


Abstract
Background/Aim: To clarify the mechanisms responsible for the anti-inflammatory/proinflammatory activities of eugenol-related compounds, we investigated the cytotoxicity and up-regulatory/down-refgulatory effects of the biphenols curcumin, bis-eugenol, magnolol and honokiol, and the monophenols eugenol and isoeugenol, on major regulators of cyclooxygenase-2 (Cox-2), nitric oxide synthase 2 (Nos2) and heme oxygenase-1 (HO-1) mRNA in RAW264.7 cells. Materials and Methods: mRNA expression was investigated using real-time reverse transcriptase-polymerase chain reaction (RT-PCR), and the theoretical parameters were calculated using the DFT/B3LYP/6-31* method. Also, the antioxidant activity of eugenol-related compounds in combination with 2-mercapto-1-methylimidazole (MMI, as a model for glutathione (GSH)) was investigated using the induction period method for polymerization of methyl methacrylate initiated by benzoyl peroxide (BPO). Results: The cytotoxicity of eugenol-related compounds showed a linear relationship with their softness (σ) and electrophilicity (ω). At a concentration of 50 μM, biphenols except for bis-eugenol elicited the expression of mRNA for both Cox-2 and Nos2, but monophenols did not. In contrast, bis-eugenol elicited Cox-2 gene expression, but down-regulated Nos2 gene expression. bis-Eugenol alone induced the expression of HO-1 mRNA, and when combined with MMI it showed a potent antagonistic effect on BPO-induced antioxidant activity. The ability of methoxyphenols to inhibit LPS-stimulated Cox-2 gene expression declined in the order curcumin >> isoeugenol > bis-eugenol >> eugenol, and the rank of ability was related to their ω value. Conclusion: Most eugenol-related compounds had proinflammatory activity at high concentrations. However, they had also anti-inflammatory activity at lower concentrations. Eugenol-related compounds may exert antioxidant and anti-inflammatory activity in LPS-stimulated RAW264.7 cells possibly by inhibiting the activation of nuclear factor-kappa B (Nf-ĸB), whereas bis-eugenol requires induction of HO-1 expression. bis-Eugenol as well as curcumin, may have anti-inflammatory and anticancer therapeutic applications.
Eugenol-related compounds are known to have antioxidant, anti-inflammatory, antiviral, antifungal, antibacterial, anticancer, antidiabetic and neuroprotective properties (1-3). We have previously shown that the biphenols curcumin, bis-eugenol, magnolol and honokiol exert antioxidant effects and also inhibit the up-regulation of cyclooxygenase-2 (Cox-2) mRNA expression elicited by lipopolysaccharide (LPS) and Porphyromonas gingivalis fimbriae (PGF) (4, 5). We have also shown that the prooxidant activity of eugenol, isoeugenol and curcumin is inhibited by visible light irradiation and treatment with horseradish peroxidase, suggesting that these compounds elicit production of intracellular reactive oxygen species (ROS) under prooxidative conditions (6-8). Interestingly, low and physiological levels of ROS are required for normal physiological activity in cells, whereas high levels of ROS promote cytotoxicity, apoptosis, and inflammatory activity. A high level of ROS modulates a number of cell signaling pathways, and regulates the expression of multiple genes such as Cox-2 and nitric oxide synthase (Nos) 2 in vitro and in vivo (9). The anti-inflammatory and anti-proliferative effects of curcumin and its related compounds are known to be induced through a ROS-independent mechanism (10). In general, inflammatory activity is accompanied by overexpression of inducible nitric oxide synthase (iNOS), leading to production of nitric oxide, which enhances the catalytic activity COX-2 via formation of the peroxinitrite anion (11). COX-2 is a downstream target of NOS2. Also, heme oxygenase-1(HO-1), the inducible isoform of HO, catalyzes the degradation of heme into biliverdin, iron, and carbon monoxide (CO), and inhibits immune responses and inflammation in vivo. Biliverdin and bilirubin are potent antioxidants that attenuate oxidative stress. HO-1 has anti-inflammatory, antioxidant, and antiproliferative effects (12, 13), and is induced by oxidative stress.
