


Abstract
Induction of corpus luteum regression and subsequent estrus using prostaglandin F2α (PGF2α) in microminipigs was investigated. Microminipigs with normal estrous cycle were treated with PGF2α as 0.75 mg (0.75 PG group, n=3) or 1.5 mg (1.5 PG group, n=4) dinoprost injected into the vulva at 24-h intervals at 10 days after the onset of estrus (D0), D1 and D2. Three microminipigs were not treated (control group). The estrous interval in the 1.5 PG group was significantly shortened compared to the control and 0.75 PG groups. Plasma progesterone levels started to decline and reached the base line in the 1.5 PG group significantly faster than in the control group. In conclusion, we demonstrate that multiple PGF2α treatments can induce corpus luteum regression and estrous synchronization in female microminipigs.
In 2011, the smallest miniature pig in the world, the microminipig was developed (1) and this strain has similar general biochemical and hematological (2-5) profiles to humans. The utility of microminipigs as experimental animals has been demonstrated in research on atherosclerosis (6-9), cardiovascular pharmacology (10-12), sleep conditions (13), coagulatory activity (2), toxicology (14, 15), genomic sequencing (16) and developmental engineering (17). Because microminipigs are accordingly expected to be an important animal model for future human health research, effective and stable production of microminipigs is needed. Unlike popular mouse models, the microminipig represents a potentially more appropriate experimental model, since their lipoprotein metabolism (9, 18), as well as their anatomy, physiology and feeding and sleep habits (13), are much more similar to those of humans.
The development of estrous synchronization in female animals is needed to improve the efficiency of animal production using novel reproductive techniques such as artificial insemination and embryo transfer. In the swine industry, the benefits of new reproductive technologies are well known in transferring germ cells (sperm and embryo) with a low risk of disease transmission and low costs compared with live animal transfer (19). In particular, embryo transfer is essential for the application of other reproductive biotechnologies, including transgenesis and cloning, not only in domestic pigs (20) but also miniature pigs (17, 21).
It is well known that prostaglandin F2α (PGF2α) is an effective regulator of the corpus luteum (CL) function in domestic animals (22). In domestic pigs, a single PGF2α treatment cannot induce luteolysis prior to the first 12 days of the estrous cycle, thus multiple PGF2α treatment is needed to induce regression of functional CL before 12 days of the estrous cycle (23-25). This estrous synchronization using multiple PGF2α treatment is an effective method for the preparation of recipients for artificial insemination (26) and embryo transfer (27, 28) in domestic pigs. However, multiple treatments for luteolysis induction and estrous exhibition in cyclic domestic pigs are an important issue in animal welfare. It was recently demonstrated that the effective dose for inducing regression of CL and subsequent farrowing in pregnant domestic pigs through intra-vulval injection of PGF2α is 25-50% of that injected intramuscularly (29, 30). PGF2α is secreted in pulses from the endometrium in many animal species, including cattle, pig and horses (22). The passage of PGF2α from the uterus to the CL involves close apposition of efferent uterine vessels (venous and lymphatic) with ovarian arterial vessels. Because prostaglandins are rapidly metabolized mainly in the lungs but also in other organs such as the liver and kidney, intra-vulval PGF2α injection is more effective in increasing the local concentration of PGF2α in the female reproductive tract without first-pass effects in the lung than intramuscular treatment. These reports (29, 30) suggest it may be possible to control luteal function through reduction of PGF2α doses and treatment times not only in domestic pigs but also experimental minipigs, such as microminipigs, as both have similar hormonal profiles during normal estrous cycles (31). In the present study, we examined the effect of multiple PGF2α treatment into the vulva on the luteal lifespan and subsequent estrous exhibition associated with hormonal changes in female microminipigs.
Effect of prostaglandin F2α (PG) treatment on regression of corpora lutea and length of estrus.
