


Abstract
Background/Aim: Cerebral ischemia is a major cause of abnormal brain development. In a cerebral ischemia model, periventricular leukomalacia (PVL), white matter lesion and a decrease in the number of subcortical neurons were observed. The aim of this study was to investigate the effect of hypoxia on neurogenesis and cell survival. Materials and Methods: In seven-day postnatal rats, the right carotid artery was ligated. The rats were incubated either in a regular normoxic chamber (control group) or in a hypoxic chamber (PVL group, 8% 02 and 92% N2 at 37°C) for 2 h. Nestin- and NeuN-positive neurons were detected by immunohistochemistry. Results: The densities of nestin-immunoreactivity (IR) cells in the cerebral parietal cortex and subventricular zone were increased with hypoxia. NeuN-IR cells in the cerebral cortex were significantly decreased in the PVL group. Conclusion: Perinatal white matter injury induced neurogenesis, while the survival of neurons was decreased in the cerebral cortex.
Abnormal brain development often occurs in premature infants (1). It has been associated with serious clinical problems including cerebral palsy, as well as cognitive, learning and behavioural deficits (2-4). Premature infancy has recently become more prevalent (5). Periventricular leukomalacia (PVL) is the most common form of brain damage in preterm infants (6). It is characterized by cerebral white matter damage and considered to play a role in cerebral palsy (7). It is defined as coagulative necrosis and hemorrhagic necrosis and diffuse gliosis in the periventricular region with microglial activation in the cerebral white matter (8).
Periventricular leukomalacia induced by hypoxia-ischemia has been suggested as a major cause of abnormal brain development (9). Total brain volume and weight are reduced by ischemic insult (10). Maturation of myelin was impaired in one neonatal rat model with hypoxia-induced PVL (11). Hypoxia also affected neurogenesis, which is the process of generation of new neurons from progenitor stem cells (12). Neuronal cell proliferation was reduced in the subventricular zone (SVZ) (13), and cortical neuron density was decreased by hypoxia (14). Cell apoptosis was triggered after a 24-h hypoxic insult (15).
As mentioned above, periventricular leukomalacia was associated with white matter injury. Moreover, subcortical neuron density was decreased in the same model (16). Even though neurons of the cortex are affected by hypoxia, cortical lesions remain poorly understood. In this study, we investigated the effect of hypoxia on the survival of neurons and neurogenesis by immunocytochemistry.
Materials and Methods
In vivo animal PVL model. The use of certified Sprague-Dawley (SD) rats (Damul Laboratory Animals, Daejeon, Republic of Korea) was approved by the Chosun University Institutional Animal Care and Use Committee (approval no. CIACUC2016-S0002). PVL was induced by previously described methods (17). Seven day old, SD rats (n=18) were anaesthetized with Zoletil (10 mg/kg; Virbac, Nice, France) and xylazine (0.15 mg/kg; Bayer, Leverkusen, Germany). Skin and musculature of the cervical region were incised to expose the right carotid artery, which was ligated with skin sutures. The incision site was closed, and the rats rested for 10 min before incubation in a hypoxic chamber (8% O2 and 92% N2 at 37°C) for 2 h. Rats who underwent the hypoxic procedure were in the PVL group and rats exposed to normal conditions constituted the control group (n=18).