In the present study, we investigated the cytotoxicity of ten eugenol-related compounds – eugenol, isoeugenol, bis-eugenol, curcumin, honokiol, magnolol, 4-cresol, 4-cresol dimer, 4-methoxyphenol and 4-methoxyphenol dimer – for RAW264.7 cells using a Cell Counting Kit-8 (CCK-8) in relation to theoretical parameters such as softness (σ), electrophilicity (ω) and the lowest unoccupied molecular orbital energy (ELUMO). Data on cytotoxicity and theoretical parameters had been obtained in our previous studies (5,14). The parameters of some compounds were recalculated using the DFT/B3LYP/6-31G* method. We also investigated whether eugenol-related compounds induced the expression of mRNAs for Nos2, Cox-2 and HO-1 in RAW264.7 cells, and the inhibitory effects of these compounds on Cox2-mRNA expression stimulated by LPS. Furthermore, to clarify the antioxidant activity of radical-mediated eugenol-related compounds in the presence of glutathione (GSH), a thiol, we investigated the antioxidant effect of eugenol-related compounds with or without 2-mercapto-1-methylimidazole (MMI, a thiol). The antioxidant activity was evaluated by the induction period method using differential scanning calorimetry (DSC) monitoring of the polymerization of methyl methacrylate (MMA), initiated by thermal decomposition of 0.5 mol% and 1 mol% benzoyl peroxide (BPO, a PhCOO* radical) under nearly anaerobic conditions (15). MMI was used as a model of GSH in our system because GSH is not soluble in MMA. The synergistic/antagonistic interaction between eugenol-related compounds and MMI was studied using the induction period method, and the results were evaluated on the basis of bond dissociation enthalpy (BDE) difference, and ΔBDE between the BDE of eugenol-related compounds (O-H BDE) and that of cysteine (S-H BDE).
Materials and Methods
Materials. Eugenol (4-allyl-2-methoxyphenol), isoeugenol (4-propenyl-2-methoxyphenol), MMA, BPO and MMI were purchased from Tokyo Kasei Co. (Tokyo, Japan). Curcumin, 1,7-bis (4-hydroxy-3-methoxyphenol)-1,6-heptadiene-3,5-dione, was purchased from Wako Pure Chemical Industries, Ltd. (Osaka, Japan). Magnolol (4-allyl-2-(5-allyl-2-hydroxy-phenyl) phenol) and honokiol (2-(4-hydroxy-3-prop-2-enyl-phenyl)-4-prop-2-enyl-phenol) were obtained from Kishida Chemical Industries, Ltd., Osaka, Japan. bis-Eugenol (3,3’-dimethoxy-5,5’-di-2-propenyl-1,1’-biphenyl-2,2’-diol) was synthesized from eugenol monomers by the CuCl (OH)-catalyzed ortho coupling reaction described previously (16). The chemical structures of these compounds are shown in Figure 1. Solutions of these compounds were prepared by dissolving each of them in dimethyl sulfoxide, followed by dilution to the indicated concentrations using serum-free RPMI-1640 (Invitrogen Co., Carlsbad, CA, USA) as test samples. Fetal bovine serum (FBS) was obtained from HyClone (Logan, UT, USA). Escherichia coli O111 B4-derived LPS was obtained from List Biological Laboratories, Inc. (Campbell, CA, USA).
Cell culture. The murine macrophage-like cell line RAW264.7, obtained from Dainippon Sumitomo Pharma Biomedical Co. Ltd. (Osaka, Japan), was used. The cells were cultured to a subconfluent state in RPMI-1640 medium supplemented with 10% FBS at 37°C and 5% CO2 in air, washed, and then incubated overnight in serum-free RPMI-1640. They were then washed again and treated with the test samples.
Preparation of total RNA and real-time polymerase chain reaction (PCR). The preparation of total RNA and the procedure for real-time PCR have been described previously (5). In brief, RAW264.7 cells in NUNC 96-flat-well-type microculture plates (105 cells per well) were treated with the test samples. Total RNA was isolated using an RNeasy Plus Micro Kit (Qiagen Japan Co. Ltd., Tokyo, Japan) in accordance with the instruction manual. cDNA was synthesized from total RNA (2 μg) of each sample by random priming using a High Capacity RNA-to-cDNA Kit (Life Technologies Japan, Tokyo, Japan). Reaction mixtures without the reverse transcriptase were used as a negative control. An aliquot of each cDNA synthesis reaction mixture was diluted and used for real-time PCR quantification. An equal-volume aliquot of each cDNA was mixed, serially diluted, and used as a standard. TaqMan probes/primers for Cox-2, Nos2, HO-1 and 18s rRNA and the PCR enzyme mix for real-time PCR were purchased from Life Technologies Japan. Real-time PCR quantification was performed in triplicate using the GeneAmp Sequence Detection System 5700 software (Life Technologies Japan, Tokyo, Japan) in accordance with the instruction manual. The relative amount of target was calculated from standard curves generated in each PCR, and quantitative data with a coefficient of variance (CV) of less than 10% were used for further analyses. Each calculated amount of mRNA was standardized by reference to that for 18s rRNA. Data are expressed as means of three independent experiments. Statistical analyses were performed using Student's t-test and one-way ANOVA.
