


Abstract
Telomeres compose the end portions of human chromosomes, and their main function is to protect the genome. In hematological disorders, telomeres are shortened, predisposing to genetic instability that may cause DNA damage and chromosomal rearrangements, inducing a poor clinical outcome. Studies from 2010 to 2019 were compiled and experimental studies using samples of patients diagnosed with hematological malignancies that reported the size of the telomeres were described. Abnormal telomere shortening is described in cancer, but in hematological neoplasms, telomeres are still shortened even after telomerase reactivation. In this study, we compared the sizes of telomeres in leukemias, myelodysplastic syndrome and lymphomas, identifying that the smallest telomeres are present in patients at relapse. In conclusion, the experimental and clinical data analyzed in this review demonstrate that excessive telomere shortening is present in major hematological malignancies and its analysis and measurement is a crucial step in determining patient prognosis, predicting disease risk and assisting in the decision for targeted therapeutic strategies.
The word telomere originates from the Greek words τέλοφ (telos, end, extremity)+μέροφ (meros, part), meaning “part of the extremity”. The telomeres compose the end portions of chromosomes and consist of many 5’-TTAGGG-3’ non-coding repeats (1).
During a normal lifespan, telomeres naturally shorten. However, this can become a problem when excessive telomere shortening is observed in stem cells of both pediatric and adult patients. Healthy cells are predisposed to a division limit, also known as Hayflick limit, of 50 to 70 divisions before the cell undergoes senesce or apoptosis due to telomere attrition (2, 3).
Short telomeres are associated with genetic instability, which may characterize them as a predisposition factor for hematological malignancies (4, 5).
Human Telomeres and their Functions
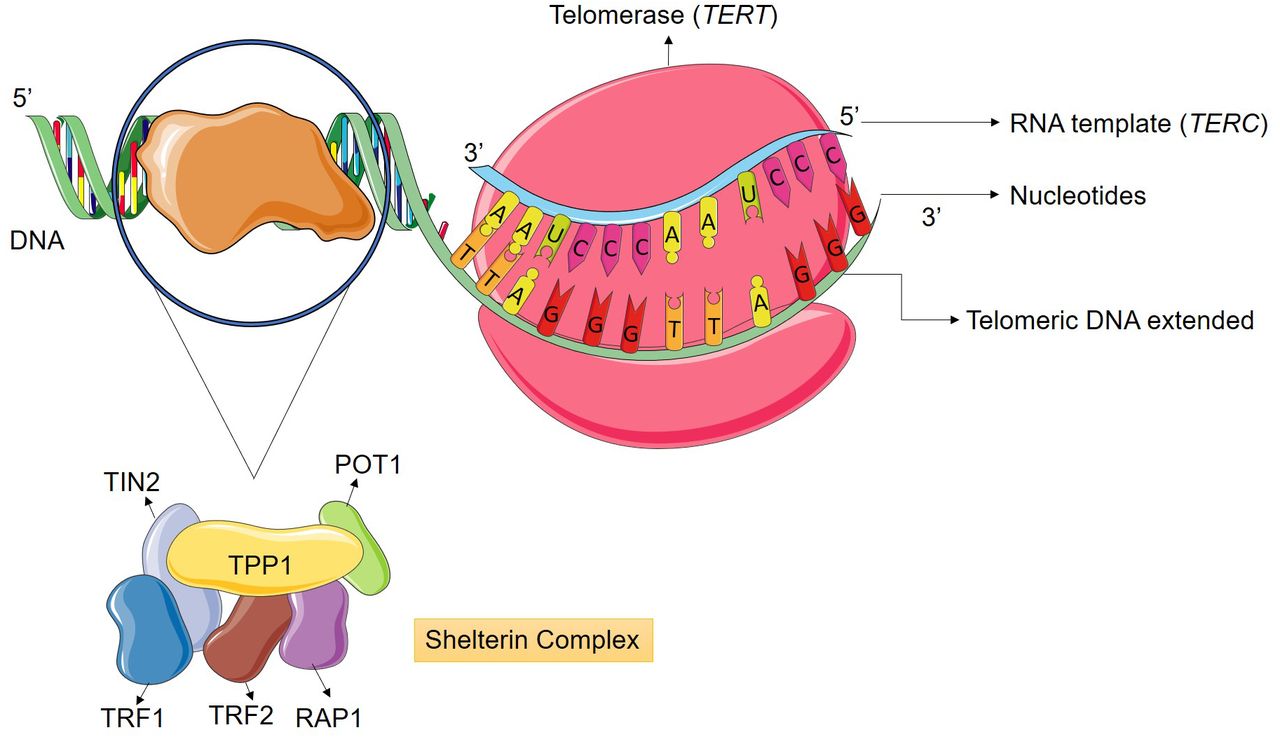
Human telomeres are the end portions of chromosomes, which act primarily in DNA protection and genomic stability, preventing fusions and damage to the genetic material. Telomeres are composed of approximately 1000 to 2000 base pairs or 5-12 kb of non-coding TTAGGG repeats, which interacts with a group of proteins denominated as Shelterin complex (Figure 1) (3, 6-8).
