


Abstract
Background/Aim: Breast cancer is the most common malignant tumor among women worldwide. In previous work, we presented results of physical activity in primary prevention in a model of induced mammary gland cancer. In the present study, we assessed the influence of physical activity on sex hormone levels (estradiol and progesterone) and the expression of their receptors (ER, PR), as well as the level of apoptosis of tumor cells in secondary prevention. Materials and Methods: Fifty 1-month-old female Sprague-Dawley rats received intraperitoneal injection of 180 mg/kg body weight of N-methyl-N-nitrosourea (MNU) for tumor induction. Three months after the administration of MNU, rats were divided into four groups: low-intensity, moderate-intensity, and high-intensity physical training groups (combined as PT) and a sedentary control (SC) group. Physical training was conducted 5 days per week with a three-position treadmill according to a precisely described protocol. The entire training was completed by 32 rats from which tissue and blood were collected for further analysis. Immunohistochemistry for ER and PR expression, terminal deoxynucleotidyl transferase dUTP nick-end labeling method for detection of apoptosis, and enzyme-linked fluorescent assay for detection of plasma hormone levels (estradiol and progesterone) were performed. Statistical analysis used p<0.05 as the significance level. Results: Significantly stronger expression of ER and PR was found in the SC in comparison to the PT group (p=0.035 and p=0.036, respectively). No statistically significant differences were found in estradiol or progesterone concentrations between SC and PT groups. Apoptosis was non-significantly increased in the PT group in comparison with the SC group. Stronger apoptosis in the PT group correlated positively with the level of training intensity (r=0.35, p=0.05). Conclusion: Physical training may reduce ER and PR expression in breast cancer cells, and reduce cell sensitivity to pro-proliferative and anti-apoptotic effects of estrogens, ultimately leading to apoptosis.
- Estrogen receptor
- progesterone receptor
- estradiol
- progesterone
- breast cancer
- physical activity
- N-methyl-N-nitrosourea
- rat experimental model
Breast cancer is the most common malignant tumor among women worldwide (30%), being the second leading cause of cancer-related death in the United States and in Europe (14%) (1). Nearly 20% of women worldwide are diagnosed at 30-49 years of age and over 40% of women aged 65 years or over (2).
Estrogens play an important role in the pathogenesis of breast cancer. Estradiol stimulates cell proliferation and differentiation by estrogen receptor (ER). Many reports indicate the relationship between exposure to estrogens and breast cancer. The risk of breast cancer is associated with increased exposure to endogenous and exogenous sex hormones in cases of an early age of menarche, late age of menopause, late first pregnancy (after 30 years of age) and hormone replacement therapy (3, 4). Moreover, risk increases with age and is the highest in the postmenopausal period, when the main source of estrogens is peripheral adipose tissue (3-5). Hence, hormonal imbalance and obesity are important risk factors for breast cancer (5-7).
Many reports suggest that physical activity is associated with a reduction in the risk of breast cancer and its recurrence. Physical activity can reduce the body weight, the amount of fat tissue and endogenous conversion to estrogens (8). Regular physical training leads to a decrease in estradiol and progesterone by reducing the ovarian secretory function, which can have a protective role against breast cancer (9). In this field contrary observations may be found. McTiernan et al. showed a decrease in estrogen level and an increase in serum level of sex hormone binding globulin as a result of physical exercise (10), while Atkinson et al. did not confirm the expected changes in estrogen metabolites as a consequence of physical activity (11). Further studies are strongly needed and an experimental rat model with tumors induced by N-methyl-N-nitrosourea (MNU) due to the similarity of the structure and function with a human mammary gland seems to be reliable and relevant (12).
The aim of our project entitled “Impact of Physical Training on the Carcinogenesis and Progression of Rat Mammary Glands” was the assessment of the impact of physical training on the course of cancer, depending on the intensity of exercise training and the examined prevention model. The results related to primary prevention were presented by Malicka et al. (13), whereas initial results related to secondary prevention were presented by Siewierska et al. (14). In the present article, we report the assessment of the influence of physical activity on sex hormone levels (estradiol and progesterone) and the expression of their receptors (ER, PR), as well as the level of apoptosis of tumor cells in the model of secondary prevention.