Radical-scavenging activity. The induction time (IT) and initial rate of polymerization (propagation rate, Rp) in the presence or absence of an antioxidant were determined by the induction period method reported previously (15, 17). In brief, the experimental resin consisted of MMA and BPO with or without additives. BPO was added at 0.5 mol% and 1 mol%, and the additives were used at 0.01 mol%. The mixture of eugenol-related compounds and MMI was prepared at a 1:1 molar ratio. Approximately 10 μl of the experimental resin (MMA: 9.15-9.30 mg) was.loaded into an aluminum sample container and sealed by applying pressure. The container was placed in a DSC (model DSC 3100; Mac Science Co., Tokyo, Japan) kept at 70°C, and the thermal changes induced by polymerization were recorded for the appropriate periods. The heat due to the polymerization of MMA was 13.0 kcal mol−1. The conversion of all samples (%) was calculated from the DSC thermograms using the integrated heat evoked by polymerization of MMA. The conversion value was 93.5-96.7%. The IT and propagation rate (Rpinh) for eugenol-related compounds with or without MMI was calculated using time-exotherm and time-conversion curves. The procedure was similar to that reported previously (15, 17).

The chemical structures of eugenol, isoeugenol, curcumin, bis-eugenol, magnolol and honokiol.
Measurement of the stoichiometric factor (n)
The n value in Eq. (1) can be calculated from the induction time in the presence of inhibitors (antioxidants) as:
 (eq.1)
where IT is the induction time in the presence of a phenolic inhibitor (IH). The number of moles of BPO radicals trapped by the antioxidant was calculated with respect to 1 mole of the inhibitor moiety. The initiation rate (Ri) for BPO at 70°C was 2.28×10−6 mol l−1s−1, calculated on the basis of n=2.0 for 2,6-di-tert-butyl-4-methoxyphenol (17).
(eq.1)
where IT is the induction time in the presence of a phenolic inhibitor (IH). The number of moles of BPO radicals trapped by the antioxidant was calculated with respect to 1 mole of the inhibitor moiety. The initiation rate (Ri) for BPO at 70°C was 2.28×10−6 mol l−1s−1, calculated on the basis of n=2.0 for 2,6-di-tert-butyl-4-methoxyphenol (17).
The inhibition rate constant (kinh) is given by
 (eq.2)
where kp is the propagation rate constant, and MMA, IT and Rpinh are defined above.
(eq.2)
where kp is the propagation rate constant, and MMA, IT and Rpinh are defined above.
Calculation