A major characteristic of telomeres is their progressive shortening that happens naturally with each division of somatic cells. The shortening occurs due to the DNA end replication problem, which consists of the inability of the cellular machinery to effectively synthesize the chromosomes ends during replication, alongside with the lack or insufficiency of pathways that promote telomere elongation, such as telomerase activity or the alternative lengthening of telomeres (ALT) (9-11).
Telomere elongation occurs primarily by human telomerase (hTERT) activity, which is a ribonucleoprotein enzyme specialized in neutralizing telomeric DNA attrition by synthesizing new TTAGGG repeats at chromosome ends. This enzyme is composed by two subunits; the catalytic domain is the telomerase reverse transcriptase (TERT), which utilizes the telomerase messenger RNA (TERC) subunit as a complementary template for the addition of nucleic acids to the telomere. The replicative immortality in most tumors is associated with a loss of homeostasis in the telomere-telomerase complex, being characterized by telomere elongation and in 90% of the cases it is due to the reactivation of telomerase enzyme (Figure 2) (12-14).
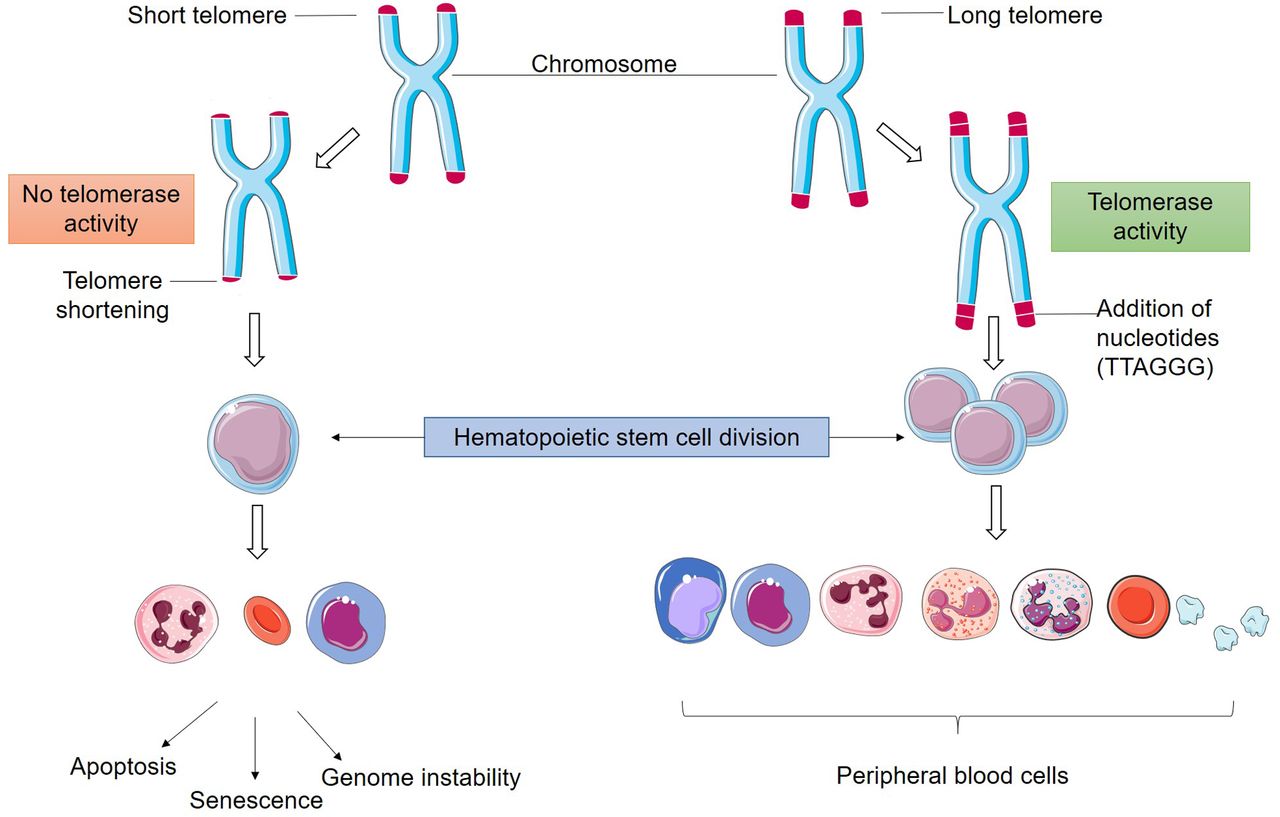
Hematopoietic stem cells express intrinsic telomerase activity which is required to sustain their accelerated replication cycle. When telomerase activity is lacking hematopoietic stem cells become unstable, leading to DNA damage and consequent bone marrow failure (5).
Telomere and Telomerase in Human Cancer
Cells with malignant potential can adapt and escape replicative senescence, by suppressing DNA damage response mechanisms or by over-activating telomere maintenance pathways, such as telomerase activity, and thus acquiring immortality and initiating the tumorigenesis process (15).
Dysfunctions affecting the telomere complex might expose the DNA to enzymatic degradation leading to breaks, deletions, fusions and accumulation of DNA mutations. The consequence of this genomic instability is cancer progression and a worse prognosis for the afflicted patients (16, 17).
The risk for cancer development aggravates in relation to the decrease in telomere length (TL) and after the disease is consolidated the telomeres tend to elongate once again. This observation shows that critically shortened and dysfunctional telomeres contribute to genetic instability and oncogenesis (1, 17, 18).
