


Abstract
Background/Aim: Evidence has indicated that fisetin induces cytotoxic effects in human cancer cell lines, including the inhibition of cell migration and invasion, however, the exact molecular mechanism of action of fisetin in human osteosarcoma cells remains unclear. Materials and Methods: The anti-metastatic mechanisms of fisetin in human osteosarcoma U-2 OS cells were investigated in vitro. Results: Fisetin reduced the viability of cells at different concentrations (2.5, 5 and 10 μM) as measured by flow cytometric assay. Fisetin suppressed cell mobility, migration and invasion of U-2 OS cells, as shown by wound healing assay and transwell filter chambers, respectively. The gelatin zymography assay showed that fisetin inhibited MMP-2 activity in U-2 OS cells. Results from western blotting indicated that fisetin reduced the levels of pEGFR, SOS-1, GRB2, Ras, PKC, p-ERK1/2, p-JNK, p-p-38, VEGF, FAK, RhoA, PI3K, p-AKT, NF-ĸB, uPA, MMP-7, MMP-9, and MMP-13, but increased GSK3β and E-cadherin in U-2 OS cells after 48 h of treatment. Conclusion: Fisetin can be used in the future, as a target for the treatment of metastasis of human osteosarcoma cells.
Osteosarcoma (OS) has been recognized to be one of the most common malignant bone tumors, which occurs frequently in children and adolescents and exhibits high destructive and metastatic potential (1, 2). OS has been reported to have an increased tendency to metastasize (1, 3, 4). In the past 30 years, the clinical outcome for OS patients with metastatic tumors has remained unchanged, thus, development of new therapeutic strategies for OS patients is urgent (5). Evidence has revealed that the regulation of oncogenes and tumor suppressor genes is vital in the development, progression and metastasis of OS (6-8).
Metastasis, the most important characteristic of malignant tumors, is a multistep process initiated by local invasion, plays a critical role in treatment efficacy and quality of life in patients with metastatic tumors (9) and accounts for more than 90% of cancer related deaths (10, 11).
A vital step in the process of metastasis is the degradation of the extracellular matrix (ECM) by matrix metalloproteinases (MMPs) (12). The MMP system includes MMPs, tissue inhibitor of metalloproteinases (TIMPs) and urokinase-type plasminogen activator (uPA). Furthermore, MMP upregulation promotes cancer metastasis (13-15). The inhibition of molecular pathways involved in cancer metastasis could be a potential strategy to inhibit metastasis (16, 17).
Currently, many chemotherapeutic drugs used in cancer patients are derived from natural sources (18). Fisetin, a dietary tetrahydroxyflavone, is found in natural plants including many fruits and vegetables. It has also been shown to have many biological activities, including inhibiting different types of cancer via the inhibition of multiple oncogenic pathways both in vitro and in vivo (19-21). In our earlier studies, fisetin induced cancer cell apoptosis of human oral cancer SCC-4 and HSC-3 cells by causing endoplasmic reticulum stress, and via caspase- and mitochondria-mediated signaling pathways (22, 23). Fisetin has also been shown to inhibit cell migration and invasion of Epstein-Barr virus latent membrane protein-1 (LMP1)-positive nasopharyngeal carcinoma (NPC) cells (24). Fisetin inhibited cell migration and invasion of cervical cancer cells by repressing uPA via interruption of the p38 MAPK-dependent NF-ĸB signaling pathway (25). Recently, it was also reported that fisetin inhibited the cell growth and migration of human lung cancer A549 cells by blocking the ERK1/2 pathway (26). However, there is no available information showing the effect of fisetin in the cell migration and invasion of human osteosarcoma cells, which was therefore the aim of the present study. The results showed that fisetin had a significant anti-migratory and anti-invasive effect on human osteosarcoma U-2 OS cells in vitro. These findings offer a deeper understanding of the anti-metastatic mechanisms of fisetin in osteosarcoma.