The O-H BDE values for eugenol-related compounds were taken from the data reported previously (3). Those for magnolol and honokiol were recalculated as follows. Firstly, the lowest and second lowest energy conformers of both phenol derivatives and their phenoxyl radical species were identified as candidates for geometry optimization using a conformer search procedure employing Merck Molecular Mechanics force fields (MMFF) calculation. The tentative conformers were them optimized for geometry by the restricted or unrestricted hybrid Hartree-Fock DFT calculation for the phenols and the phenoxyl radicals in vacuo using B3LYP functional on the 6-31G* basis set level to afford the respective energetic minimized structures (18):
 where Hr is the enthalpy of the phenoxyl radical generated by H-abstraction, Hh is the enthalpy of the hydrogen radical, and Hp is the enthalpy of the parent phenol. The energy values of both the highest occupied molecular orbital (HOMO) and the lowest unoccupied molecular orbital (LUMO) energy of eugenol-related compounds were taken from our previous reports (5). The O-H BDE values for curcumin, magnolol and honokiol were recalculated. Briefly, all of the molecular modeling and calculations were performed using the Spartan 10 for Windows software package (Wavefunction Inc., Irvine, CA, USA). Chemical hardness (η) softness (σ), electronegativity (χ) and electrophilicity (ω) and Koopman's theorem ionization potential (IP, | EHOMO |) were calculated using equations 2, 3, 4 and 5, respectively:
where Hr is the enthalpy of the phenoxyl radical generated by H-abstraction, Hh is the enthalpy of the hydrogen radical, and Hp is the enthalpy of the parent phenol. The energy values of both the highest occupied molecular orbital (HOMO) and the lowest unoccupied molecular orbital (LUMO) energy of eugenol-related compounds were taken from our previous reports (5). The O-H BDE values for curcumin, magnolol and honokiol were recalculated. Briefly, all of the molecular modeling and calculations were performed using the Spartan 10 for Windows software package (Wavefunction Inc., Irvine, CA, USA). Chemical hardness (η) softness (σ), electronegativity (χ) and electrophilicity (ω) and Koopman's theorem ionization potential (IP, | EHOMO |) were calculated using equations 2, 3, 4 and 5, respectively:
 (eq.3)
(eq.3)
 (eq.4)
(eq.4)
 (eq.5)
(eq.5)
 (eq.6)
(eq.6)
Results