Telomeres in Hematopoietic Dysfunction
The lifespan of mature hematopoietic cells is very limited, demanding their constant production and replacement by the bone marrow. It is expected that hematopoietic cells replicate 50 to 200 times before proceeding to replicative senescence (19).
The telomere attrition in stem cell is accelerated during an individual's early years of life, when one stem cell will replicate from 15 to 30 times. During the adult life, between 50 and 60 years, telomere shortening progresses more slowly, however, after this period it accelerates once again. In T lymphocytes 33 base pairs of telomeric sequence are usually lost every year, while B lymphocytes only lose around 15 base pairs a year (19, 20).
Mature hematopoietic cells derive from the proliferation of hematopoietic stem and progenitor cells (HSPC). The stem cells are capable of auto-renewal and production of other stem cells, however, this renewal will only occur when these cells are near senescence and in need of replacement. Usually, the telomeres of stem cells shorten over the years, but when this shortening is excessive it may lead to bone marrow insufficiency and failure, predisposing stem cells to malignancy (19, 21, 22).

Telomerase enzyme counteracts nucleotide loss during cell replication. The catalytic domain has telomerase reverse transcriptase (TERT) activity, which utilizes the telomerase mRNA (TERC) subunit as a complementary template for the addition of nucleotides to telomeric DNA. The Shelterin complex is composed of six proteins (POT1, TPP1, TRF1, TRF2, TIN2 and Rap1) responsible for maintaining genetic integrity, assisting in the prevention of excessive DNA damage and regulating telomere-telomerase interaction. The complex protects telomeric DNA from unwanted degradation and chromosomal fusions (13).
Although the standard model of oncogenesis for the majority of cancers predicts the presence of telomerase over-expression and consequent telomere elongation, some cancer subtypes are able to maintain an indefinite proliferation by expressing telomerase but not necessarily elongating the telomeres, as is the case of hematological neoplasms (1, 10, 23, 24).
