


Abstract
Background: In order to investigate the combination effect of anticancer drugs and X-ray irradiation on neurotoxic side-effects (neurotoxicity), a method that provides homogeneously X-ray-irradiated cells was newly established. Materials and Methods: PC12 cell suspension was irradiated by X-ray (0.5 Gy) in serum-supplemented medium, immediately inoculated into 96-microwell plates and incubated overnight. The medium was replaced with fresh serum-depleted medium containing 50 ng/ml nerve growth factor to induce differentiation toward nerve-like cells with characteristic neurites according to the overlay method without changing the medium. The differentiated cells were treated by anticancer drugs as well as antioxidants, oxaliplatin or bortezomib, and the viable cell number was determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide method. Results: Antioxidants and anticancer drugs were cytotoxic to differentiating PC12 cells. Combination of anticancer drugs and X-ray irradiation slightly reduced cell viability. Conclusion: The present ‘population irradiation method’ may be useful for the investigation of the combination effect of X-ray irradiation and any pharmaceutical drug.
X-Rays are used for many purposes in various fields. This includes the visualization of the distribution and therapeutic effects of drugs by X-ray computed tomography (1); the assessment of chemical element changes by energy dispersive X-ray spectrometry (2); the analysis of crystal structure by X-ray diffraction (3); the analysis of oral mucosal distribution of trace metal elements by X-ray fluorescence with synchrotron radiation and particle-induced X-ray emission, and the estimation of chemical states of elements by X-ray absorption fine-structure analysis (4); the visual inspection of passenger baggage in airports by X-ray images (5); and therapeutic applications in medicine (6). Radiation, such as X-rays, and drugs, are reported to exert dose-dependent biphasic effects (7).