Materials and Methods
Animals and estrous detection. Eleven mature microminipig gilts were purchased from Fuji Micra Inc. (Shizuoka, Japan) and maintained at Kagoshima University in a dedicated, climate-controlled room with a temperature of 18-25°C and humidity of 30-65%, with a 12-h light/dark cycle at the breeder's facility. They were kept in individual cages with water available ad libitum and were fed a commercial diet for domestic pigs (Kodakara 73; Marubeni Nissin Feed Co., Tokyo, Japan) in quantities of 1.8-2.2% of body weight once daily. Estrous detection was performed twice daily using a mature male microminipig. At least one estrous cycle of normal length (18-22 days) (31) was observed in each microminipig before the start of each experiment. The ages and weights (mean±SEM) of the study animals at treatment were 15.8±0.8 months and 14.5±0.3 kg, respectively. All protocols were approved by the Kagoshima University Ethics Committee of Animal Care and Experimentation (VM13037) and the research was performed according to the Institutional Guidelines for Animal Experiments and in compliance with the Japanese Law Concerning the Protection and Control of Animals (Law No. 105 and Notification No. 6).
Study design. We investigated the effect of multiple intra-vulval administrations of PGF2α on estrus exhibition and ovarian steroid profiles in microminipigs. They were given PGF2α (5 mg dinoprost/ml, Panaceran Hi; Meiji Seika Falma Co., Ltd., Kyobashi, Tokyo, Japan) as subcutaneous injection of 0.75 mg (0.75 PG group, n=3) or 1.5 mg (1.5 PG group, n=4) dinoprostin into the vulva with a 22-G needle at 24-h interval at 10 days after the onset of estrus (D0), D1 and D2. The effective dose of PGF2α for regression of CL was derived from previous research (32). Three animals were no treated with PGF2α (control group). Each animal was fitted with a heparin-coated polyurethane catheter (CBAS-C70; Solomon Scientific, Plymouth Meeting, PA, USA) in the jugular vein at least 7 days from the onset of estrus. Blood samples were collected daily beginning at D −2 continuing until 7 days after the onset of the subsequent estrus. Additionally, blood samples were collected at 12-h intervals from D0 until the end of the subsequent estrus. Estrous detection in all female microminipigs was carried out every 12 h starting from D3 to the end of the subsequent estrus. Plasma was recovered after centrifugation of the blood samples and stored at −30°C until analysis.
Hormone assay. Time-resolved fluoroimmunoassay was used to measure plasma progesterone and estradiol-17β with DELFIA estradiol and progesterone kits (PerkinElmer Japan, Yokohama, Japan) as previously reported (33). The intra-assay and inter-assay CVs were 8.1% and 8.1% for estradiol-17β and 10.0% and 9.1% for progesterone, respectively.
Statistical analyses. Statistical analyses were performed using the general linear model procedure in SAS version 9.4 (SAS Institute Inc. Cary, NC., USA). Data pertaining to the hormonal profiles were subjected to repeated measures analysis of variance (ANOVA) (34). When a significant effect was detected with ANOVA, the significance of the difference between the means was determined using Tukey's test. The differences in intervals from D0 to the day of progesterone concentration >1 ng/ml among groups were also tested for significance using the Tukey's test. Because we were unable to detect estrous exhibition or increased estradiol-17β level in one microminipig in the 0.75 PG group after treatment, the differences in the estrous duration, the estrous length and the interval from D0 to the day of peak estradiol-17β concentrations were compared between the control and 1.5 PG groups and tested for significance using the Student's t-test. Differences with a p-value of less than 0.05 were considered statistically significant.
Results
The interval from D0 to the day of luteolysis in the 1.5 PG group was shorter (p<0.05) than that in the control and 0.75 PG groups (Table I). Although there was no difference in the estrous duration after treatment between the control and 1.5 PG groups, the estrous length after treatment in the 1.5 PG group was significantly shorter compared with the control group (p<0.05).
Ovarian steroid hormone profiles in the microminipigs after treatment are shown in Figure 1. Peripheral progesterone concentrations started to decrease (p<0.05) on day 6 in the control group and day 2.5 in the 1.5 PG group compared with concentrations at D0. Progesterone concentrations between D2 and D6.5 in the 1.5 PG group were significantly lower than those in the control group during the same period (p<0.05). There was a significant difference in progesterone levels on days 10, 10.5, 12, 15, 16 and 17 between the control and 1.5 PG groups (p<0.05). Estradiol-17β concentrations were significantly greater (p<0.05) on D9.5 in the control group and D5.5 in the 1.5 PG group than between D−2 and 0, respectively. The maximum estradiol-17β levels reached 30.7±1.0 pg/ml (day 11.3±0.9) in the control group and 38.8±2.8 pg/ml (day 7.3±0.1) in the 1.5 PG group. Concentrations of estradiol-17β from day 3 to 7.5 and from day 10 to 10.5 in the 1.5 PG group were higher and lower (p<0.05), respectively, than those in the control group.