Relationships between cytotoxicity and σ, ω or ELUMO. Ten eugenol-related compounds – 4-cresol, 4-methoxyphenol and eugenol and their ortho dimers, curcumin, isoeugenol, magnolol and honokiol – were selected. The theoretical parameters and 50% cytotoxic concentrations (LC50) are shown in Table I, respectively.
From the data in Table I, eqns. 7, 8 and 9 were formulated:
 (eq.7)
(eq.7)
 (eq.8)
(eq.8)
 (eq.9)
The logarithm of 1/LC50 for eugenol-related compounds was linearly related to their softness (σ), ELUMO and electrophilicity (ω) values.
(eq.9)
The logarithm of 1/LC50 for eugenol-related compounds was linearly related to their softness (σ), ELUMO and electrophilicity (ω) values.
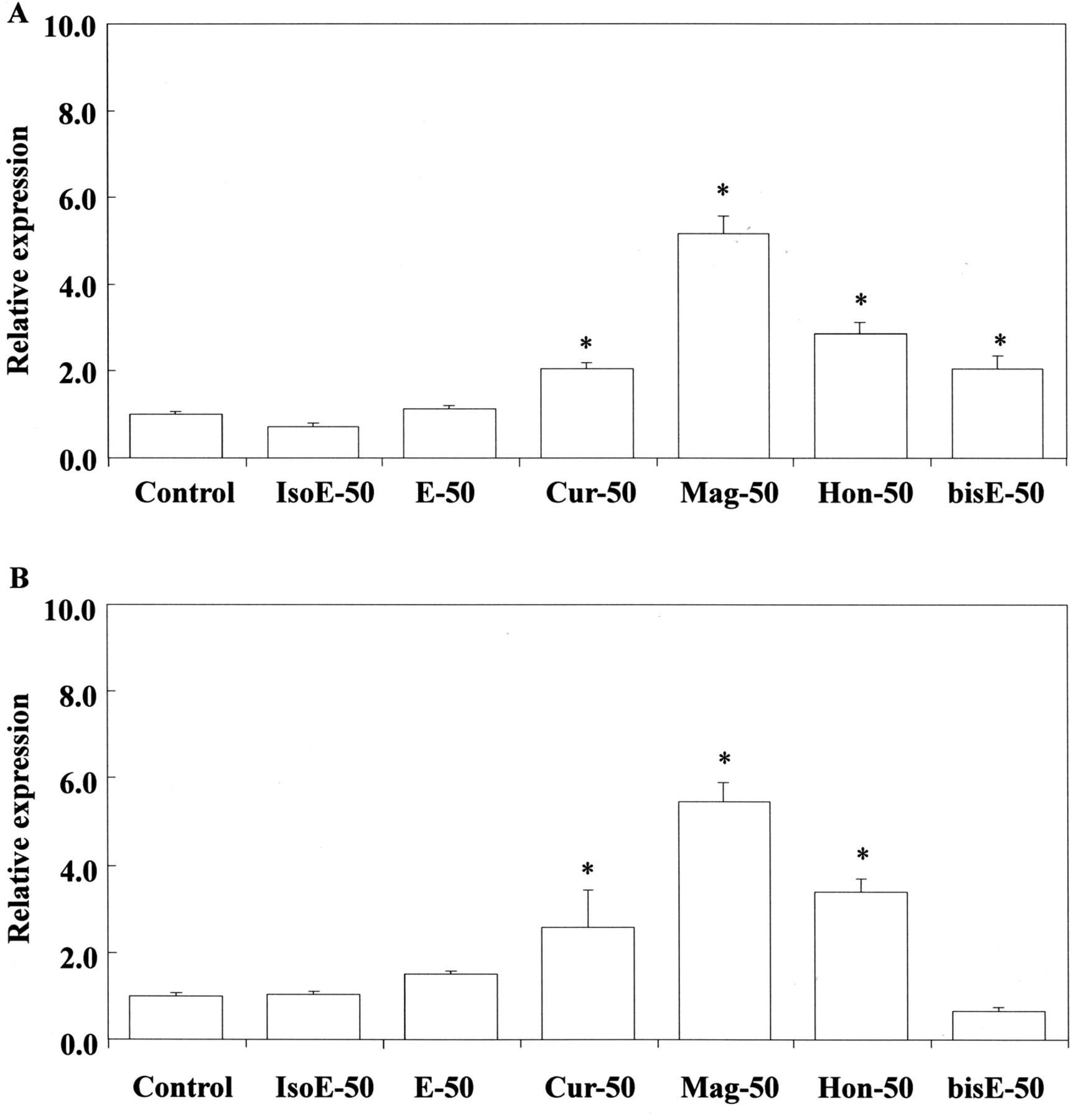
Cox-2, Nos2 and HO-1 mRNA expression. First, we examined the expression of mRNAs for Cox-2 and Nos2 in RAW264.7 cells after treatment with eugenol-related compounds at 0.1, 50 and 1000 μM. The results are shown in Figure 2A and B. Curcumin, eugenol and isoeugenol up-regulated the expression of Cox-2 mRNA at the high concenration of 1000 μM. Isoeugenol at 1000 μM elicited much higher Cox-2 mRNA expression than did eugenol or curcumin (p<0.05) (Figure 2A). There was no significant difference between the effects of curcumin and eugenol. We also examined the expression Cox-2 mRNA elicited by bis-eugenol, magnolol and honokiol at 1000 μM (Figure 2B). bis-Eugenol and magnolol, but not honokiol, a stereo isomer of magnolol, up-regulated the expression of Cox-2 mRNA, but to a lesser degree than the monophenols and curcumin. Note that the values were expressed relative to the normal control as 1. Next, we investigated whether eugenol-related compounds up-regulated the expression of Nos2 mRNA at 1000 μM. Eugenol, isoeugenol and curcumin up-regulated the expression of Nos2 mRNA (Figure 3); bis-eugenol and magnolol also did so, but the effect was weaker than that of the monophenols or curcumin. Honokiol had no effect on Nos2 mRNA expression. These effects were similar to those on Cox-2 mRNA expression (Figure 3). We then investigated the effects of eugenol-related compounds on Cox-2 and Nos2 mRNA expression at the low concentration of 50 μM. Magnolol, honokiol and curcumin – and particularily magnolol – up-regulated the expression of Nos2, whereas bis-eugenol down-regulated it (Figure 4). Monophenols had no effect on Nos2 mRNA expression. The effect of magnolol on Cox-2 and Nos2 mRNA expression was greater than that observed for the positive control treated with LPS.
Theoretical parameters for eugenol-related compounds.