X-Ray irradiation has been reported to stimulate the production of reactive oxygen-species such as superoxide and nitrite (8, 9), the secretion of Fas ligand (10), the accumulation of cells in the G2+M phase in the cell cycle (11), and the shortening of telomeres (12). On the other hand, at lower doses of X-irradiation, beneficial effects (known as hormesis) can be induced. For example, X-irradiation at 10 mGy (but not 50 or 100 mGy) reduced the frequency of micronucleated cells in human lymphocytes to below the spontaneous level (13).
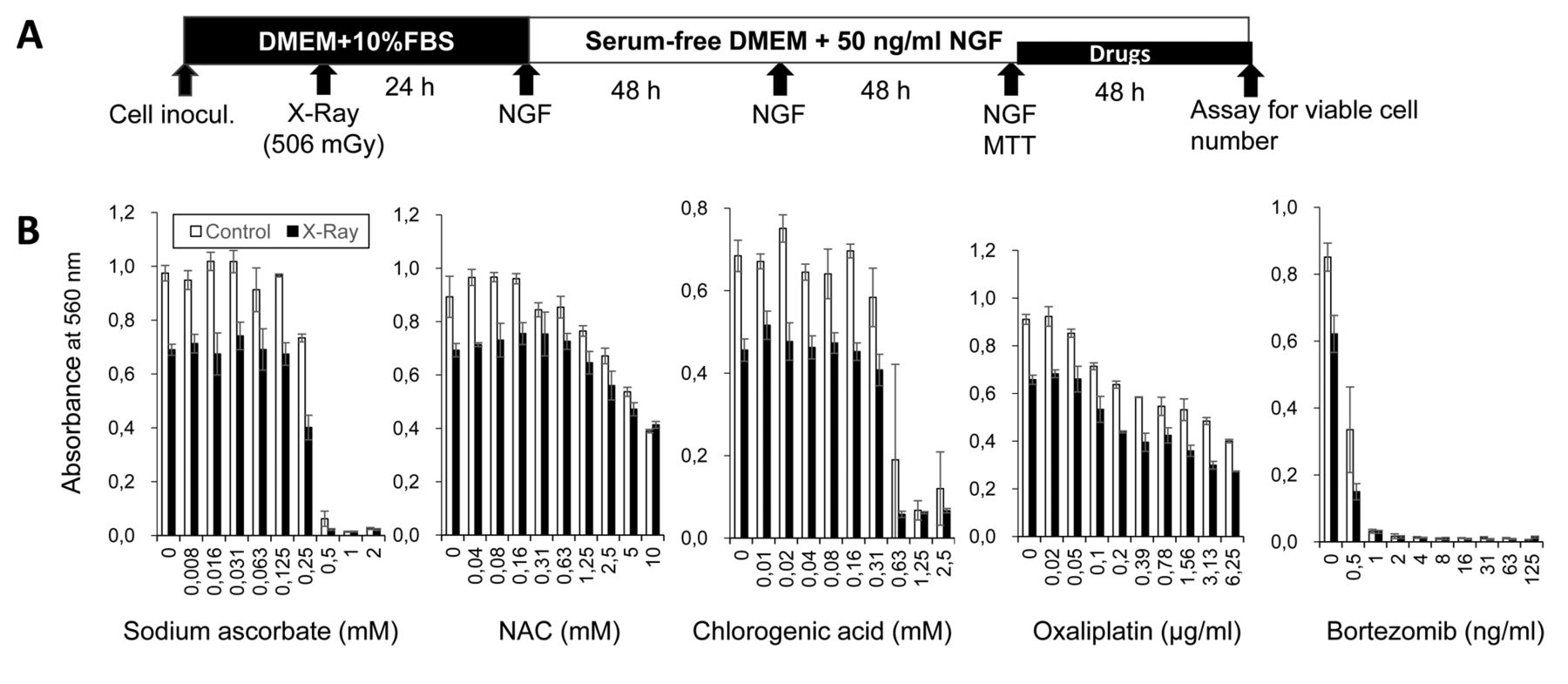
Recently, we discovered that among anticancer drugs, platinum preparations (cisplatin, carboplatin, oxaliplatin) and proteasome inhibitor bortezomib showed potent cytotoxicity not only against normal oral keratinocytes (14) but also against PC12 nerve-like cells (Iijima et al., unpublished data). The neurotoxicity of anticancer agents and X-ray irradiation is a recent research topic in today's aging society. In the present study, we investigated whether cytotoxic doses of X-ray irradiation further augment neurotoxicity induced by oxaliplatin and bortezomib. To perform this experiment, the anode heel effect of an X-ray tube that makes the cells on the anode side more susceptible to X-ray damage needs to be taken into consideration. Therefore, we first established a method which enabled the homogeneous irradiation of the cell population in each 96-microwell plate for anticancer drug treatment. This method, referred to as the ‘population irradiation method’, comprised five steps (Figure 1): (i) The preparation of a rat pheochromocytoma PC12 cell suspension; (ii) X-ray irradiation (Figure 2); (iii) inoculation and attachment in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS); (iv) induction of neuronal differentiation by nerve growth factor (NGF) and addition of anticancer drugs in serum-free method; and (v) viability assay (Figure 1).

Population irradiation method used for the X-ray irradiation and induction of differentiation of PC12 cells. A: Five steps for measuring X-ray-induced cytotoxicity. B: Morphological changed induced by nerve growth factor (NGF; 50 ng/ml) in serum-free medium. DMEM: Dulbecco's modified Eagle's medium; FBS: fetal bovine serum; MTT: 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide.

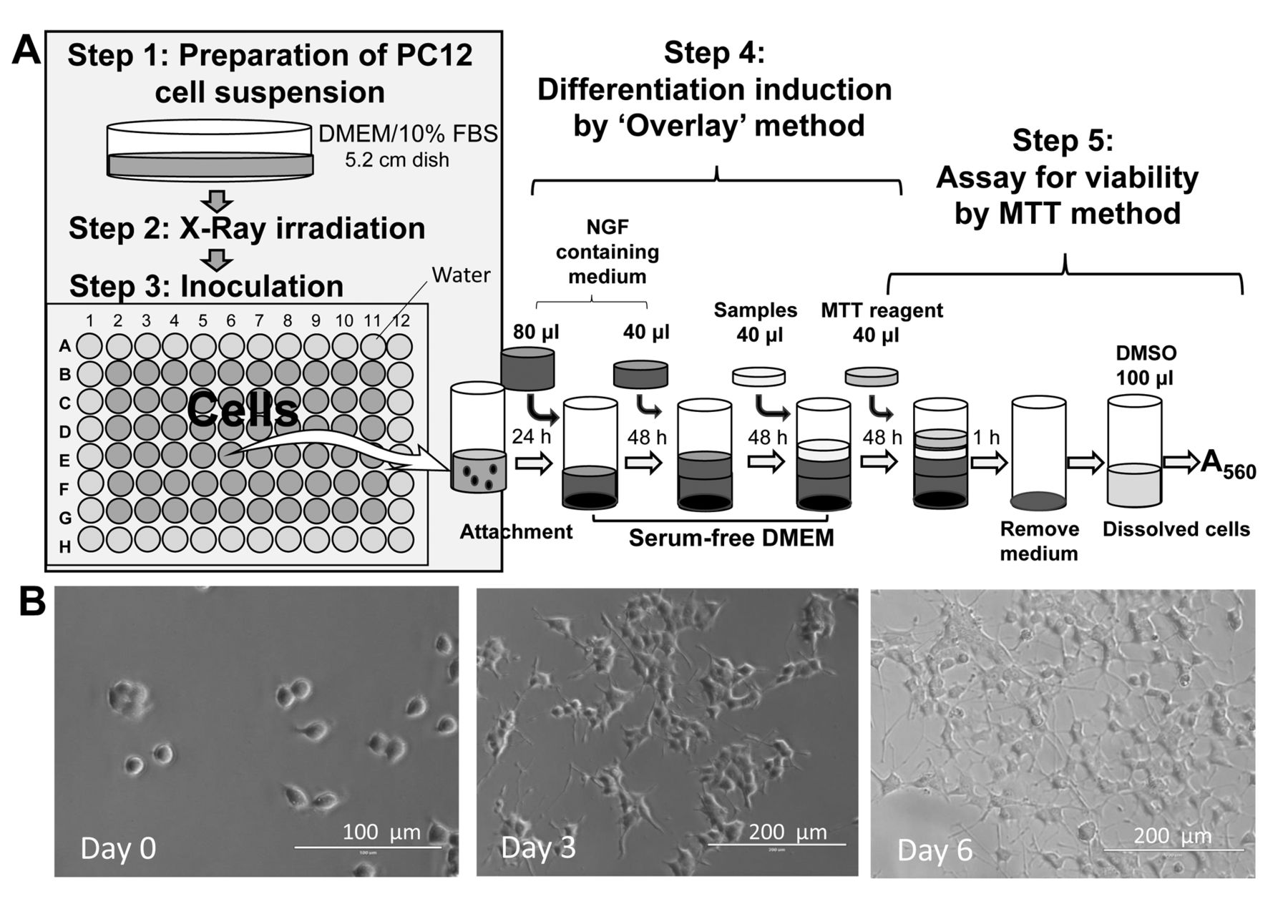
A: X-Ray apparatus and B: skin exposure dosimeter used in this study.