Discussion
We clearly showed that triple administration of 1.5 mg PGF2α into the vulva during the functional luteal phase can shorten the inter-estrous interval in microminipigs. When female pigs were given 12.5 mg PGF2α 12 times (25) or 15 mg PGF2α six times intramuscularly (35, 36) during the luteal phase of the estrous cycle, inter-estrous intervals in the treatment groups (13.3-15.7 days) were significantly shortened compared to the control groups (19.8-21.2 days). Our results are in accordance with these inter-estrous intervals (25, 35, 36), thereby demonstrating that the estrous length in cyclic microminipigs can indeed be controlled with the fewer injections than that needed in domestic pigs.
Intramuscular treatment of a total of 90 mg dinoprost (6×15 mg dinoprost treatments) between 8 and 10 days from the last day of the estrous cycle induced luteolysis within 3 days after the first injection in all cyclic domestic pigs (average body weight of 176 kg) (36). In contrast the efficiency of shortening the estrous length in domestic pigs (average body weight of 167-199 kg) with treatment of 60 or 30 mg total dinoprost (6×5 or 10 mg dinoprost treatments) for the same period was 70% (36). From previous reports in domestic pigs, at least 75 mg dinoprost (approximately 0.6 mg/kg/experimental period) during the experimental period may be needed to induce luteolysis during the early luteal phase of the estrous cycle with high efficiency. Plasma progesterone concentrations, a general index of CL condition (37), in microminipigs treated with 1.5 mg intra-vulva dinoprost injection on D0 to D2 reached baseline 3.7 days from the first treatment day, which is similar to previous reports (25, 35, 36). In our research, total treatment dose (4.5 mg dinoprost) per experimental period (approximately 0.3 mg/kg/experimental period) in microminipigs treated with intra-vulval administration of PGF2α induced the decline of progesterone levels in the 1.5 PG group but not in the 0.75 PG group. In domestic pigs, it has been demonstrated that the effective dose for regressing CL function using intra-vulval PGF2α injection can be reduced to 25-50% of the effective intramuscular dose (29, 30). Our results and previous reports suggest that reaction of CL following exogenous PGF2α treatment is similar in microminipigs and domestic pigs.