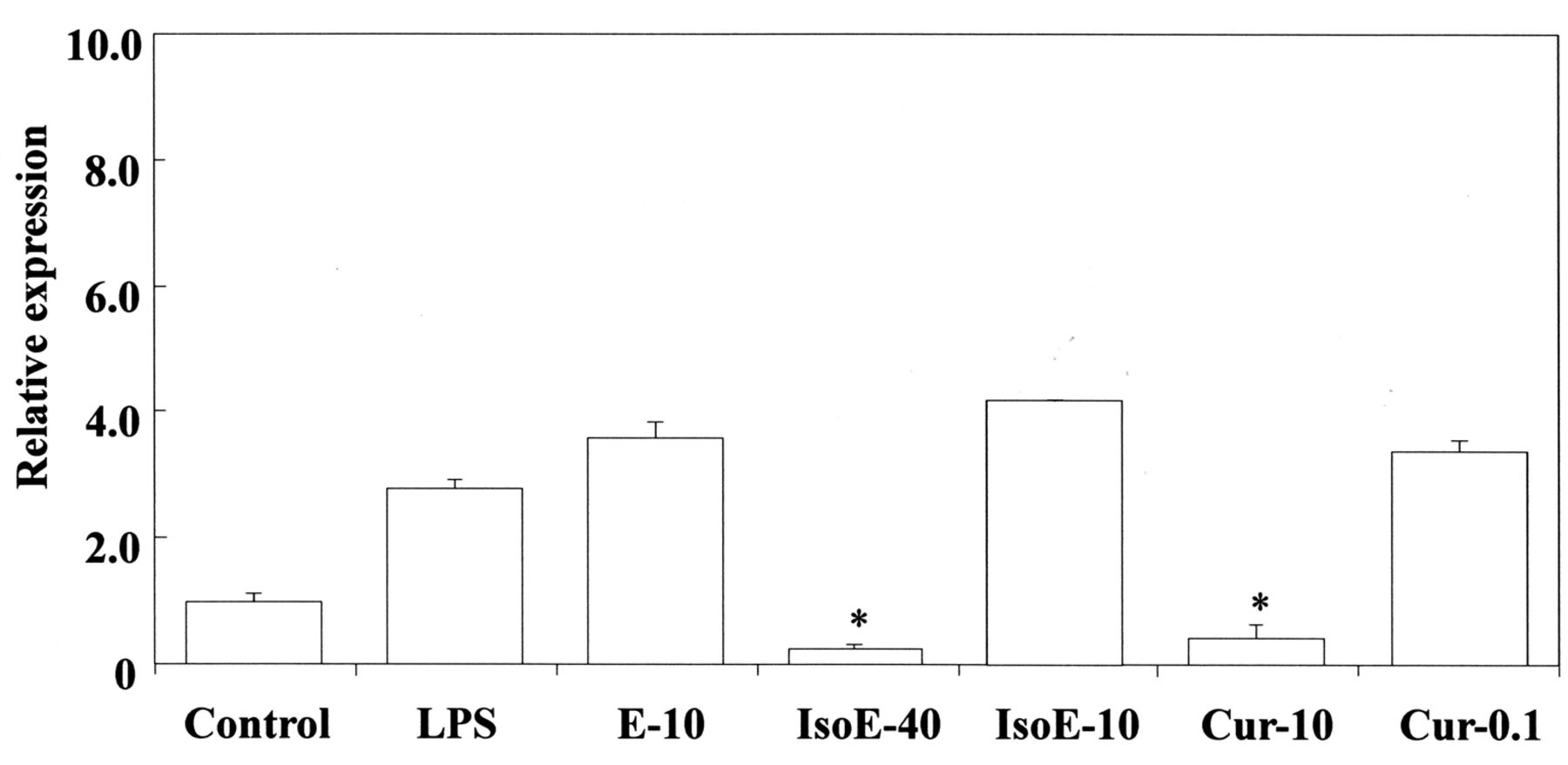
Next, we investigated the effects of eugenol-related compounds on HO-1 mRNA expression. The results are shown in Figure 5A and B. bis-Eugenol up-regulated the expression of HO-1 mRNA at both 1000 μM and 50 μM, whereas the other compounds had no effect at either of the two concentrations. The up-regulatory effect on HO-1 mRNA expression was observed in the absence of LPS. The gene expression was compared to that of the control untreated with the other compounds. Also, we investigated the effects of curcumin and isoeugenol on LPS-stimulated Cox-2 mRNA expression in RAW264.7 cells. The 50% inhibitory concentration of curcumin was about 2 μM, whereas that of isoeugenol was about 22 μM (Figure 6); those for bis-eugenol and eugenol were about 67 μM and 310 μM, respectively. The 50% inhibitory concentrations of magnolol and honokiol for PGF-stimulated Cox-2 mRNA expression in RAW264.7 cells were about 9 μM and 40 μM, respectively (Table I).

Stimulation of Cox-2 gene expression by isoeugenol, eugenol and curcumin (A), magnolol, honokiol and bis-eugenol (B) in RAW264.7 cells. The cells were incubated for 3.5 h with each eugenol-related compound at a concentration of 1000 μM or 0.1 μM, and then their total RNAs were prepared. Each cDNA was synthesized, and the expression level of Cox-2 mRNA was determined by real-time PCR and standardized against the expression of 18s rRNA. The results are presented as means±standard error (SE) of three independent experiments, SE<15%. Significant differences between samples for each eugenol-related compound were observed for stimulation of Cox-2 gene expression. *p<0.01 vs. control group. IsoE: Isoeugenol; E: eugenol; Cur: curcumin; Mag: magnolol; Hon: honokiol; bisE: bis-eugenol. 1,000:1,000 μM; 0.1: 0.1 μM.