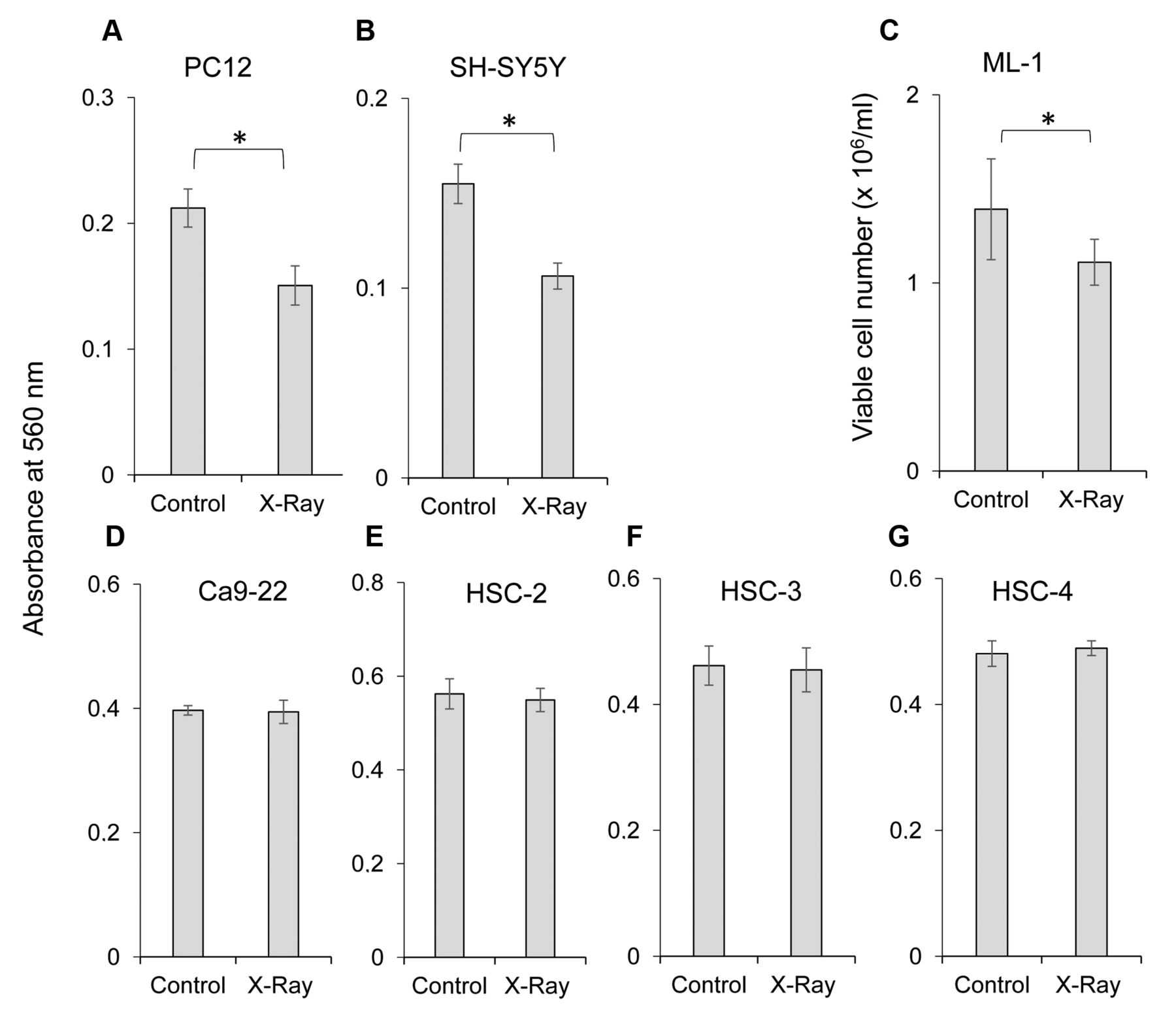
Comparison of X-ray sensitivity among several cultured cell lines. Near-confluent cells were exposed to 0 (control) or 505 mGy X-ray in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS), and then incubated for 48 h without changing the medium. The viable cell number was then determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide method and expressed as a percentage that of the control. Each value represents the mean±S.D. of six determinations. *Significantly different at p<0.05.