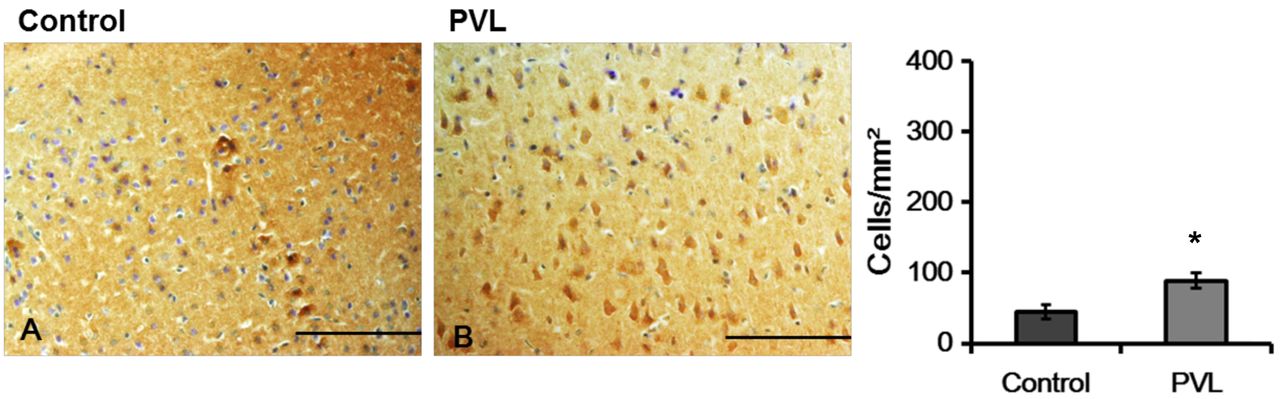
Representative photographs (A) and density (B) of nestin-immunoreactivity (IR) cells in the cerebral parietal cortex. Nestin-IR cells in PVL groups were stained strongly. The data are expressed as mean and SEM values. *p<0.05. Scale bars=100 μm.
Tissue preparation. Seven days after incubation, rats were sacrificed and their brains were harvested for analysis. The brains were kept in 4% paraformaldehyde (PFA) at 4°C for one day and were then washed in staining jars with distilled water. Dehydration and clearing were performed with gradient ethanol (70%-100%) and xylene. The cerebrums were embedded in paraffin and cut serially into 7 μM sections in the sagittal plane. The sections were mounted on gelatin-coated slides (Fisher Scientific, Pittsburgh, PA, USA).
Immunohistochemistry. The sections were deparaffinized in xylene and ethanol solutions (100%-70%) and washed in 0.1 M phosphate-buffered saline (PBS; pH 7.4). The antigen retrieval procedure was performed with 0.01 M sodium citrate buffer (pH 6.0) and heating in a microwave oven. After cooling, the slides underwent endogenous peroxidase blocking with 0.3% hydrogen peroxide for 20 min. Slides were incubated with the primary antibodies overnight at 4°C including mouse anti-hexaribonucleotide-binding protein-3 (NeuN; 1:100, Millipore) and nestin (1:200, Millipore). The next day, the slides were washed with PBS and incubated with a second antibody, which would bind to the primary antibody. The immunoreactivity was visualised with DAB Peroxidase (HRP) Substrate Kit, 3,3’-diaminobenzidine (Vector Laboratories, Burlingame, CA, USA). Thionin was utilised for counterstaining. Coverslips and PolyMount (Polysciences, Warrington, PA, USA) were applied for mounting the sections onto slides.
Statistical analysis. The nestin and NeuN-immunoreactive (IR) cells were quantified using ImagePro Plus software (Media Cybernetics). All of the experimental data were analysed using Statistical Package for Social Sciences (Information Analysis Systems, SPSS, USA). All quantified measurements were compared between the control and hypoxic groups using Student's t-tests. The statistical significance was set at p<0.05.
Results
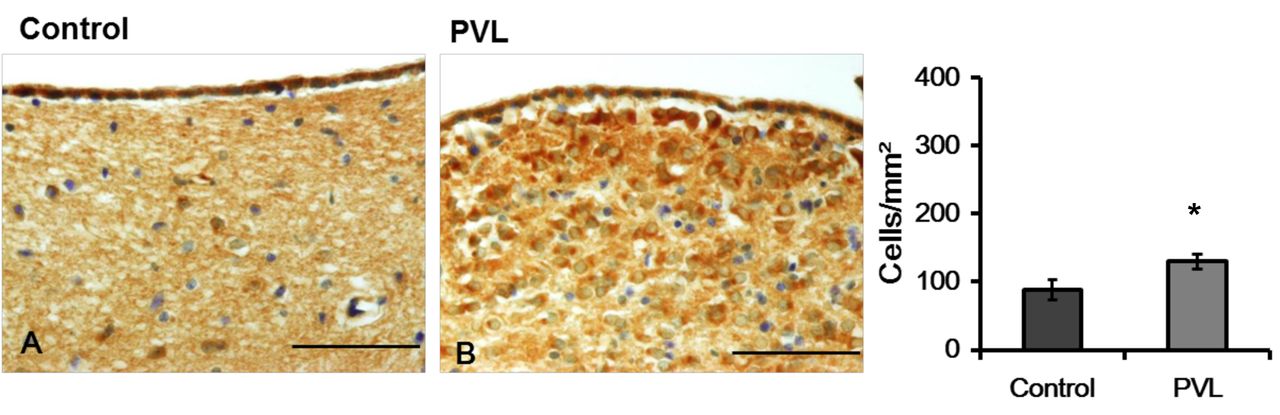
Nestin immunoreactivity. The density of nestin- IR cells in the cerebral parietal cortex was increased in the PVL group when compared to that in the control group (Figure 1). Similarly, the density of nestin-IR cells in the SVZ, defined as the neurogenic zone, was significantly greater in the PVL group than in the control group (Figure 2).