Materials and Methods
Animals. For the whole project, fifty 1-month-old female Sprague-Dawley rats obtained from the Medical University of Silesia (Katowice, Poland) were used. All the procedures, e.g. housing, feeding and tumor induction [intraperitoneal injection of 180 mg/kg body weight of MNU (Sigma-Aldrich, Germany)] were performed in accordance with the protocol described by Malicka et al. (13). The project met the European Union standards and was approved by the Institute of Immunology and Experimental Therapy of the Polish Academy of Sciences (consent number 37/2010).
Physical training. Three months after the administration of MNU, rats were divided into four groups, i.e.: low-intensity (n=12), moderate-intensity (n=12), high-intensity (n=12) physical training groups (PT); and a sedentary control group (SC; n=14). Physical training was conducted 5 days per week with a three-position treadmill (Exer 3/6; Columbus Instruments, Columbus, OH, USA) according to the previously described protocol (9). The speed and the duration of training on the treadmill were gradually increased depending on the group's defined level of intensity (Table I). The parameters of the low-intensity group were 20% lower and those of the high-intensity group were 20% higher than those of the moderate-intensity group. The entire training was completed by 32 rats.
Protocol of moderate-intensity training.
Tissue collection. Six months following MNU administration, the animals were sacrificed (13, 14). All tumors in the animals that were detected on palpation were collected and measured. The obtained tissues were fixed in 4% buffered formalin and then embedded in paraffin blocks. Subsequently, for future immunohistochemistry assays, tissue microarrays (TMAs) were prepared using a 2-mm-gauge needle and Manual Tissue Arrayer I (Beecher Instruments Inc., Sun Prairie, WI, USA). Moreover, for biochemical assays, the blood was collected, centrifuged (500×g for 15 min at 4°C) and the obtained plasma was frozen.
Immunohistochemistry. All reactions were performed on 4-μm-thick TMA sections in an automated Autostainer Link48 staining platform. Deparaffinization, rehydratation, and antigen retrieval were performed by boiling the sections in Target Retrieval Solution buffer using a Pre-Treatment Link Platform. The sections were incubated with the EnVision FLEX Peroxidase-Blocking Reagent to block the endogenous peroxidase activity. Subsequently, the sections were incubated with the polyclonal rabbit anti-rat primary antibodies directed against ERα (1:100; Novus Biologicals, Abingdon, UK) and PR (1:100; Novus Biologicals). The sections were then incubated (20 min at room temperature; RT) with EnVision FLEX Horseradish Peroxidase-conjugated secondary antibodies. The substrate for peroxidase, diaminobenzidine, was then applied and the sections were incubated at RT for 10 min. Finally, the sections were rinsed and counterstained with Mayer's hematoxylin and mounted using SUB-X Mounting Medium. All the reagents, except primary antibodies, and equipment used for the reactions were provided by Dako (Glostrup, Denmark).
Terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) method. The apoptosis assay was conducted using ApopTag in situ Apoptosis Detection Kit (Millipore, Billerica, MA, USA). TMA sections were dewaxed in xylene, rehydrated in ethanol and rinsed with distilled water and phosphate-buffered saline. The sections were digested using Proteinase K (Dako) for 5 min at RT. The activity of endogenous peroxidase was blocked by 5-min incubation in 3% H2O2 in phosphate-buffered saline. The sections were incubated, first with Equilibration Buffer for 10 min at RT and then with TdT Enzyme and Reaction Buffer at 37°C for 1 h. The reaction was terminated by incubation with Stop Buffer for 10 min at RT. Subsequently, the sections were incubated with anti-digoxigenin peroxidase-conjugated secondary antibodies and diaminobenzidine was applied for 10 min at RT. Finally, the sections were counterstained with Mayer's hematoxylin and mounted in SUB-X Mounting Medium (Dako).