Materials and Methods
Materials. The following chemicals and reagents were obtained from the indicated companies: DMEM from GIBCO BRL (Grand Island, NY, USA); human recombinant NGF, chlorogenic acid and dimethyl sulfoxide from Wako Pure Chem. Ind. (Osaka, Japan); FBS and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) from Sigma-Aldrich Inc. (St. Louis, MO, USA); sodium ascorbate and N-acetyl-L-cysteine (NAC) from Tokyo Chemical Industry Co., Ltd (Tokyo, Japan); bortezomib from Janssen Pharmaceutical K.K. (Tokyo, Japan); oxaliplatin from Yakult Honsha Co., Ltd. (Tokyo, Japan); and culture plastic dishes and 96-well plates from Techno Plastic Products AG (Trasadingen, Switzerland).
Cell culture. PC12, a rat pheochromocytoma cell line derived from adrenal medulla; SH-SY5Y, a human neuroblastoma cell line cloned from a human bone marrow biopsy-derived line called SK-N-SH; human myeloblastic leukemia (ML-1); and human oral squamous cell carcinoma (OSCC) cell lines [Ca9-22 (derived from gingival tissue), and HSC-2, HSC-3, and HSC-4 (derived from tongue)] were purchased from Riken Cell Bank (Tsukuba, Japan). PC12, SH-SY5Y (15), Ca9-22, HSC-2, HSC-3 and HSC-4 cells (14) were grown at 37°C in DMEM supplemented with 10% heat-inactivated FBS, 100 units/ml, penicillin G and 100 μg/ml streptomycin under a humidified atmosphere with 5% CO2, while ML-1 cells were cultured in RPMI-1640 supplemented with 10% FBS and antibiotics (16). These cell lines were used for the comparison of X-ray sensitivity.

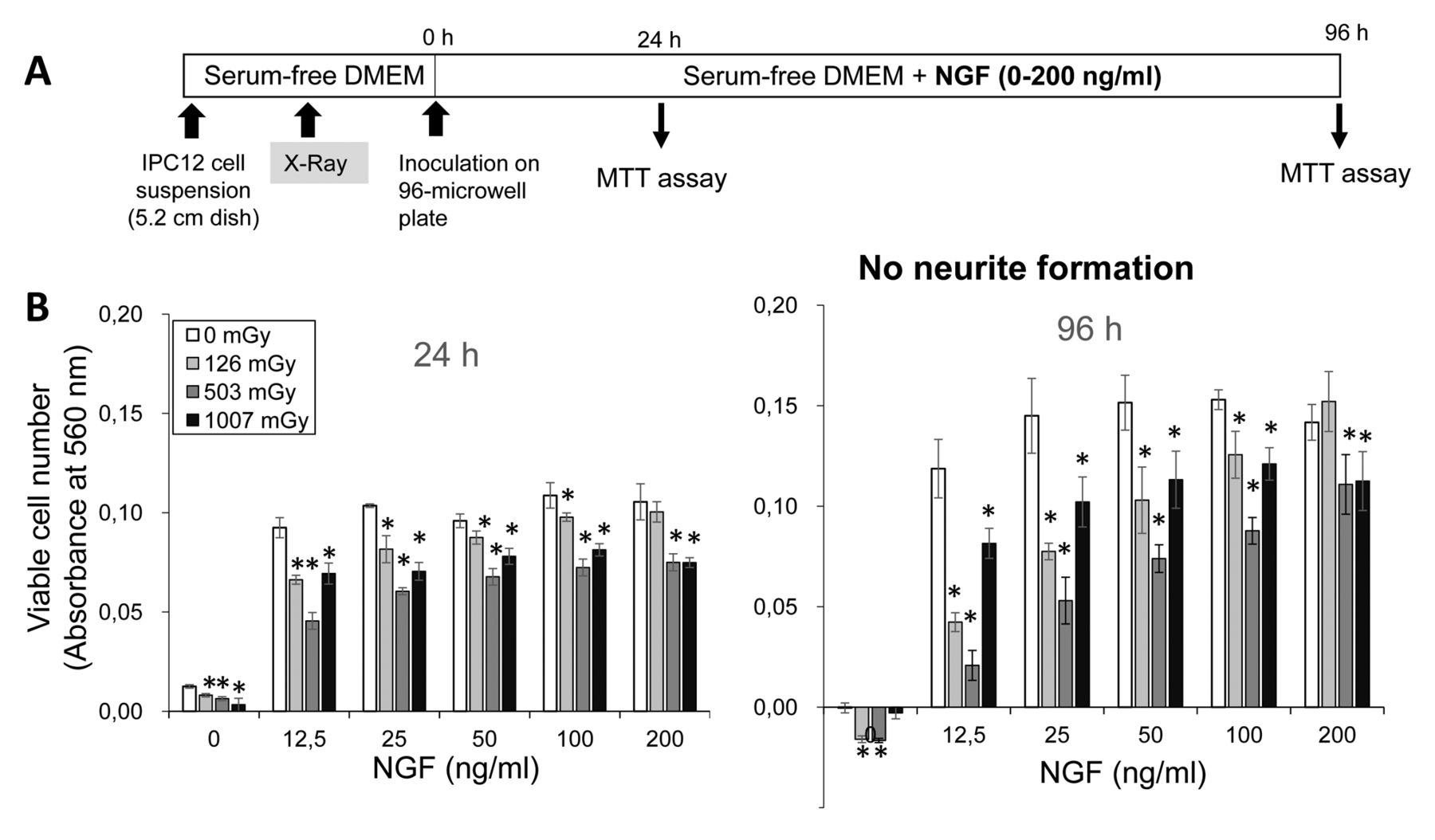
Protection by nerve growth factor (NGF) against X-ray-induced damage of PC12 cells. A: Experimental scheme. B: PC12 cells were seeded onto four 5.2 cm diameter dish [105 cells/5 ml of serum-free Dulbecco's modified Eagle's medium (DMEM)] and exposed to X-ray irradiation (0, 126, 503 or 1007 mGy). Immediately after the addition of 12.5, 25. 50, 100 or 200 ng/ml NGF, cells were incubated for 24 or 96 h in serum-free medium and the viable cell number was then determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) method. Each value represents the mean±S.D. of five determinations. *Significantly different from the control at p<0.0167.