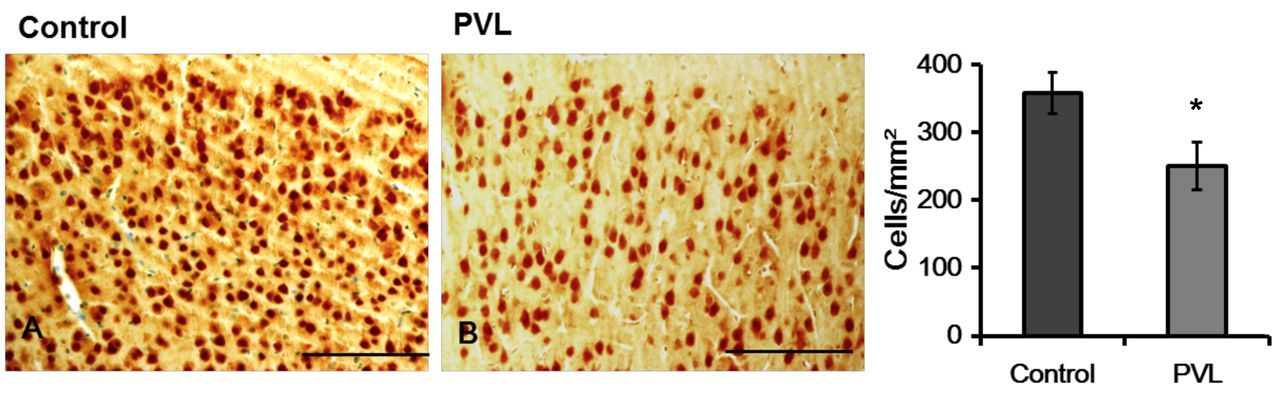
NeuN immunoreactivity. The density of NeuN-IR cells in the cerebral cortex was significantly decreased in the PVL group as compared to that in the control group (Figure 3). In the SVZ, a few NeuN-IR cells were detected. The density of NeuN-IR cells did not differ significantly (p>0.05) between the PVL group and the control group (Figure 4).
Discussion
We measured the densities of nestin- and NeuN-IR cells in the SVZ and cerebral cortex. The SVZ is capable of neurogenesis (18), which is characterized by cell proliferation, migration, maturation, and survival (19). Neuronal stem cells in the SVZ migrate to the cerebral cortex via the rostral migratory stream (20).
Nestin is a marker of neuronal progenitor cells in the developing brain and has been used to distinguish undifferentiated from differentiated cell types (21, 22). It is expressed in very immature or incompletely differentiated cells, including cells of the primitive streak and Henson node (23). In our study, the density of nestin-IR cells in the SVZ was increased with hypoxic injury. Some PVL studies have used nestin as a stem cell marker (24). Okoshi et al. showed increased nestin expression in the white matter adjacent to VPL lesions (25). SVZ-derived glial progenitor cells were considered to be self-renewing sources in neonatal rats with PVL (26). It was suggested that the early stage of neurogenesis is triggered in the SVZ in the PVL model. Interestingly, the density of nestin-IR cells in the cerebral cortex was also increased. A study by Fagel also observed newly-generated cortical cells under hypoxic conditions (27). It was suggested that cortical neurogenesis is induced in cerebral cortex as in the SVZ in PVL model.

Representative photographs (A) and density (B) of nestin-immunoreactivity (IR) cells of in the subventricular zone (SVZ). The density of Nestin-IR cells was higher in the PVL group than in the control group. The data are expressed as mean and SEM values. *p<0.05. Scale bars=100 μm.