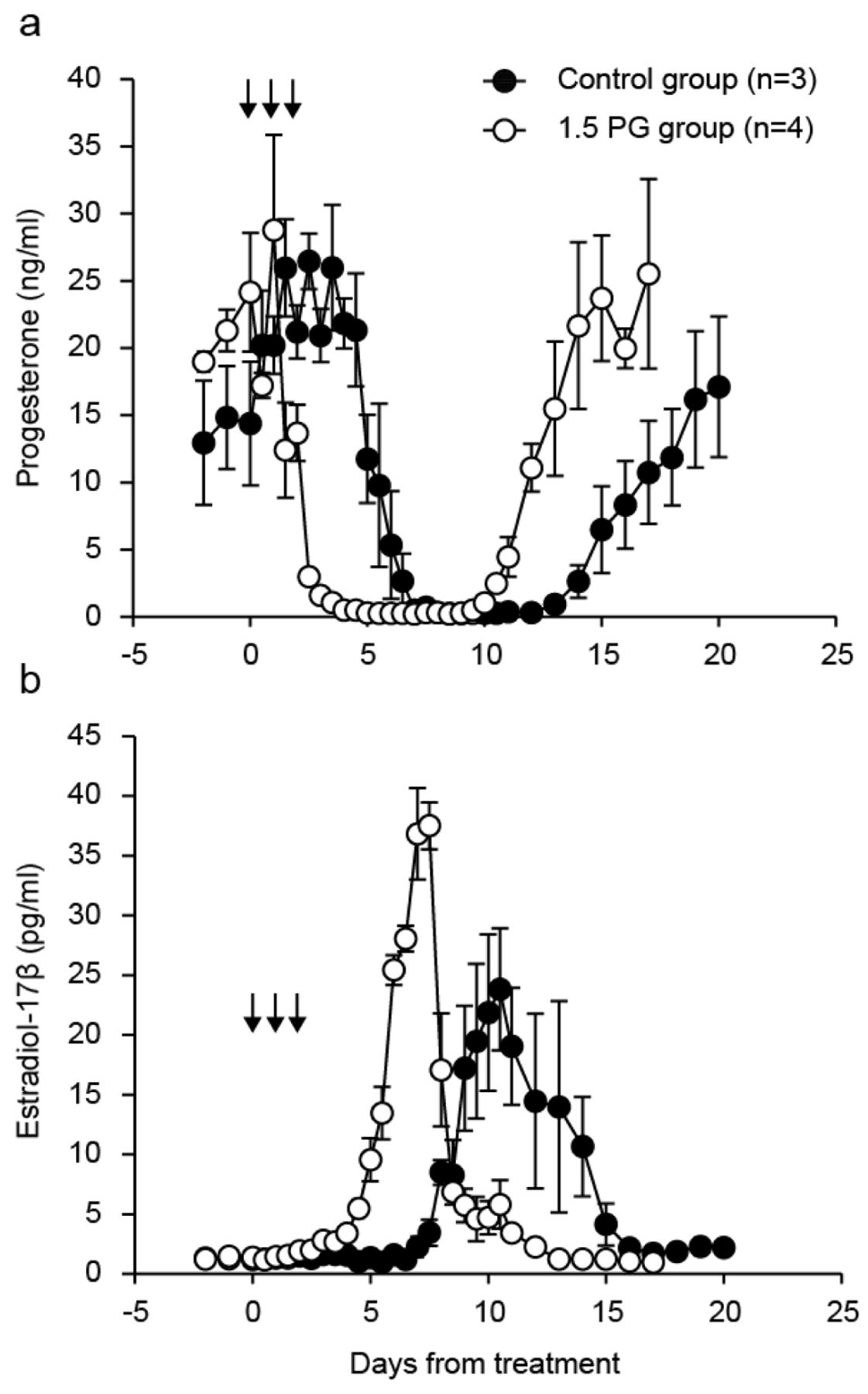
Plasma concentrations of progesterone (a) and estradiol-17β (b) in the control group (n=3) and the group treated with 1.5 mg prostaglandin F2α (PG) (n=4) groups. Each arrow indicates a treatment day in the 1.5 PG group. Values are means±SEM.
Progesterone levels started to decrease on the last day of treatment in the 1.5 PG group. This finding is the same as those in previous reports using domestic pigs with multiple PGF2α injection (35, 36). Even though progesterone concentrations in domestic pigs gradually decreased following multiple PGF2α treatment, progesterone levels increased again after treatment when the CL maintained their function within 24 h after the final PGF2α treatment (36). Thus, we conclude that it is difficult to induce luteolysis with fewer than three 1.5 mg dinoprost injections in microminipigs.
Because estradiol-17β is secreted from large mature follicles in domestic pigs (37) and microminipigs (31), the profile of estradiol-17β in plasma reflect the development of mature follicles in the ovaries. The peak of estradiol-17β concentrations in the 1.5 PG group was defined at D7 or D7.5 in contrast to the control group (which varied from D10 to D13) in this study. This means that the protocol of PGF2α treatment in microminipigs affects not only CL regression but also synchronization of follicular development and subsequent estrous exhibition.
In conclusion, we indicate a new protocol for induction of CL regression and synchronization of estrous in female microminipigs using multiple PGF2α treatments. Development of assisted reproductive technology such as synchronization of estrus and ovulation serve to improve reproductive technologies and productive efficiency in animals. The new estrous synchronization protocols from our research will help to improve the efficiency of microminipig production.
Acknowledgements
This work was partly supported by JSPS KAKENHI Grant Numbers 26870455 and 16H05176
Footnotes
This article is freely accessible online.
Conflicts of Interest
The Authors report no conflicts of interest exist in regard to this study.
- Received August 19, 2017.
- Revision received September 17, 2017.
- Accepted September 19, 2017.
- Copyright© 2017, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved




{kind=link}