Induction of neuronal differentiation. PC12 cells were differentiated into maturing neuronal cells by the overlay method established in our laboratory (17). In brief, cultured PC12 cells were detached by trypsin, and the cell pellet after centrifugation was resuspended in serum-free or FBS (10%)-supplemented DMEM. When cells were resuspended in serum-free medium, they were exposed to X-ray irradiation at 0 (control), 126, 503 or 1007 mGy, inoculated into 96-microwell plates, and then 0 (control), 12.5, 25, 50, 100 or 200 ng/ml NGF was added in quintuplicate. When cells were resuspended in DMEM supplemented with 10% FBS, they were exposed to X-ray irradiation at 0 (control) or 506 mGy and incubated for 24 h for cell attachment. The culture medium was then replaced with serum-free medium containing 50 ng/ml NGF (differentiation medium) and incubated for 72, 96, 120 or 144 h with occasional NGF overlay in triplicate. Plates were not precoated with collagen-I, collagen-IV, nor with poly-L-lysine, since the overlay method reproducibly yielded the highest number of differentiated cells having characteristic neurites, possibly due to the endogenous production of survival and differentiation-inducing factor(s) (17). The extent of differentiation was assessed under light microscopy (EVOSfl; ThermoFisher Scientific, Waltham, MA, USA) 77, 96, 120 or 144 h after NGF addition (18).
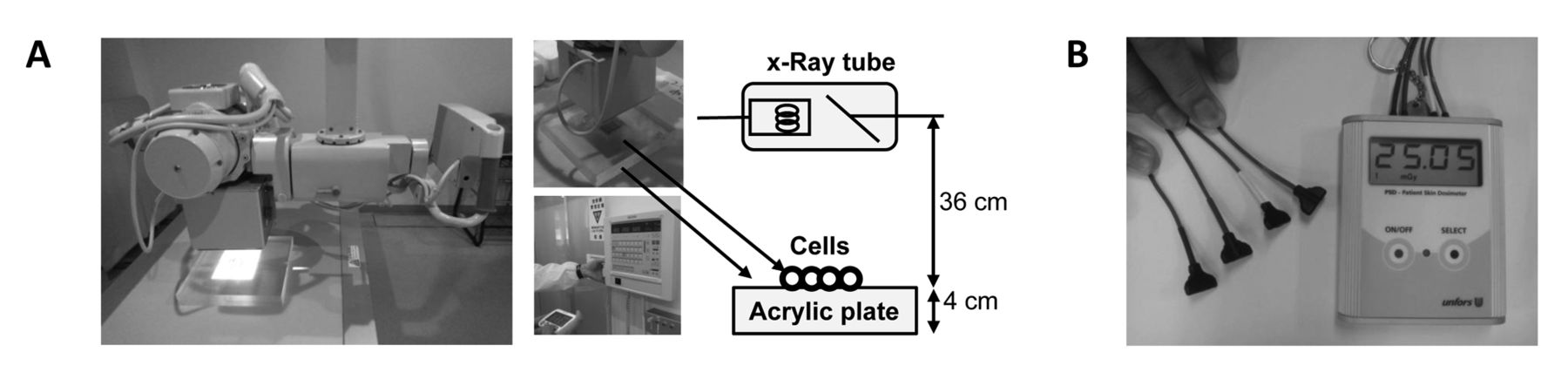
X-Ray irradiation. X-Ray irradiation was performed using a diagnostic X-ray apparatus (UD150L-30W; SHIMAZU, Kyoto, Japan) (Figure 2A). Culture plastic dishes or 96-well plates that contained cells were placed at the cathode side of an X-ray tube (SHIMAZU, P38DE 80s) on an acrylic plate (4 cm high) and irradiated with X-rays for 4, 8, 16 or 32 cycles of 0.2 s at 200 mA and 100 kV (corresponding to 126, 503 or 506 and 1007 mGy, respectively). Dosimetry was performed with a patient skin exposure dosimeter (Unfors Instruments Inc., New Milford, CT, USA) (Figure 2B). The measured value of the dose of X-ray absorbed by the cells was corrected by multiplying the value obtained by exposure dosimeter with the correction factor.