Representative photographs (A) and density (B) of NeuN-immunoreactivity (IR) cells in the cerebral parietal cortex. The cell densities in the cerebral cortex differ significantly between control and PVL groups. The data are expressed as mean and SEM values. *p<0.05. Scale bars=100 μm.
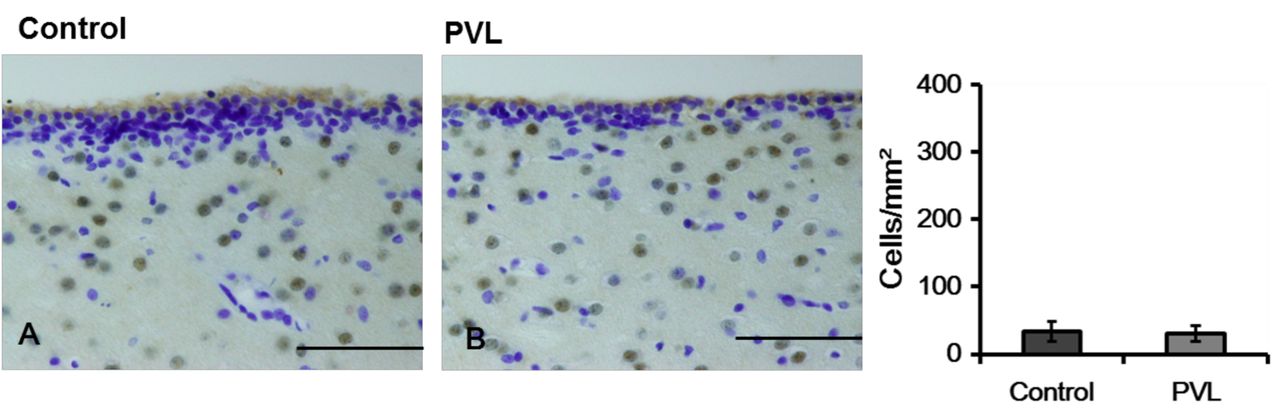
Representative photographs (A) and density (B) of immunoreactivity (IR) cells in the subventricular zone. NeuN-IR cells were seen sparsely in the SVZ in control and PVL groups. The data are expressed as mean and SEM values. *p<0.05. Scale bars=100 μm.
NeuN is a neuronal-specific nuclear protein (28). NeuN antibody was used to estimate the total numbers of neuronal and non-neuronal cells (29). The density of NeuN-IR cells in the cerebral cortex was significantly decreased in the PVL group. Similarly, premature infants with PVL have shown to present focal neuron loss in the frontal lobe (30). Later in childhood, some premature infants are reported to have persistent reduction in cortical volume (31, 32).
Conclusion
In summary, our results indicated that PVL results in decreased number neurons in the cerebral cortex despite evidence of increased neurogenesis. Given that progenitor cells in the SVZ migrate into the cortex, our results suggest that neurogenesis cannot compensate for the VPL-induced cerebral neuronal loss.
Acknowledgements
This study was supported by the ‘Chosun University Research Aid Scholarship Program’ for undergraduate students, 2017.
Footnotes
Authors' Contributions
Yonghyun Jun designed the study and participated in the surgical procedures. Hanbit Cho, Gyeong Hyeon Jo analyzed the obtained data. Yoonyoung Chung performed the immunohistochemistry. All Authors read and approved the final manuscript.
This article is freely accessible online.
Conflicts of Interest
The Authors declare that they have no competing interests.
- Received December 27, 2018.
- Revision received January 17, 2019.
- Accepted January 21, 2019.
- Copyright© 2019, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Dexmedetomidine Stereotaxic Injection Alleviates Neuronal Loss Following Bilateral Common Carotid Artery Occlusion via Up-Regulation of BDNF Expression
- Differential Expression of Pax6 Following Bilateral Common Carotid Artery Occlusion
- IGF-1 Protects Neurons in the Cortex and Subventricular Zone in a Periventricular Leucomalacia Model
- Immunoreactivity of MAPK Signaling in a Rat Model of Intrauterine Growth Retardation Induced by Uterine Artery Ligation
- Differential Expression of Vascular Endothelial Growth Factor in the Cortex and Hippocampus upon Cerebral Hypoperfusion