diluted in PBST buffer (phosphate-buffered saline with 0.1% of Tween-20) and 1% BSA. The membranes were washed with PBST, incubated with peroxidase-conjugated secondary antibodies diluted in PBST buffer for 1 h at room temperature. The immune-labeled bands (protein bands) were then detected chemiluminescence signals using ECL detection kit (Amersham Biosciences ECLTM) as described previously (42, 43).
Observation of confocal laser scanning microscopy. HaCaT cells (5×104 cells/well) were maintained on coverslips and treated with H2O2 (600 μM), or pre-treated with PTE at 100 μM for 1 h and then co-treated with H2O2 (600 μM) for 12 h. At the end of treatment, cells were fixed with 4% formaldehyde in PBS and were permeabilized by using 0.2% Triton-X 100 in PBS for 15 min. Subsequently, cells were washed with PBS and probed with anti-SOD (Cu/Zn), -catalase, and -glutathione primary antibody. After washed with PBS, cells were stained by FITC-conjugated goat anti-mouse IgG (green fluorescence), and the nucleus was stained by PI (red fluorescence). All samples were examined and photographed under a Leica TCS SP2 Confocal Spectral Microscope, as described previously (44, 45).
Statistical analysis. All data are presented as mean±S.D. Statistical analysis were performed by one-way analysis of variance followed by Dunnett’s or Tukey test for comparing with the control group (***p<0.001) or determining significant differences among group means (p<0.05), respectively, with the Sigma Plot 12 software (Systat Software, Inc., San Jose, CA, USA).
Results
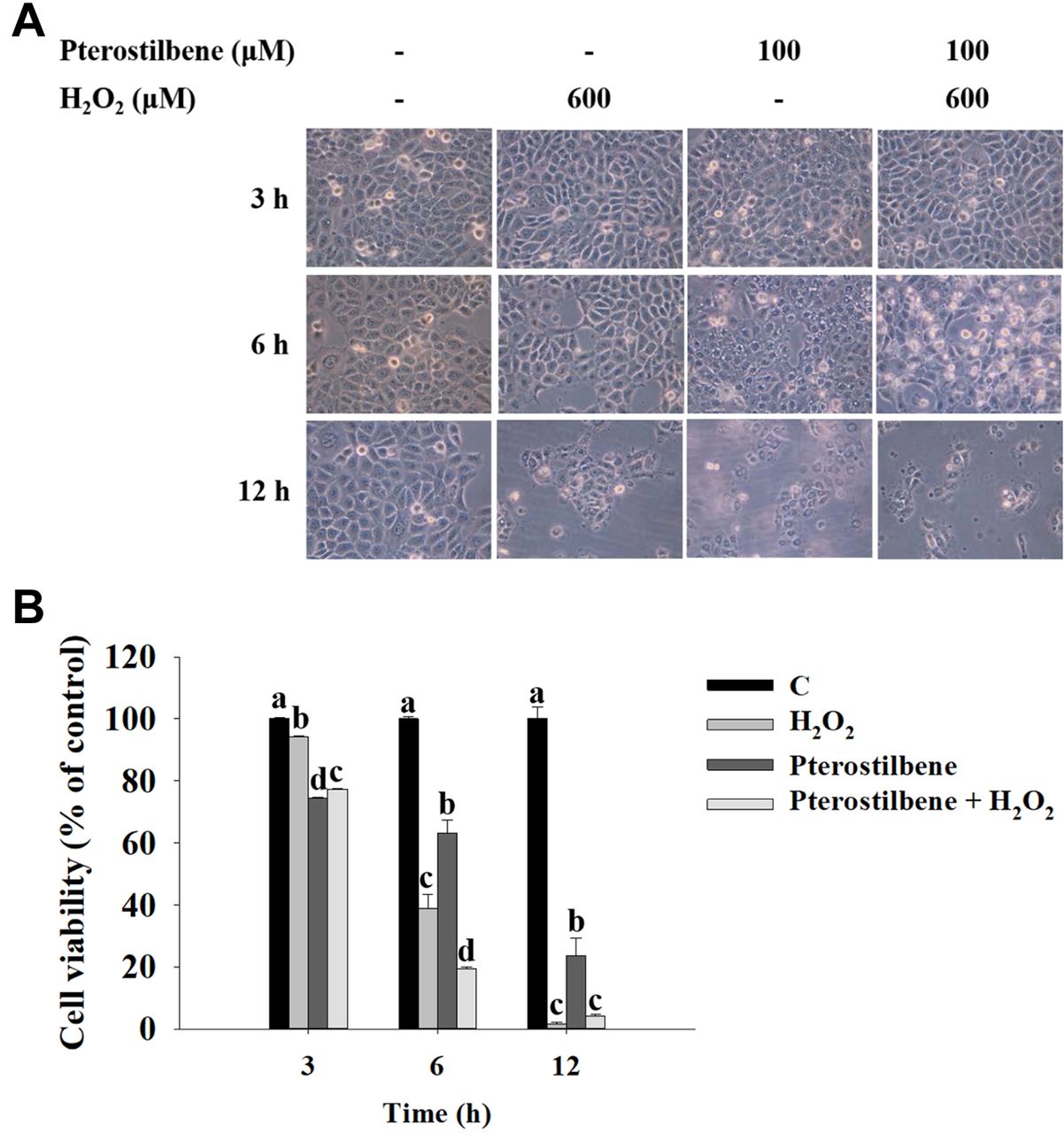
PTE affects morphology and total cell viability in HaCaT cells. HaCaT cells were treated with H2O2 (600 μM), PTE (100 μM), or pre-treated with PTE at 100 μM for 1 h and then co-treated with H2O2 (600 μM) for 3, 6, and 12 h, and cell morphology was monitored and photographed under phase-contrast microscopy (Figure 1A). Subsequently, the total cell viability was calculated by PI exclusion assay and results were presented in Figure 1B. As shown in Figure 1A and B, PTE significantly increased H2O2-induced cell morphological change and cell death at 6 and 12 h treatment based on cell morphological change and reduced total cell number (cell viability).