Materials and Methods
Test chemicals, reagents and culture medium. Fisetin, dimethyl sulfoxide (DMSO), Tris-HCl, trypan blue, trypsin, propidium iodide (PI), gelatin and Coomassie blue R-250 were purchased from Sigma Chemical Co. (St. Louis, MO, USA). McCoy's 5A medium, fetal bovine serum (FBS) and penicillin-streptomycin were purchased from Gibco®/Invitrogen Life Technologies (Carlsbad, CA, USA). Antibodies specific for MMP-7, MMP-9 and MMP-13 were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA) and antibodies for Ras, GRB2, Sos1, uPA, PKC, p-EGFRTyr1068, p-ERK1/2, p-JNK, p-P38, VEGF, FAK, Rho A, PI3K, p-AktTHr308, NF-ĸB, GSK3β, Snail, E-cadherin, β-catenin, N-cadherin, Vimentin, p-AktSer473, and peroxidase conjugated secondary antibodies were purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA). Fisetin was dissolved in DMSO (carrier solvent) and 0.5% DMSO was used in control groups. Fisetin was further diluted in culture medium to reach the appropriate final concentrations.

Fisetin decreased cell viability of U-2 OS cells. Cells (1×105 cells/well) were incubated with fisetin (0, 2.5, 5, 10, 20 and 40 μM) for 48 h. Cells were collected for measurement of the percentage of total viable cells as described in Materials and Methods. *p<0.05, **p<0.01, ***p<0.001, significant difference between fisetin-treated groups and the control as analyzed by one-way ANOVA.
Cell line and culture. Human osteosarcoma U-2 OS cell line was obtained from the Food Industry Research and Development Institute (Hsinchu, Taiwan). U-2 OS cells were cultured in McCoy's 5A medium supplemented with 10% FBS, 2 mM L-glutamine, 10 g/l non-essential amino acids, 100 μg/ml streptomycin and 100 units/ml penicillin at 37°C in a humidified atmosphere containing a 5% CO2.
Cell viability assay. U-2 OS cells (1×105 cells/well) were placed onto 12-well plates with McCoy's 5A medium and were incubated with fisetin at the final concentrations 0, 2.5, 5, 10, 20 and 40 μM, in triplicate for 48 h. At the end of the incubation, cells were harvested, washed, counted and stained with PI (5 μg/ml) to measure the percentage of cell viability by using flow cytometry (Becton-Dickinson, San Jose, CA, USA) as previously described (27).
In vitro scratch wound healing assay for cell mobility. Scratch wound healing assay was used to examine cell mobility characteristics as previously described (28). Briefly, U-2 OS cells (1×105 cells/well) were grown in a 12-well plate until they reached a confluent monolayer. Medium was replaced with serum-free McCoy's 5A culture medium. Cell monolayers were scratched (wound) using a sterile 200 μl-pipette tip and PBS was used for washing and removing cell debris. Cells were incubated with various concentrations of fisetin (0, 2.5, 5 and 10 μM) for 24 h. In the denuded zone, the migrating cells were monitored and photographed under phase contrast microscopy and experiments were repeated three times. Image J software was used to quantify the relative wound size. Cell mobility inhibition (%)=new scratch width/original scratch width ×100% as previously described (28, 29).
Gelatin zymography assay for gelatinolytic activity. U-2 OS cells (1×105 cells/well) were placed in a 12-well plate for 24 h, after which medium was replaced with serum-free McCoy's 5A culture medium containing fisetin (0, 2.5, 5 and 10 μM) for 24 and 48 h. At the end of incubation, conditioned medium from each well was harvested and loaded onto 10% polyacrylamide gels and co-polymerized with 0.2% gelatin. Gel was soaked twice in 2.5% Triton X-100 in dH2O at 25°C for 30 min as previously described (28, 30). Then, gel was incubated in zymogen developing buffer (Sigma Aldrich), containing 50 mM Tris (pH 7.5), 200 mM NaCl, 5 mM CaCl2, 1 μM ZnCl2 and 0.02% Brij 35, overnight at 37°C. Band corresponding to MMP-2 activity was stained with 0.2% Coomassie blue in 10% acetic acid and 50% methanol and photographed. The band of gelatinolytic activity was measured using NIH Image J software, version 1.47 (National Institutes of Health, Bethesda, MA, USA), as previously described (30, 31).

Fisetin affected in vitro wound closure of U-2 OS cells. Cells (1×105 cells/well) were kept in a 12-well plate for 24 h, scratched (wounded), and incubated with fisetin (0, 2.5, 5 and 10 μM) for 24 h. The relative wound closures were photographed using phase contrast microscopy (A) and percentage of wound area was calculated (B) as described in Materials and Methods. *p<0.05, **p<0.01, significant difference between fisetin-treated groups and the control as analyzed by one-way ANOVA.