Immunohistochemical (IHC) scores for nuclear expression of estrogen receptor (ER; A) and progesterone receptor (PR, B) in tumor cells of induced mammary gland cancer. Comparison of ER (C) and PR (D) expression and plasma level of estradiol (E) and progesterone (F) between SC and PT groups. The values are expressed as the mean±SD. *Significantly different at p<0.05.
Evaluation of the reactions. All TMA sections were evaluated under a BX-41 light microscope equipped with CellD software for computer-assisted image analysis (Olympus, Tokyo, Japan). For the evaluation of ER, PR and TUNEL reaction in the TMA sections, three fields with the highest number of tumor cells yielding a color reaction were selected (hot spots). The percentage of positively stained cells was evaluated by scoring labeled cells in relation to all cancer cells distinguished under ×400 magnification. The final score was the average of three hot-spot values for each case.
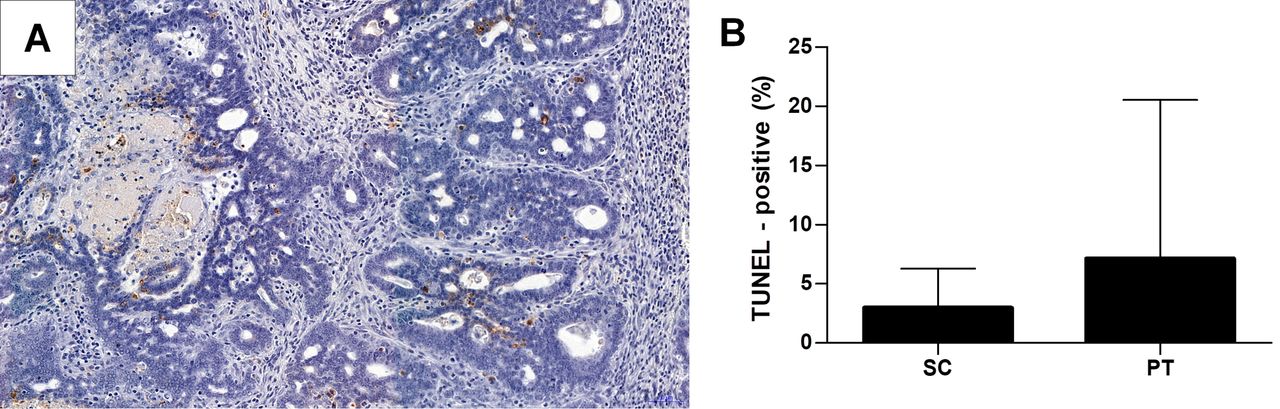
A: Visualization of cells undergoing apoptosis by terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) method. B: Comparison of apoptosis level between sedentary control (SC) and physical training (PT) groups. The values are expressed as the mean±SD.
Clinicomorphological parameters of the study animals.
Determination of plasma hormone levels. 17β-Estradiol and progesterone in serum were determined by enzyme-linked fluorescent assay (BioMerieux, Marcy-L'etoile, France), according to the manufacturer's recommendations. All steps were performed automatically by a VIDAS immunoanalyzer (BioMerieux), including the automated calculation of the results in relation to the programmed calibration curve.
Statistical analysis. Shapiro-Wilk, Levene, Spearman, Mann-Whitney and Kruskal-Wallis tests were used in our study. Statistical analysis was performed using Statistica 10.0 (Statsoft, Cracow, Poland) and Prism 7.0 (GraphPad, La Jolla, CA, USA). The results were considered statistically significant at p<0.05.