Assay for viable cell number. The viable cell number of undifferentiated PC12 cells, OSCC and ML-1 cells were determined by removing the medium and then incubating for 2 h with 0.2 mg/ml MTT in fresh DMEM. The viability of differentiated PC12 cells were determined by overlaying a final concentration of 0.1 mg/ml MTT reagent (Figure 1) and incubating for 2 h. The MTT-containing medium was then removed, and the purple crystals were lysed with 0.1 ml of dimethyl sulfoxide. The relative viable cell number was determined from the absorbance of the cell lysate at 560 nm, using a microplate reader (Infinite F50R; TECAN, Männedorf, Switzerland) (14).
In further experiments, the concentration of antioxidants (sodium ascorbate, NAC, chlorogenic acid) and anticancer drugs (oxaliplatin, bortezomib) that reduced the viable cell number by 50% (CC50) was determined from the dose–response curve in triplicate to sextuplicate.
Statistical treatment. Each value represents the mean±S.D. of 3-6 determinants. The statistical differences between control and X-ray-irradiated groups were evaluated by paired or unpaired t-test. A value of p<0.05 was considered to be significant. Significance levels were recalculated by Bonferroni correction when X-ray-irradiated groups differed in radiation doses. Control and X-ray-irradiated groups were compared at each NGF concentration.

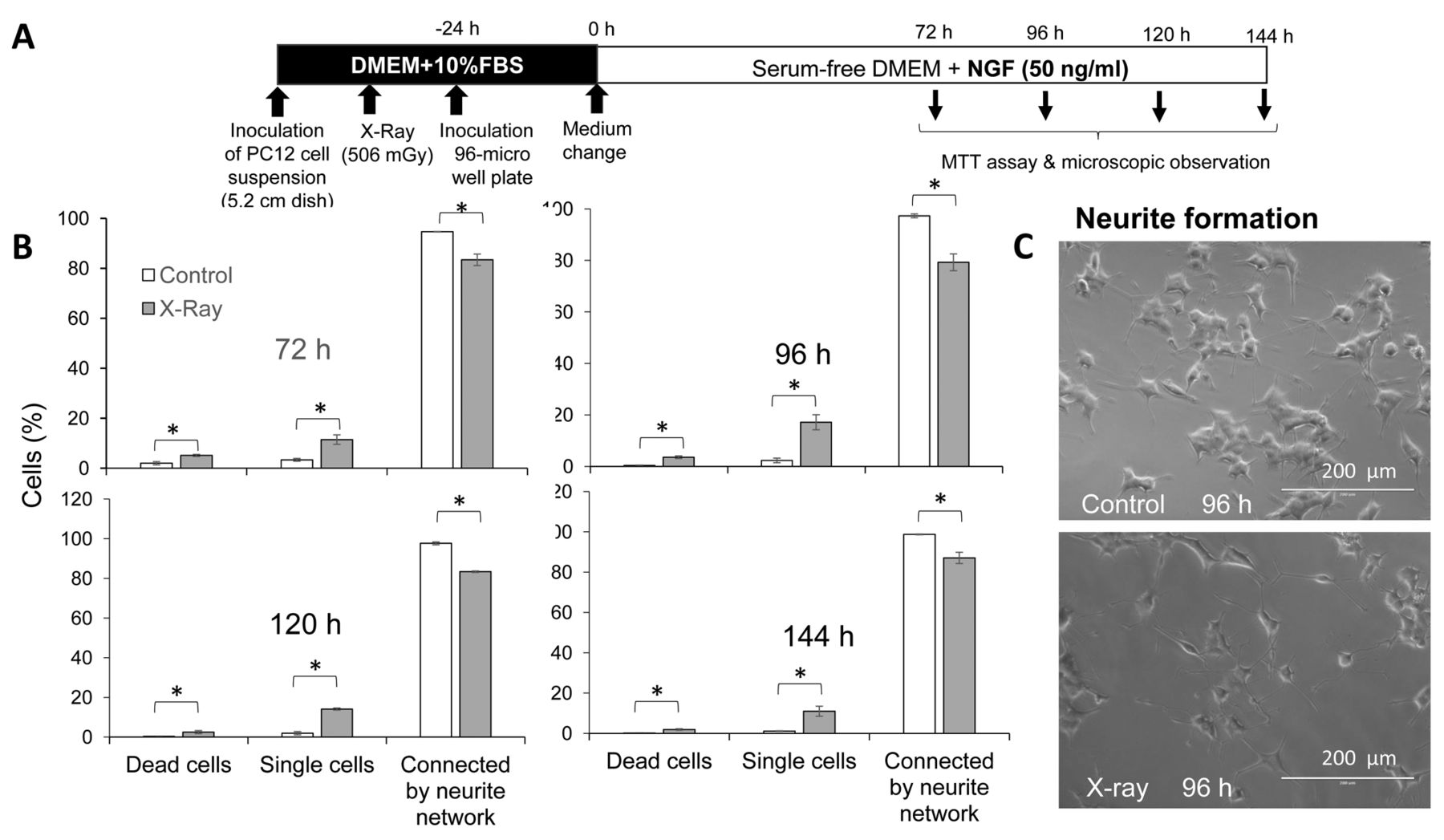
Disruption of connected neurite network by X-irradiation. A: Experimental scheme. B: PC12 cells were exposed to X-ray irradiation, inoculated and attached to 96-microwell plates, and incubated for 24 h to achieve cell attachment in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS). Culture medium was replaced with fresh serum-free medium containing 50 ng/ml nerve growth factor (NGF) and cells were further incubated for 72, 96, 120 or 144 h, and subjected to morphological observation. More than 400 cells were scored for classifying them as dead, single or cells connected by neurite network. Each value represents the mean±S.D. of three determinations. C: Representative cell morphology of control and X-ray-irradiated cells. MTT: 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide. *Significantly different at p<0.05.
Results