Cell migration and invasion analyzed with the Transwell assay. Cell migration and invasion in vitro were examined by using Collagen and Matrigel assay system as previously described (28, 30). Briefly, U-2 OS cells (5×104 cells/well) in serum-free McCoy's 5A culture medium containing different concentrations of fisetin (0, 2.5, 5 and 10 μM) were placed in the upper chamber (transwell insert) (8 μm pore size; Millipore, Temecula, CA, USA) which was coated with 50 μl collagen (for cell migration examination) overnight. In the lower chamber, 800 μl of McCoy's medium with 10% FBS were placed for 48 h. The non-migrated cells found on the upper surface of the membrane were removed. The migrated cells (those adhered to the lower surface of the membrane) were fixed with 4% formaldehyde in PBS, treated with methanol, stained with 2% crystal violet and all samples were photographed under light microscopy. The percentage of cells that migrated were calculated. The cell invasion assay was performed similarly to the cell migration assay, except that the membrane of the insert (upper chamber) was covered with Matrigel (Matrigel: serum-free medium 1:9) (28, 30).
Western blotting analysis for cell metastasis-associated protein expression. U-2 OS cells (1×106 cells/dish) were placed in 10-cm culture dishes and incubated with fisetin (0, 2.5, 5 and 10 μM) for 48 h. At the end of incubation, cells were collected and resuspended in lysis buffer (50 mM Tris-HCl pH 7.5, 400 mM NaCl, 2 mM EGTA, 1 mM EDTA, 1 mM DTT, and protease inhibitor cocktail) (Roche). The cell lysates were centrifuged at 10,000 × g at 4°C for 10 min and total protein concentration was measured using Bradford protein assay kit as previously described (28, 30). Each sample (30 μg of total protein) was separated by 12% SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto PVDF membrane (Millipore, Bedford, MA, USA). All membranes were blocked with 5% non-fat milk in TBS-T buffer (10 mM Tris–HCl, 150 mM NaCl, and 0.05% Tween-20, pH 7.8) for 1 h at room temperature. After washing with TBS-T buffer, membranes were incubated with primary antibodies against MMP-7, MMP-9, MMP-13, p-EGFRTyr1068, Ras, GRB2, Sos1, uPA, PKC, p-ERK1/2, p-JNK, p-p38, VEGF, FAK, Rho A, PI3K, p-AktThr308, p-AktSer473, NF-ĸB, GSK3β, Snail, E-cadherin, β-catenin, N-cadherin, Vimentin. Membranes were incubated with the diluted corresponding HRP-conjugated secondary antibodies (diluted 1:5000; Cell Signaling Technology) and developed with ECL (Amersham). The Biospectrum Imaging System (UVP, Inc., Upland, CA, USA) was used to detect the corresponding bands as previously described (28, 32).


Fisetin inhibited MMP-2 activity and suppressed cell migration and invasion of U-2 OS cells in vitro. Cells (1×105 cells/well) were placed in a 12-well plate and treated with fisetin (0, 2.5, 5 and 10 μM) for 24 and 48 h and then conditioned medium was harvested for gelatin zymography assay, as described in Materials and Methods. (A) Representative gelatin gel pictures. Cells (5×104 cells/well) were placed on a transwell insert coated with collagen for migration or Matrigel for invasion, and were treated with fisetin (0, 2.5, 5 and 10 μM) for 48 h. U-2 OS cells penetrated the lower surface of the transwell membrane for migration (B) and invasion (C) were stained with crystal violet and were photographed under a light microscope at 200×. Penetrated cells were counted as described in Materials and Methods. *p<0.05, **p<0.01, ***p<0.001, significant difference between fisetin-treated groups and the control, as analyzed by one-way ANOVA.
Statistical analysis. Data are presented as mean±SD and were statistically analyzed by one-way ANOVA analysis of variance. *p<0.05, **p<0.01, ***p<0.001 were determined as significant.
Results
Fisetin decreased viability of U-2 OS cells. After treated with various concentrations of fisetin, total viable cell number was measured by flow cytometry. As indicated in Figure 1, fisetin at a concentration of 2.5-5 μM did not show morphological changes and only slightly reduced the percentage of viable cells after 48 h of treatment in U-2 OS cells. However, fisetin at 10 μM induced cell morphological changes and reduced the percentage (about reduced 10%) of viable cells when compared to control groups.