Radical-scavenging activity. The results are shown in Table I. Eugenol-related compounds act as chain-breaking inhibitors. The first reaction is the limiting step of chain termination. The rate and activation energy of the reactions depends on the dissociation enthalpy of the phenolic OH group (O-H BDE). Therefore, as a reference, the O-H BDE values for these compounds are also shown in Table II.
Using eq. (1), the n value, i.e. the number of free radicals trapped by one mole of antioxidant moiety, was determined. The relative n value declined in the order curcumin (3.9) > honokiol (3.2) > bis-eugenol (2.8) > isoeugenol (2.6) > magnol (2.2) > eugenol (1.5). In general, fully oxidized phenols with two OH groups in the molecule show an n value of 4, whereas those with one OH group show an n value of 2. The n value for eugenol and isoeugenol would be less than 2, as their radical-scavenging activity is influenced by the intramolecular hydrogen bond formed between the phenolic OH and the adjacent methoxy groups (19, 20). However, in this study, the n value of 2.6 for isoeugenol may have been due to the formation of products which themselves act as inhibitors. This would have led to higher n values, which would vary according to the secondary nature of the reactions. Curcumin also has an intramolecular hydrogen bond between the phenolic OH and adjacent methoxy groups, but this compound showed an n value of about 4. In contrast, magnolol and honokiol, which are lignans with two phenolic OH groups, should have an n value of 4, but the actual value for magnolol was about 2, suggesting that this may be responsible for the intramolecular hydrogen bond formed between the OH groups. For honokiol, a dimer of p-allylphenol and o-allylphenol, the phenolic OH group at the o-position is probably first abstracted by peroxy radicals, since its O-H BDE is smaller than that of the OH group at the p-position (Table II). Honokiol can be fully oxidized via a two-electron process and is capable of having an n value of 4. Therefore, the radical-scavenging activity should be evaluated not only in terms of the n value but also the inhibitory propagation rate. Eugenol-related compounds having an intramolecular hydrogen bond between the phenolic OH and adjacent methoxy groups suppressed the propagation rate in the presence of a phenolic inhibitor (Rpinh) (Table II). In contrast, the Rpinh for both magnolol and honokiol without methoxy groups was similar to the propagation rate of the control (Rpcon). Thus, it was clarified that the Rp of eugenol, isoeugenol, bis-eugenol and curcumin is influenced by their methoxy groups. Using eq.2, kinh/kp declined in the order eugenol (7.8) > magnolol (6.8) > isoeugenol (5.69) > honokiol (4.8) > bis-eugenol (4.6) > curcumin (3.7). The inhibition rate constant (kinh, mol−1s−1) for honokiol, bis-eugenol and curcumin was less than that of eugenol, isoeugenol and magnolol, indicating that the former compounds scavenge radicals much more efficiently (see the n value) whereas their radical trapping time is shorter than that of the latter.