Pterostilbene affected cell morphology and total cell viability in HaCaT cells. HaCaT cells were treated with H2O2 (600 μM), pterostilbene (100 μM), or pre-treated with pterostilbene at 100 μM for 1 h and then cells were co-treated with H2O2 (600 μM) for 3, 6, and 12 h. Cells were monitored the cell morphology and photographed under phase-contrast microscopy (A), and total cell viability was calculated by flow cytometric assay (B) as described in Materials and Methods. Group comparisons showing different letters indicated significant differences by one-way analysis of variance and Tukey post hoc test are shown (p<0.05).
PTE induces apoptotic cell death in HaCaT cells. HaCaT cells were placed on 12-well plates and treated with H2O2 (600 μM), or pre-treated with PTE at 100 μM for 1 h and then co-treated with H2O2 (600 μM) for 12 h. After treatment, cells were doubly stained with Annexin V and PI and examined under microscopy, and total apoptotic cell death was measured by flow cytometric assay. As shown in Figure 2A and B, results indicated that H2O2 (600 μM) treatment alone induced 72% of early- and late-stage apoptotic cells; however, after pre-treatment with 100 μM of PTE led to 88%. These results indicated that HaCaT cells pre-treated with PTE induced higher number of H2O2 (600 μM)-induced apoptotic cells than that of H2O2 (600 μM)-treated alone.

Pterostilbene induced apoptotic cell death in HaCaT cells. HaCaT cells were placed on the 12-well plates and treated with H2O2 (600 μM), or pre-treated with pterostilbene at 100 μM for 1 h and then cells were co-treated with H2O2 (600 μM) for 12 h. Cells were analyzed by Annexin V/PI kit as described in Materials and Methods. (A) Representative profiles of apoptotic cell death. (B) Percentage of apoptotic cell death. ***p<0.001, significant difference between experimental groups and the control as analyzed by Dunnett’s test.