Fisetin inhibited cell mobility in U-2 OS cells. As indicated in Figure 2A and B, fisetin at 5-10 μM inhibited the closure rate of the scratch in U-2 OS cells. After 24 h of incubation, the control cells but not the fisetin-treated cells covered the opening of the scratch. The inhibited effect was dose-dependent, since at the high dose (10 μM), the edge distance was significantly longer, compared to that observed at the low dose (2.5 μM) (Figure 2B). Based on these observations, fisetin significantly suppresses the cell mobility of U-2 OS cells in vitro.

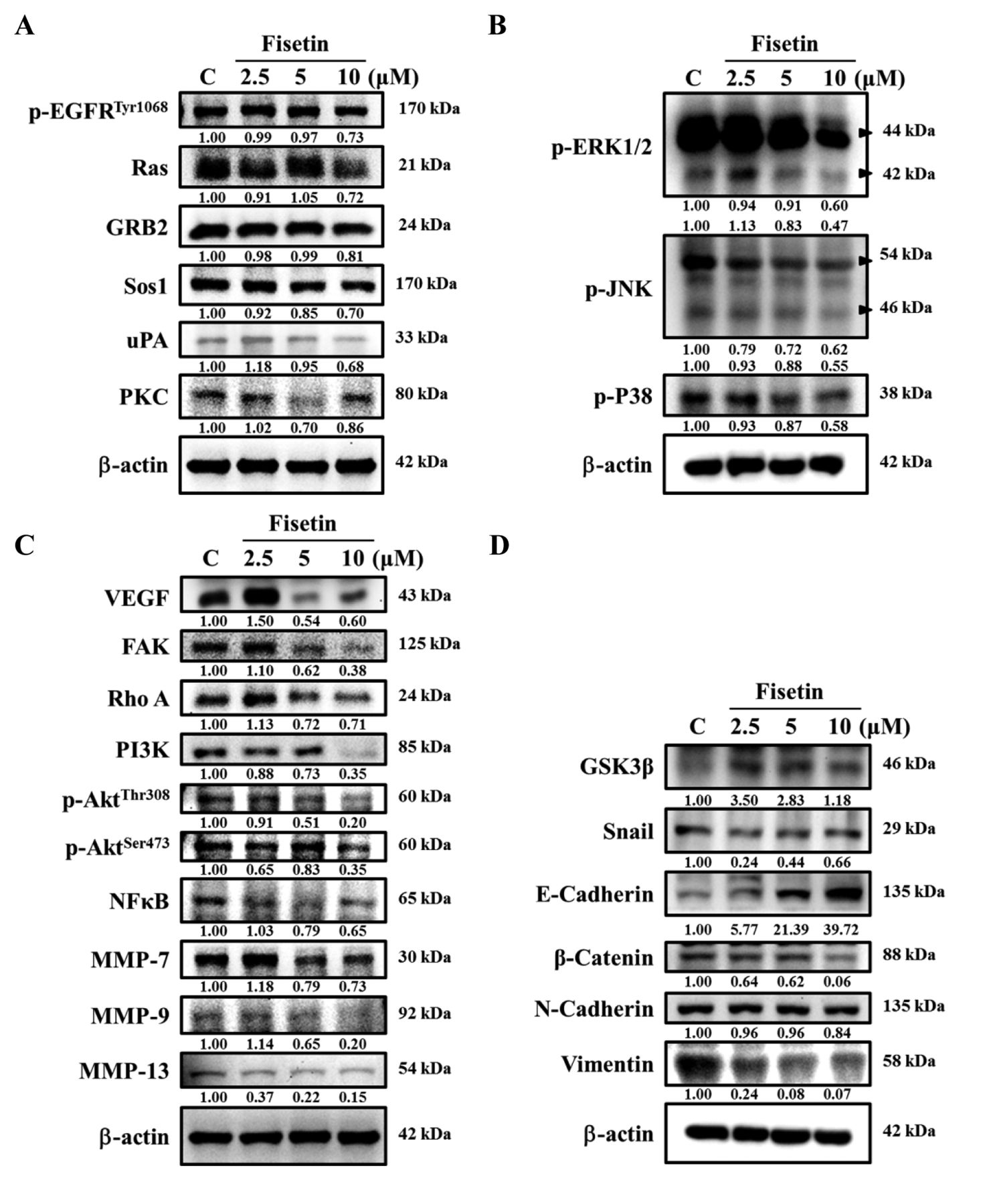
Fisetin affected the levels of proteins associated with migration and invasion of U-2 OS cells. Cells (1×106 cells/dish) were treated with fisetin (0, 2.5, 5 and 10 μM) for 48 h and collected. The proteins were separated by SDS-PAGE as described in the Materials and Methods. The levels of p-EGFRTyr1068, Ras, GRB2, Sos1, uPA and PKC (A); p-ERK1/2, p-JNK, and p-P38 (B); VEGF, FAK, Rho A, PI3K, p-AktThr308, p-AKTThr473, NF-ĸB, MMP-7, MMP-9 and MMP-13 (C); GSK3β, Snail, E-cadherin, β-catenin, N-Cadherin and Vimentin (D) expressions were estimated by western blotting as described in Materials and Methods.