Hematological Malignancies
Most hematological neoplasms possess an uncertain etiology, however, there are some predisposing factors. Malignant transformations initiate due to complex interactions between genetic factors such as chromosomal rearrangements, aneuploidies, punctual mutations, deletions, insertions, duplications and amplifications. Among them, chromosomal translocations and aneuploidies are the main biomarkers for genetic instability and development of hematological malignancies (25, 26).
The dysfunctional shortening of telomeres may be present in congenital anemias, as in Fanconi anemia or Shwachman Diamond syndrome, but disease progression and telomere shortening do not have a strong association. However, telomere shortening alongside chromosomal mutations and telomerase reactivation may be predisposing factors for the development of hematological neoplasms such as myelodysplastic syndrome and leukemias (5, 27).
Hematological disorders usually present similar clinical signs amongst each other, making an early differential diagnosis necessary. The occurrence of telomere shortening in hematological malignancies is associated with major genomic rearrangements and DNA damage that is not detected in cells with longer telomeres. This observation leads to the understanding that the study and analyses of TL is of utmost importance for a better prognosis of patients in the clinical practice (15, 28).
Telomere Length and Hematological malignancies
Table I presents studies relating telomere length and prognosis of patients afflicted by hematological malignancies.
Myelodysplastic syndromes (MDS) are a heterogeneous group of clonal diseases that are characterized by an ineffective hematopoiesis consequentially leading to cytopenia and dysplasia in the peripheral blood. MDS patients express a progressive shortening of the telomeres which leads to persistent physiological damage to the DNA and other classical MDS characteristics, demonstrating a correlation between TL, MDS pathophysiology and progression to malignancy as acute myeloid leukemia (15, 21, 48, 49).
Cytogenetics and prognosis associated with shorter telomeres at diagnosis of hematological malignancies.
Leukemias are malignant disorders characterized by increased numbers of white blood cells in the peripheral blood or bone marrow. In chronic leukemias there is usually a prevalence of defective mature cells that may have a myeloid origin, as in chronic myeloid leukemia (CML), or a lymphoid origin, as in chronic lymphoid leukemia (CLL). On the other hand, in acute leukemias there is a predominance of blastic cells, being determined as acute myeloid leukemia (AML) or acute lymphoid leukemia (ALL) (50).
Amongst acute leukemias, ALL patients express shorter telomeres than AML patients, however, as the diseases progress, AML telomeres tend to shorten in an increased pace compared to ALL telomeres. ALL expressing a malignant phenotype for B lymphocytes present shorter telomeres than those from AML (41, 51, 52).
CML usually originates from a reciprocal translocation between chromosome 9 and chromosome 22, creating the Philadelphia chromosome (Ph+) and the chimeric gene BCR-ABL. Telomeres are shorter in CML during the accelerated phase and blast crisis when compared to the chronic phase. Moreover, in patients afflicted by CLL, there is an association between shorter telomeres and an unfavorable prognosis (15, 41, 52-54).
Telomere Length Studies
The most common neoplasms related to shorter telomeres and age are MDS and AML (49). Patients with short telomeres present a greater risk for developing MDS, even though only a minor percentage actually does (36, 49).
The TL of patients afflicted by AML and those with a high risk for MDS were analyzed simultaneously; in both cases TL was similar (31, 35). In a study by Warny et al., TL was analyzed in three stages, the first was at disease diagnosis, the second was during treatment, where telomere elongation was already observed, and the third at patient relapse when it was estimated that TL was shorter than at the time of diagnosis (31).
Patients diagnosed with AML have shown a notable shortening of telomeres and those with longer telomeres present an increased overall survival at 6-month period compared to those with shorter telomeres (37, 38). Patients with complex cytogenetics tend to express shorter telomeres in comparison to those with a normal karyotype. The TL also changes in comparison to treatment outset and complete remission, showing an elongation pattern in the bone marrow cells from patients at treatment completion, when compared to normal cells (55).
Acute Promyelocytic Leukemia (APL) is an AML subtype presenting an incidence of 10 to 15% of AML cases (56). Telomeres in APL patients are shorter in comparison to healthy controls and their length is comparable to that of patients afflicted by regular AML. Patients in complete remission from APL demonstrate considerable telomere elongation, suggesting that TL is a predictive marker for patient prognosis (39).