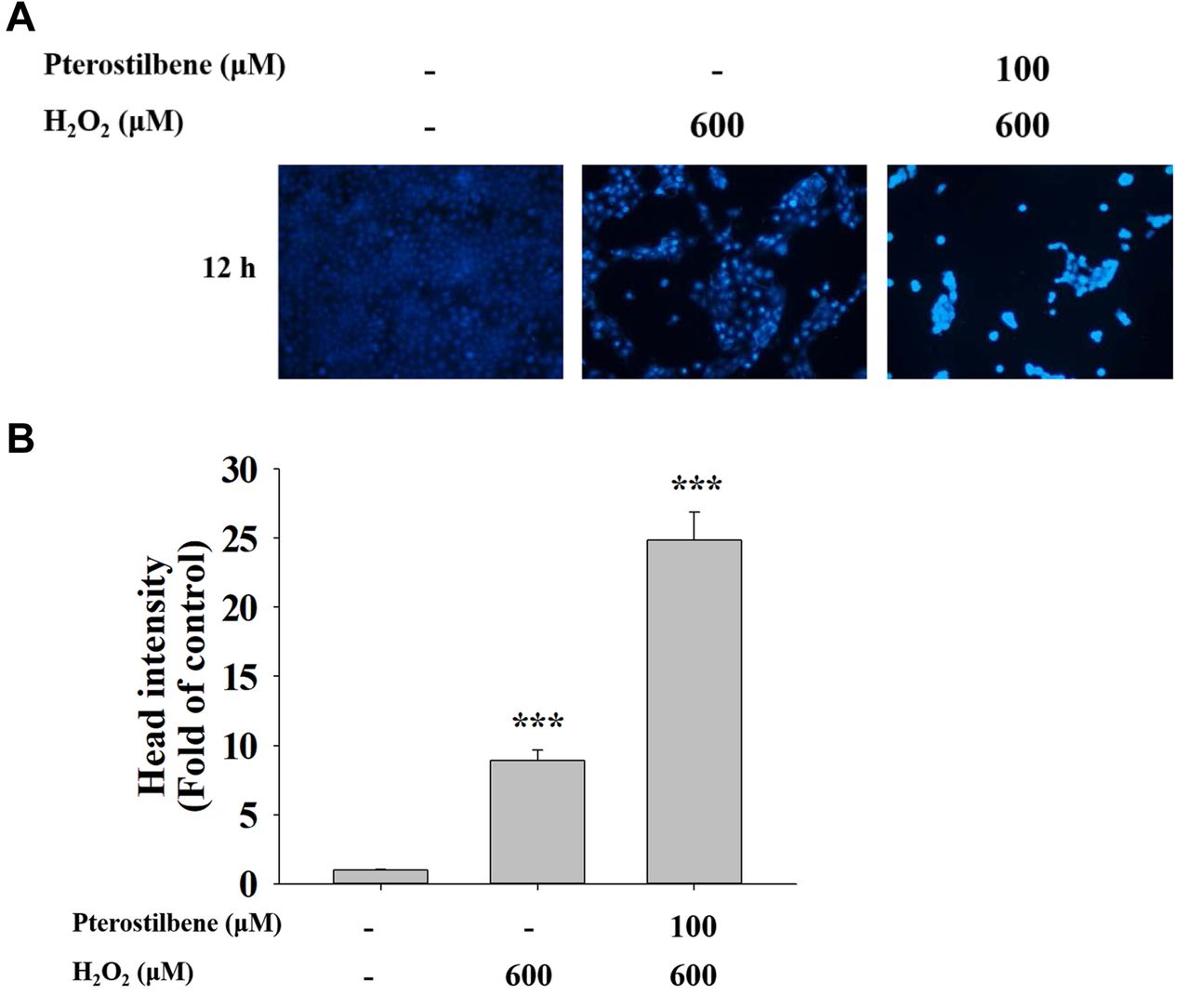
PTE enhances chromatin condensation (apoptotic cell death) in HaCaT cells. HaCaT cells were placed on 12-well plates and treated with H2O2 (600 μM), or pre-treated with PTE at 100 μM for 1 h and then co-treated with H2O2 (600 μM) for 12 h. Cells were stained with DAPI solution and results are shown in Figure 3. Results indicated that HaCaT cells pre-treated with 100 μM of PTE and treated with H2O2 (600 μM) led to an increase in DNA condensation (apoptotic cells) compared to cells with H2O2 (600 μM) treatment alone. These results indicated that cells pre-treated with PTE enhanced the effects of H2O2 treatment on the increase in the number of apoptotic cell death.

Pterostilbene pre-treatment induced DNA condensation (apoptotic cell death) in HaCaT cells. HaCaT cells (1×105 cells/well) were seeded in the 12-well plates and treated with H2O2 (600 μM), or pre-treated with pterostilbene at 100 μM for 1 h and then co-treated with H2O2 (600 μM) for 12 h and cells were examined and photographed under fluorescent microscopy at ×200, as described in Materials and Methods. (A) Representative photographs of cell DNA condensation. (B) Fluorescent intensity of DNA staining. ***p<0.001, significant difference between experimental groups and the control as analyzed by Dunnett’s test.