Fisetin affected matrix metalloproteinase activity and cell migration and invasion in U-2 OS cells. Each conditioned medium was collected from fisetin treated U-2 OS cells (2.5, 5 and 10 μM) for 24 and 48 h to measure MMP-2 activity by using gelatin zymography (Figure 3A). Fisetin treatment at 10 μM for 48 h significantly inhibited MMP-2 activity. The results of cell migration and cell invasion assays are presented in Figure 3B and C. Figure 3B indicates that fisetin (2.5, 5 and 10 μM) significantly inhibited the migration of U-2 OS cells by about 40-70% compared to control cells. Figure 3C indicates that fisetin (10 μM) significantly inhibited the invasion of U-2 OS cells about 30% compared to control cells.

The possible signaling pathways involved in fisetin-induced inhibition of migration and invasion in U-2 OS cells in vitro.
Fisetin affected key metastasis-related proteins in U-2 OS cells. Western blotting results are presented in Figure 4. Results indicated that fisetin (5-10 μM) significantly decreased the levels of p-EGFRTyr1068, Ras, GRB2, Sos1, uPA and PKC (Figure 4A), p-ERK1/2, p-JNK, p-P38 (Figure 4B), VEGF, FAK, Rho A, PI3K, p-AktThr308, p-AktSer473, NF-ĸB, MMP-7, MMP-9 and MMP-13 (Figure 4C), however, it increased GSK3β and E-cadherin levels (Figure 4D) in U-2 OS cells after 48 h treatment. Based on these findings, fisetin suppresses cell metastasis of U-2 OS cells through multiple signaling pathways (Figure 5).
Discussion
Currently, preoperative chemotherapy, surgery, and adjuvant postoperative chemotherapy are the standard treatment strategies for OS patients. Although surgery (tumor excision) combined with adjuvant chemotherapy and radiotherapy have been used, the cure rate of OS patients remain still unsatisfied (33). The reason is due to metastatic lesions in the lungs, multi-drug resistance and the lack of better molecular biomarkers to detect OS at an early stage (34). Some of the chemotherapy drugs were developed to inhibit tumor cell proliferation, apoptosis or metastasis (35-37). Thus, blockage of the signaling pathways leading to cancer cell metastasis is one of best strategies against cancer cells (37, 38). To this end, natural products are being tested for the treatment of OS. Fisetin, which is present in fruits and vegetables, has been shown to induce apoptosis of cancer cells and to inhibit migration and invasion of latent membrane protein-1 (LMP1)-positive nasopharyngeal carcinoma (NPC) cells (24). Fisetin has been found to inhibit cervical cancer cell migration and invasion by repressing uPA via interruption of the p38 MAPK-dependent NF-ĸB signaling pathway (25). Herein, the effects of fisetin on migration and invasion in U-2 OS cells in vitro were investigated.