In regard to patients diagnosed with ALL presenting complex karyotypes, the telomeres are shorter than those of patients harboring normal karyotypes (55). It has also been observed that adult patients diagnosed with ALL have short telomeres, indicating aggressive disease (51, 55).
In CLL, telomeres are shorter in B lymphocytes lineages than in T lymphocytes and these shorter telomeres are also associated with chromosomal aberrations (44, 45). In a study conducted by Steinbrecher et al., shorter telomeres were observed in CLL patients compared to controls, and 83% of the analyzed patients had telomeres shorter than the average (33).
Defining the TL in CLL might help predict the disease risk and propose a specific therapeutic course for patients, as shorter telomeres have a strong association with an inferior progression free survival (PFS) and disease aggressiveness (29, 30, 34, 40, 42, 46, 47, 57, 58).
Mantle cell lymphoma (MCL) is an aggressive lymphoma subtype, representing only 4% of lymphoma cases (59). In a study conducted by Jebaraji et al., TL in MCL cells was not significantly associated with chromosomal aberrations, while CLL patients harboring complex karyotypes had a strong association with shorter telomeres, suggesting that the increase in aberration incidence was a consequence of genomic instability caused by telomere attrition (43).
Hairy cell leukemia (HCL) is a mature B lymphocyte leukemia, which presents a distinct immunophenotype and is classified in two subgroups: the classic HCL and HCL variant (HCL-v) (60). In a study conducted by Arons et al., it was shown that patients afflicted by HCL had longer telomeres than those afflicted by HCL-v, suggesting that the poorer prognosis associated with HCL-v might be due to increased telomere attrition (41). Furthermore, in a study by Walsh et al., TL was found to be longer in classic HCL patients than in those diagnosed with CLL (61).
Ohyashiki et al., observed that individuals diagnosed with CML had shorter telomeres in comparison to healthy controls, increased predisposition to developing blast crises as well as greater disease severity during the chronic phase. Patients undergoing blastic phase presented shorter TL than expected by their age and also when compared to chronic phase patients (62). Caocci et al., suggested that telomere shortening in CML cases happens before Philadelphia chromosome origination, and influences chromosomal instability and disease progression (63).
Bouillon et al., found that telomeres of CML patients were shorter in leukemic stem cells than in hematopoietic stem cells. Furthermore, leukemic cells can alter normal hematopoietic cells through extrinsic pathways that impact in the differentiation and auto-renewal cellular capacity, also altering the TL of hematopoietic cells. This study suggested that non-leukemic cells may already present a deficit in TL that would contribute to disease onset and progression (32).
Telomere homeostasis disorders have been heavily associated with leukemogenesis and neoplasm treatment, especially when these disorders induce telomere attrition. The “telomere crises” influences resistance to chemotherapy as their shortening might induce karyotype complexity and malignant cell adaptation, allowing their increased proliferation to overcome treatment cytotoxicity (42, 43, 47, 64, 65).
Conclusion
Experimental and clinical data analyzed in this review demonstrate that excessive telomere shortening is present in major hematological malignancies and its analyses and measurement is a crucial step in determining patient prognosis, predicting disease risk and assisting in the decision for targeted therapeutic strategies.
Acknowledgements
This study was supported by Brazilian funding agencies National Counsel of Technological and Scientific Development (CNPq; to RCM, MEAM and CAMN).
Footnotes
Authors' Contributions
Nogueira BMD, Machado CB and Moreira-Nunes CA designed the study; Nogueira BMD, Machado CB and Moreira-Nunes CA prepared the figures; Nogueira BMD, Machado CB, Montenegro RC, Moraes MEA and Moreira-Nunes CA wrote the article; Nogueira BMD, Machado CB and Moreira-Nunes CA revised the final version. All Authors read and approved the final article.
This article is freely accessible online.
Conflicts of Interest
The Authors declare no conflicts of interest regarding this study.
- Received July 27, 2020.
- Revision received August 6, 2020.
- Accepted August 10, 2020.
- Copyright© 2020, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.