PTE affects ROS production, DNA damage, and apoptosis-associated proteins in HaCaT cells. HaCaT cells were treated with H2O2 (600 μM), or pre-treated with PTE at 100 μM for 1 h and then co-treated with H2O2 (600 μM) and protein expression was examined by western blotting. As presented in Figure 4A, B and C, pre-treatment with PTE and then treatment with H2O2 increased the expressions of SOD (Cu/Zn) and SOD (Mn) but decreased that of catalase in HaCaT cells (Figure 4A). Besides, pre-treatment with PTE (100 μM) and co-treated with H2O2 (600 μM) for 12 h significantly increased the levels of DNA repair associated proteins, such as p-ATM, p-ATR, p53, p-p53, p-H2A.X, and MGMT in HaCaT cells (Figure 4B). Furthermore, pre-treatment with PTE (100 μM) and treatment with H2O2 (600 μM) for 12 h significantly increased the expressions of Bax, cleaved caspase-9, cleaved caspase-8, cleaved caspase-3 and PARP but decreased that of Bcl-2 in HaCaT cells (Figure 4C).
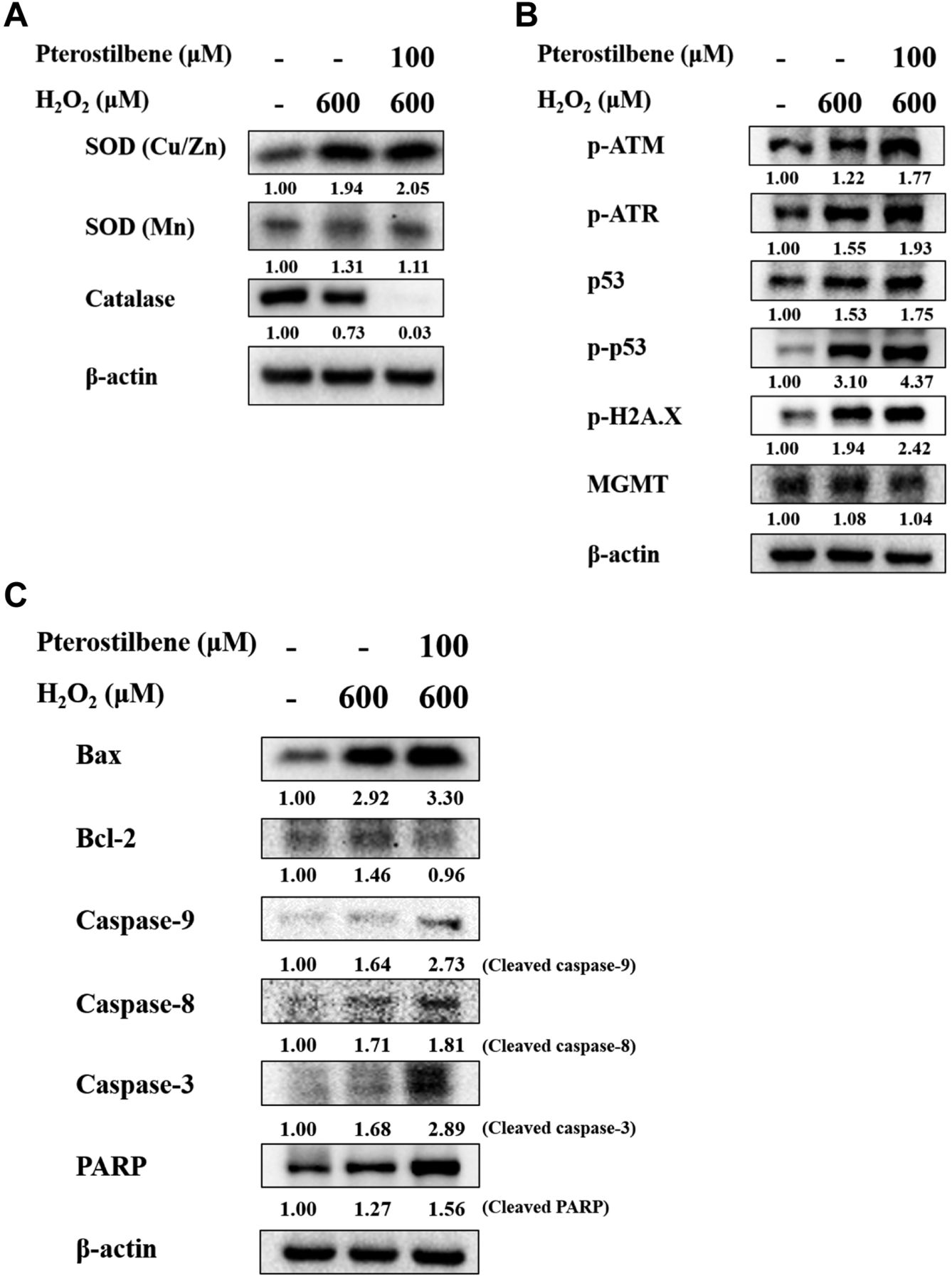
Pterostilbene enhanced ROS, DNA damage, and cell apoptosis associated proteins in HaCaT cells. HaCaT cells were treated with H2O2 (600 μM), or pre-treated with pterostilbene at 100 μM for 1 h and then co-treated with H2O2 (600 μM) for 12 h, and cells from each treatment were harvested for protein expression estimation by western blotting, as described in Materials and Methods. (A) SOD (Cu/Zn), SOD (Mn), and catalase; (B) p-ATM, p-ATR, p53, p-p53, p-H2A.X, and MGMT; (C) Bax, Bcl-2, caspase-9, caspase-8, caspase-3, and PARP.