High X-ray sensitivity of neuronal cells. Cells grown in 96-microwell plates were irradiated by X-ray. When seven cell lines were subjected to X-ray irradiation (505 mGy), the viability of nerve precursor cells (rat pheochromocytoma cell line PC12, human neuroblastoma cell line SH-SY5Y) (Figure 3A and B) and human myeloblastic leukemia cells (ML-1) (Figure 3C) were significantly reduced, whereas those of the four OSCC cell lines (Ca9-22, HSC-2, HSC-3, HSC-4) were not affected (Figure 3D-G). Irradiation reduced the viable cell number of PC12, SH-SY5Y and ML-1 cells by 29%, 31% and 19%, respectively (Figure 3A-C). Subsequent experiments used cells that had been pre-irradiated by X-ray using the ‘population irradiation method’ to provide homogeneously irradiated cells.
X-Ray-induced cytotoxicity in PC12 cells by NGF. When PC12 cells were incubated with serum-free medium (Figure 4A), the cell growth stopped within 24 h. Addition of NGF stimulated cell growth (Figure 4B), confirming our previous report (17). X-Ray irradiation before NGF addition reduced the viable cell number in most combinations (Figure 4B). The viable cell number of X-ray-treated (126 mGy) cells with NGF (200 ng/ml) was not significantly different from that of the control. It was unexpected that there was no neurite growth (data not shown). This problem was solved by the addition of FBS (10%) during the cell-attachment period (Figure 5A), as judged from the presence of elongating neurites (Figure 5B and C).
Effect of X-ray irradiation on neurotoxicity of antioxidant and anticancer drugs by X-ray irradiation. X-Ray irradiation produced significantly higher numbers of dead and single cells and lower degree of neurite network formation (Figure 5B). Antioxidants (sodium ascorbate, NAC, chlorogenic acid) and anticancer drugs (oxaliplatin, bortezomib) showed cytotoxicity against differentiating PC12 cells with CC50 values of 0.36 mM, 7.9 mM, 0.43 mM, 4.3 μg/ml and 0.45 ng/ml, respectively (Figure 6B). X-Ray irradiation at 506 mGy reduced cell viability by 29%, 22%, 33%, 27% and 27%, respectively. Combination of X-ray irradiation with sodium ascorbate, chlorogenic acid, oxaliplatin or bortezomib slightly reduced the CC50 value to 0.27 mM, 0.35 mM, 1.4 μg/ml and 0.42 ng/ml, inducing an increase of neurotoxicity by 1.3-, 1.2-, 3.1- and 1.1-fold, respectively.