Results
Fifty rats started physical training, of which 32 finished it. Statistical analysis was performed only for those animals with induced tumors (seven rats in SC; 19 in PT). Due to the fact that the groups were small in size, rats were assessed as two groups, namely the PT group (regardless of training intensity) and the SC group. The expression of ER and PR was observed in nuclei of cancer cells (Figure 1A and B). Significantly stronger expression of ER (p=0.035, Figure 1C) and PR (p=0.036, Figure 1D) were found in the SC in comparison to the PT group. No statistically significant differences were found in estradiol or progesterone concentrations between the SC and PT groups (Figure 1E and F). Apoptosis was increased in the PT group, however, in comparison with the SC group, the difference was not statistically significant (Figure 2). Stronger apoptosis in the PT group positively correlated with the level of training intensity (r=0.35, p=0.05). There were no statistically significant differences in terms of described clinical features between individual groups at the beginning and at the end of the experiment (Table II).
Discussion
Many scientific reports indicate a protective effect of physical activity on breast cancer (8, 9, 13, 15-17), including in patients after comprehensive breast cancer treatment (8, 9, 18-24). From the results of our study, we found two mechanisms which confirm these observations, namely desensitization of cells due to reduced expression of ER and PR, as well as increased apoptosis which positively correlated with the level of training intensity.
The reduction in ovarian secretory activity that results in a decrease in sex hormone levels in the blood is one of the mechanisms that reduce the risk of breast cancer or its recurrence as a result of physical activity. Others suggest that physical training increases levels of sex hormone-binding globulin. In experimental studies Brown et al. showed that mammary cancer was statistically significantly less prevalent in physically active rats compared to less active controls (25). Brown et al. also reported that women exposed to high estradiol concentrations had an increased risk of breast cancer. After 12 months of exercise, levels of hormones circulating in the blood decreased. Further decrease was observed after reduction in body weight (25, 26).
According to another hypothesis, physical training does not significantly reduce levels of sex hormones but significantly reduces the expression of their receptors (9, 17, 27). A reduction in receptor expression results in the lack of cell stimulation by sex hormones, followed by cell apoptosis (17). We found statistically insignificant differences between the estradiol levels in PT vs. SC groups, in accordance with other authors (9, 15, 28).
Estradiol affects only cells showing ER expression, while the absence of ER prevents hormonal influence (27, 29-31). In addition, statistically significantly lower ER and PR IHC scores were found in the PT groups in our study. Wang et al. also reported reduced expression of ER as a result of physical training (32). However, Faustino-Roch et al. obtained different results (27).
Many authors point to the key role of estradiol in inducing apoptosis in breast cancer cells (33-35). Our study also revealed increased apoptosis in the training groups (although statistically insignificant) which was associated with reduced tumor volume (17, 36). Figueira et al. observed a positive effect of long-term moderate physical activity on the development of breast tumors and the balance between cell proliferation and apoptosis. Similarly, Westerlind et al. found statistically significantly higher apoptosis in physically trained rats compared to non-exercised animals (17). Coelingh et al. reported an anti-apoptotic effect of estrogens (37).
Based on the preliminary results, we conclude that physical training may reduce the expression of ER and PR by breast cancer cells. Moreover, it may reduce cell sensitivity to pro-proliferative and anti-apoptotic effects of estrogens, finally leading to their apoptosis. The findings obtained from this model of secondary prevention need further testing to overcome the main study limitation of a small number of cases and to strengthen this research hypothesis.
Acknowledgements
The Authors thank Bozena Przygodzka, Agnieszka Baranska and Elzbieta Polejko for their technical assistance. This work was funded by the grant of the Polish Ministry of Science NN404088240 “Impact of Physical Training on the Carcinogenesis and Progression of Rat Mammary Glands”.
Footnotes
Authors' Contributions
KS, IM, MPO, PD and MW conceived the study design. KS, IM, AP, UP and MC conducted the experiment. JG performed statistical analysis. KS, CK and JG analyzed the results. KS and IM performed literature reviews and drafted the article. CK revised the article. All Authors read and approved the final article.
This article is freely accessible online.
Conflicts of Interest
The Authors declare no conflicts of interest regarding this study.
- Received December 4, 2019.
- Revision received December 22, 2019.
- Accepted December 30, 2019.
- Copyright© 2020, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved




{kind=link}
{kind=link}