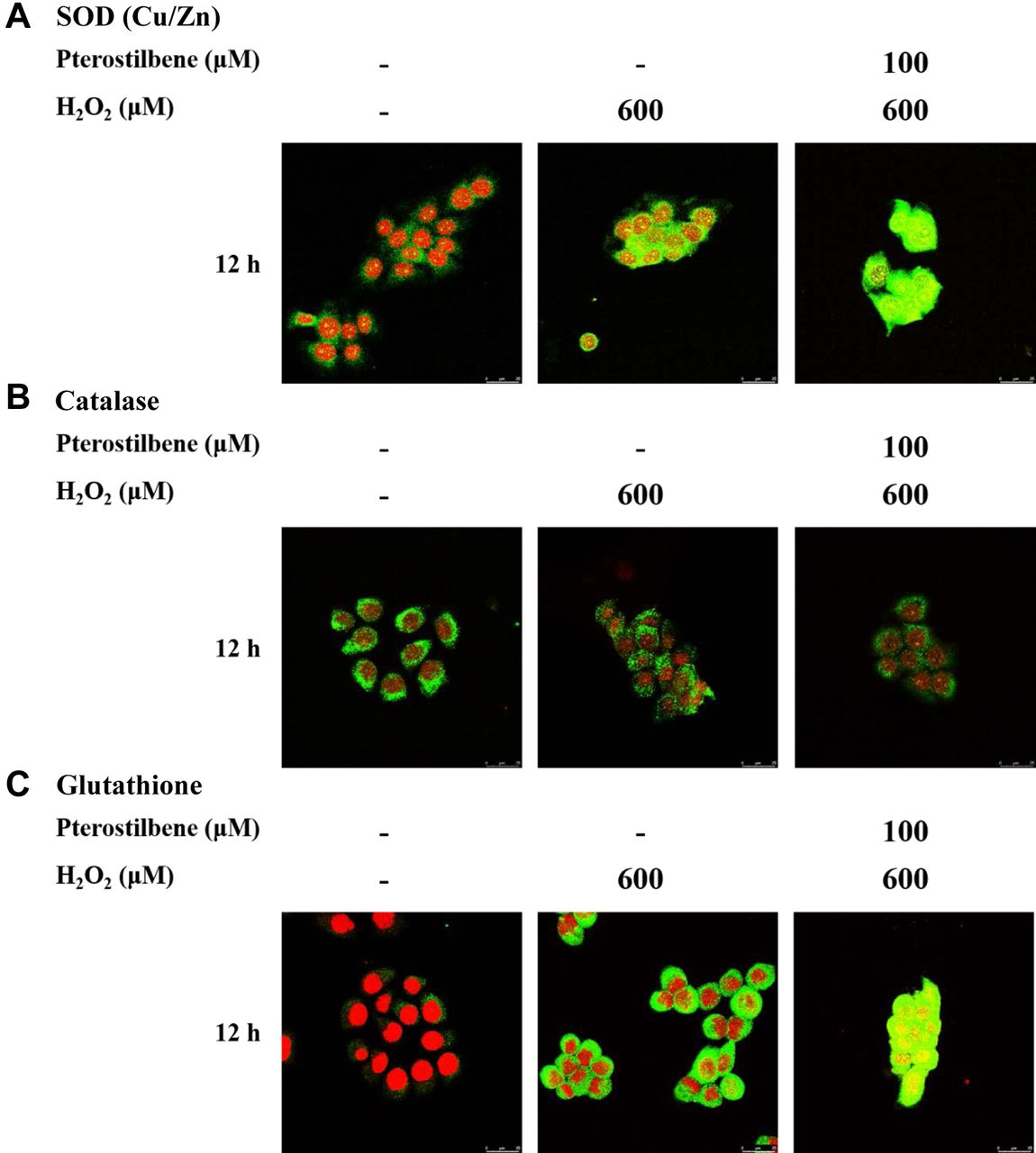
PTE affects the expression of SOD (Cu/Zn), catalase, and glutathione. We further investigated the expression of oxidation-association protein that is involved in the oxidative stress in HaCaT cells. Cells were exposed to H2O2 (600 μM), PTE (100 μM), or pre-treated with PTE at 100 μM for 1 h and then co-treated with H2O2 (600 μM) for 12 h and probed with SOD (Cu/Zn), catalase, and glutathione antibodies, and observed under confocal laser microscopy system. As shown in Figure 5, PTE promoted the expressions of SOD (Cu/Zn) and glutathione but decreased that of catalase in HaCaT cells.
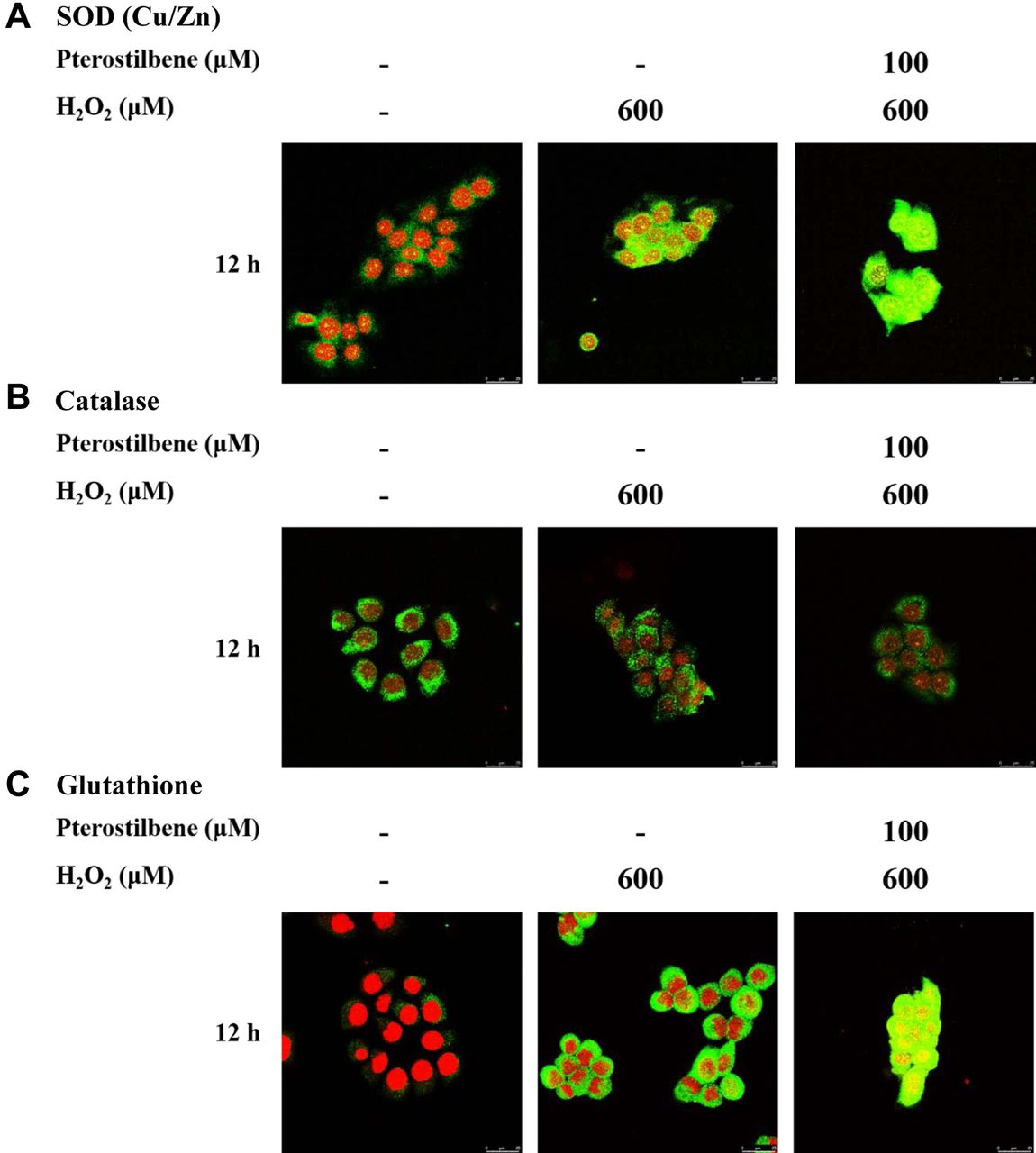
Pterostilbene affected ROS-associated proteins in HaCaT cells. HaCaT cells (5×104 cells/well) were maintained on coverslips and treated with H2O2 (600 μM), or pre-treated with pterostilbene at 100 μM for 1 h and then co-treated with H2O2 (600 μM) for 12 h. Cells were fixed with 4% formaldehyde, permeabilized by using 0.2% Triton-X 100 for 15 min, and were probed with anti-SOD (Cu/Zn) (A), -catalase (B), and -glutathione (C) primary antibody and then by FITC-conjugated goat anti-mouse IgG (green fluorescence). Finally, the nucleus was stained by PI (red fluorescence) as described in Materials and Methods.
Discussion