Augmentation of neurotoxicity of X-ray irradiation by antioxidants and anticancer drugs. A: Experimental scheme. B: PC12 cells were first exposed to X-ray irradiation (500 mGy) and induced to differentiated into nerve-like cells by NGF, and then treated with antioxidants sodium ascorbate, N-acetyl-L-cysteine (NAC), or chlorogenic acid, or anticancer drugs oxaliplatin, or bortezomib. Viable cells were then determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) method. Each value represents the mean±S.D. of six determinations. DMEM: Dulbecco's modified Eagle's medium; FBS: fetal bovine serum; NGF: nerve growth factor.
Sensitivity of X-ray-irradiated cells to anticancer drugs. Data of control and X-ray-irradiated differentiated PC12 cells were derived from Figure 6. Data of oral squamous cell carcinoma (OSCC) and human normal oral cells [human gingival fibroblast (HGF), human periodontal ligament fibroblast (HPLF), human pulp cell (HPC)] were derived from our recent article (14).
Discussion
In this study, we first established the optimal conditions for X-ray irradiation of differentiating PC12 cells. We selected PC12 cells as a model system since (i) the viability of this cell line and SH-SY5Y (another popular cell line used in neuroscience) were significantly reduced by X-ray irradiation (Figure 3), and (ii) this cell line can be differentiated by NGF into nerve-like cells characterized by neurites (17). We found that the optimal dose of X-ray irradiation was 0.5 Gy, nearly two orders lower than that used for cancer treatment of papillary tumor (45 to 60 Gy for 3D conformal radiation therapy and from 12 to 36 Gy for stereotactic radiosurgery) (19) and locally advanced cervical cancer (45 to 50.4 Gy) (20). NGF stimulated PC12 cell growth in serum-free medium, however, X-ray irradiation reduced the viable cell number under most combinations of irradiation dose and NGF concentration, even in the presence of NGF (Figure 4). It was unexpected that FBS needed to be present during the cell-attachment period (24 h) after X-ray irradiation. Cell attachment in the absence of FBS completely inhibited the formation of neurites by an as yet unidentified mechanism. It remains to be investigated whether NGF stimulates the growth of the cells remaining after X-ray irradiation or inhibits cell death induced by X-ray irradiation.
The present study demonstrated that X-ray irradiation significantly reduced the viability of two neuronal cell lines (undifferentiated PC12, and SH-SY5Y) and human myeloblastic leukemia cell line ML-1, but not that of four OSCC cell lines (Ca9-22, HSC-2, HSC-3, HSC-4) (Figure 3). When we compared these with our previous data (14), differentiated PC12 cells showed much higher sensitivity to two popular anticancer drugs (oxaliplatin, bortezomib) as compared with four OSCC cell lines and human normal oral cells (Table I). The sensitivity of differentiated PC12 cells against oxaliplatin was calculated to be 1.0- and 6.8-fold higher than that of OSCC and normal cells, respectively. Similarly, the sensitivity of differentiated PC12 cells against bortezomib was 2.4- and 1464-fold higher than that of OSCC and normal cells, respectively (Table I). The sensitivity of differentiated PC12 cells was further augmented by X-ray treatment. This suggests the necessity of cautional use of the combination of X-ray irradiation and classical anticancer drug (oxaliplatin) and proteasome inhibitor (bortezomib) for the treatment of cancer.
We found that X-rays did not in fact enhance but rather reduced the cytotoxicity of NAC. This may be due to the relatively lower cytotoxicity of NAC, necessitating the use of a higher concentration to induce cytotoxicity. This clearly reduced the pH in the culture medium due to its acidic property, possibly having affected the cellular metabolism.
It remains to be investigated whether reactive oxygen species may be involved in the combinational treatment of X-ray and radiotherapy (21, 22). We also found that the cytotoxicity of X-ray irradiation was also enhanced by sodium ascorbate and chlorogenic acid, well-known antioxidants. This suggests a common mechanism of the augmentation of anticancer activity of X-rays, regardless of the accompanying compound.
Acknowledgements
This work was partially supported by KAKENHI from the Japan Society for the Promotion of Science (JSPS) (16K11519) and research fund from Nihon Institute of Medical Science (Ogawa).
Footnotes
Authors' Contributions
G.N., N.T., H.T established X-ray irradiation method and protocol. H.S., Y.I., Y.K-O., T.N., D.U., A.S and A. D-G performed X-ray irradiation and cytotoxicity assay and data analysis. S.N., K.K. and R.G-C. prepared samples and designed the experiments. H.S., M.T. S.T., and S.K. wrote and edited the article.
This article is freely accessible online.
Conflicts of Interest
The Authors wish to confirm that there are no known conflicts of interest associated with this publication and there has been no significant financial support for this work that could have influenced its outcome.
- Received January 10, 2020.
- Revision received January 28, 2020.
- Accepted January 30, 2020.
- Copyright© 2020, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}