Treatment of U-2 OS cells with 20-40 μM fisetin for 48 h decreased their viability. Thus, in the wound healing assay, lower concentrations (2.5-10 μM) were used. Treatment of U-2 OS cells with 5-10 μM fisetin for 24 h suppressed cell mobility (Figure 2A and B) in a dose-dependent manner (Figure 2B). This is in agreement with another report showing that fisetin inhibited migration in MCF-7 cells in vitro (39). In order to further confirm this finding, transwell chambers assay was used to examine cell migration and invasion. Fisetin suppressed cell migration at 2.5-10 μM (Figure 3B) and inhibited cell invasion at 10 μM after 48 h treatment (Figure 3C) in U-2 OS cells. These results are also in agreement with another report indicating that fisetin suppressed cell migration and invasion in A549 cells (40). Our results indicate that fisetin suppresses mobility, migration and invasion of U-2 OS cells in vitro. Gelatin zymography was also used to measure the MMP-2 gelatinase activity of U-2 OS cells after treating them with 10 μM of fisetin for 48 h. Results showed that fisetin significantly reduced MMP-2 activity.
In order to understand the mechanism by which fisetin suppresses migration and invasion of U-2 OS cells, proteins associated with EMT and metastasized were analyzed (41). Western blotting analysis indicated that the levels of E-cadherin increased, while those of N-cadherin decreased. Both, E-cadherin and N-cadherin are well known to be involved in cancer cell migration and invasion (42, 43). Furthermore, fisetin was found to decrease the levels of Ras protein in U-2 OS cells (Figure 4A). It is well documented that Ras-related proteins regulate cell adhesion (44, 45). When cancer cells metastasize, decreased E-cadherin and increased N-cadherin expression are observed during EMT, that promotes adhesion to stroma and increased tumor cell motility and invasiveness (46, 47). It was reported that fisetin reduced the levels of Twist protein, an EMT regulator in LMP1-positive nasopharyngeal carcinoma cells (24). Our results showed that fisetin reduced the protein levels of the transcriptional factor Snail. Fisetin also suppressed the protein expression of MMP-7, -9 and -13 in U-2 OS cells (Figure 4C), that is in agreement with another report indicating that fisetin inhibits MMP-1, MMP-3, MMP-7, and MMP-9 in tumor cells (48). In addition, fisetin significantly reduced Rho A, FAK, p-AktThr308 and p-AktSer473 (Figure 4C) protein expression in U-2 OS cells. Rho A and activated PI3K/AKT signaling are associated with cancer cell invasion and oncogenesis (43, 49-51). Focal adhesion kinase (FAK) is involved in cell migration and invasion and the FAK-Rho A signaling pathway is also associated with cell motility (50). In the present study, fisetin was found to suppress the expression of uPA (Figure 4A), p-ERK1/2, p-JNK (Figure 4B), p-AktThr308, p-AktSer473 and NF-ĸB (Figure 4C) in U-2 OS cells. Activated AKT promoted cancer cell invasion and metastasis via stimulating secretion of MMPs (49). p-ERK1/2, and p-JNK have also been involved cell metastasis (52, 53), while NF-ĸB has been linked with tumor cell metastasis (54). Inhibition of NF-ĸB has been recognized as one of the strategies to inhibit cancer cell metastasis (55). The role of uPA has also been reported to be involved in cancer cell metastasis (1) and MMPs have been shown to be up-regulated by uPA and down-regulated by TIMPs (56). Our results suggest that fisetin suppresses migration in U-2 OS cells by inhibiting NF-ĸB and uPA.
In conclusion, fisetin significantly inhibited mobility, migration and invasion of U-2 OS cells by reducing FAK, Rho A, NF-ĸB and uPA protein levels and inhibiting MMP-2 and MMP-9 in vitro.
Acknowledgements
This work was supported by grant RD106003 from Chang Bing Show Chwan Memorial Hospital, Changhua, Taiwan, and by grant CMU107-ASIA-17 from China Medical University, Taichung, Taiwan. Experiments and data analysis were performed in part through the use of the Medical Research Core Facilities Center, Office of Research & Development at China medical University, Taichung, Taiwan.
Footnotes
Authors' Contributions
J.K. Chen, F.S. Chueh and J.G. Chung conceived and designed the experiments; K.C. Lai, H.C. Liu performed the experiments; J.K. Chen, F.S. Chueh and H.C. Liu analyzed the data; Y.P. Huang, C.C. Lin. and A.C. Huang contributed reagents/materials/analysis tools; S.F. Peng, F.S. Chueh and J.G. Chung wrote the paper.
This article is freely accessible online.
Conflicts of Interest
The Authors have no conflicts of interest to disclose.
- Received January 30, 2019.
- Revision received February 25, 2019.
- Accepted February 26, 2019.
- Copyright© 2019, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}