It is well documented that ROS plays a prominent role in cell metabolism, which regulates cell proliferation, differentiation, and death (38); however, excessive production of ROS and overall cell antioxidant capacity may induce irreversibly severe oxidative damage to the cells (46, 47). Antioxidants reduced skin injury, aging, and cancer risk and some of them have been extracted from natural products to present antioxidant activities (48, 49). The study’s major reason is that there is no available information showing the influence of PTE at a high dose which enhanced H2O2-induced cell apoptosis through oxidative stress in human skin cells. Therefore, we investigated the effects of PTE pre-treatment at a high dose on the increase of cell apoptosis induced by H2O2 in human epidermal keratinocytes (HaCaT) cells in vitro. HaCaT cells are non-tumor epidermal keratinocytes and these cells bear the mutant p53; thus, numerous studies already use it (50-52).
H2O2 is a type of ROS and can also be used as an inducer of oxidative stress in in vitro and in vivo studies. Herein, we found that H2O2 reduced viable cell numbers in HaCaT cells; however, cells that were pre-treated with PTE (100 μM) and then co-treated with H2O2 (600 μM) had significantly decreased total cell viability (Figure 1B) when compared to H2O2 treatment alone. Cells loss of their original morphology may be caused by exposure to the chemical agents. Herein, we observed cell morphological changes accompanied by reduced cell numbers after exposure to PTE and H2O2. It was reported that intracellular redox disrupted ROS balance, if excessive ROS accumulation may lead to oxidative stress and induced oxidative damage (53, 54). ROS, causing most free radicals, can damage cell proteins, lipids, and DNA (55). In cells, ROS keeps at low levels to avoid extracellular invaders and to maintain cellular signaling. Furthermore, ROS is involved in cell migration, proliferation (56), and differentiation (57) in keratinocytes. The doses of H2O2 may lead cells to survival or death. Here, the dose of H2O2 used, was of high amount, that caused cell death.
Cell death can be divided into cell necrosis and apoptosis; the former leads to damaged cell membranes and death; the latter is involved in programmed cell death called cell apoptosis. Therefore, to further investigate whether combined treatment (PTE and H2O2) led to enhancing cell apoptotic death, Annexin V/PI reagents were used to stain cells, then cells were assayed by flow cytometry, and results indicated that HaCaT cells pre-treated with PTE (100 μM) and co-treated with H2O2 (600 μM) for 12 h had increased cell morphological changes (Figure 1A) and decreased total viable cell number (Figure1B). Furthermore, this increased apoptotic cell death compared to H2O2-alone-treated cells (Figure 2A and B) by Annexin V/PI double staining assay, a well-documented method for cell apoptosis assay. For further confirming these observations, we used the DAPI staining assay, a well-known protocol for examining cell apoptosis. Results indicated that HaCaT cells pre-treated with PTE and co-treated with H2O2 had increased apoptotic cell death based on increased DAPI intensity (Figure 3).
It is well known that H2O2, a type of ROS, is one of the factors involved in oxidative stress. However, epidermal keratinocytes are very susceptible to oxidative stress in the skin (16, 17). To understand the effects of PTE on the expression of endogenous antioxidant enzymes, HaCaT cells were pre-treated with and without PTE (100 μM) and then treated with H2O2 for 12 h and then cells were harvested for western blotting. Results from Figure 4C indicate that PTE pre-treatment increased SOD (Cu/Zn) and SOD (Mn), two superoxide dismutases, but decreased catalase when compared to H2O2 treatment alone in HaCaT cells. These results were also confirmed by confocal laser microscopy examination (Figure 5A, B, and C). The enzymatic antioxidants such as superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPX), glutathione reductase, and thioredoxin reductase (TRX) are the components of the skin defense system against ROS-mediated damages and they played crucial roles for the protection of skin (58). Based on this observation, pre-treatment of PTE (100 μM) and then co-treated with H2O2 for 12 h did not exert protection or antioxidant activity in HaCaT cells because of the increased number of apoptotic cell death.
Diminished apoptosis and excessive proliferation of keratinocytes in skin diseases have been reported in psoriatic lesions. Psoriasis is a common chronic inflammatory skin disease characterized by hyperproliferation with incomplete differentiation of epidermal keratinocytes and decreased keratinocyte apoptosis (59). Induction of apoptosis may be a potent therapy for psoriasis. Results from Figure 4C indicated that PTE pretreatment enhanced H2O2-induced apoptosis and resulted in elevated Bax, PARP, active caspase-9, caspase-8, and caspase-3 but decreased anti-apoptotic protein Bcl-2 in HaCaT cells. It was reported that H2O2 induced apoptosis via the production of ROS (60). Cancer cells also have been shown to evade apoptosis to maintain their survival and metastasis (61, 62). It was reported that several molecular mechanisms are involved in the apoptosis suppression of tumor cells. Tumor cells avoid apoptosis through the expression of anti-apoptotic proteins (Bcl-2) or through down-regulation or mutation of pro-apoptotic proteins (Bax). The expressions of Bcl-2 and Bax are regulated by the p53 tumor suppressor gene (63). Bcl-2 and Bax are Bcl-2 members who are localized in the mitochondria and the ratio of Bax and Bcl-2 are involved in cell apoptosis. Herein, results from Figure 4C indicated PTE pretreatment enhanced the expressions of caspase-8 and caspase-3. Thus, both expressions may lead to cell apoptosis via the extrinsic signaling pathway. Furthermore, caspase-9 also was activated, indicating the involvement of the intrinsic signaling pathway in HaCaT cells in vitro.
In conclusion, in the present study, we explored the mechanism of PTE on enhancing the apoptotic effects of H2O2 on HaCaT cells. Our results indicated that PTE enhanced the effects of H2O2 on apoptosis induction via extrinsic and/or intrinsic signaling pathways in HaCaT cells, as shown in Figure 6.

The possible molecular mechanism of apoptosis induction by pterostilbene pre-treatment and then H2O2 treatment in HaCaT cells.
Acknowledgements
This work was supported by Grant CMU-108-MF-109 from China Medical University, Taichung, Taiwan. Experiments and data analysis were performed in part through the use of the Medical Research Core Facilities Center, Office of Research & Development at China medical University, Taichung, Taiwan.
Footnotes
Authors’ Contributions
Study conception and design: YCC, YCC and SFP; Acquisition of data: YCC, PYC, TDW, CLC and SFP; Analysis and interpretation of data: YCC, YPH, TCH and SFP; Drafting of manuscript: YCC, YCC and SFP; Critical revision: YCC and SFP. All Authors discussed the results and commented on the article.
This article is freely accessible online.
Conflicts of Interest
The Authors confirm that there are no conflicts of interest.
- Received October 15, 2020.
- Revision received December 6, 2020.
- Accepted December 16, 2020.
- Copyright© 2021, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.