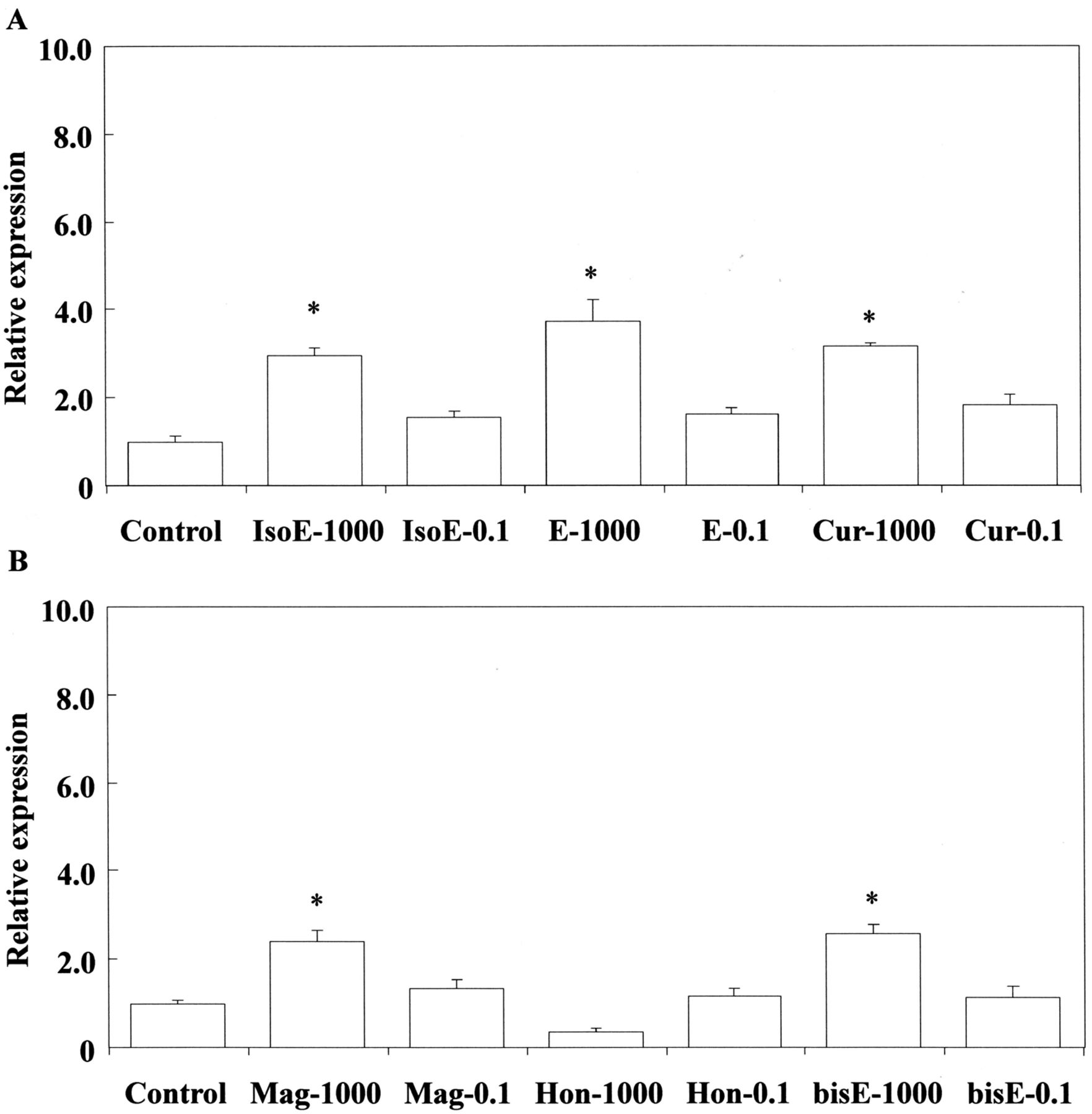
Stimulation of Nos2 gene expression by isoeugenol, eugenol and curcumin (A), magnolol, honokiol and bis-eugenol (B) in RAW264.7 cells. The cells were incubated for 3.5 h with each eugenol-related compound at a concentration of 1,000 μM or 0.1 μM, and then their total RNAs were prepared. Each cDNA was synthesized, and the expression level of Nos2 mRNA was determined by real-time PCR and standardized against the expression of 18s rRNA. The results are presented as means±standard error (SE) of three independent experiments, SE<15%. Significant differences between samples for each eugenol-related compound were observed for stimulation of Nos2 gene expression. *p<0.01 vs. control group. IsoE: Isoeugenol; E: eugenol; Cur: curcumin; Mag: magnolol; Hon: honokiol; bisE: bis-eugenol. 1,000:1,000 μM; 0.1: 0.1 μM.

Stimulation of Cox-2 (A) and Nos2 (B) gene expression by isoeugenol, eugenol, curcumin, magnolol, honokiol and bis-eugenol in RAW264.7 cells. The cells were incubated for 3.5 h with each eugenol-related compound at a concentration of 50 μM, and then their total RNAs were prepared. Each cDNA was synthesized, and the expression level of each mRNA was determined by real-time PCR and standardized against the expression of 18s rRNA. The results are presented as means±standard error (SE) of three independent experiments, SE<15%. Significant differences between samples for each eugenol-related compound were observed for stimulation of each gene expression. *p<0.01 vs. control group. IsoE: Isoeugenol; E: eugenol; Cur: curcumin; Mag: magnolol; Hon: honokiol; bisE: bis-eugenol. 50:50 μM.
Next, we investigated the effect of MMI on the antioxidant activity of eugenol-related compounds using the induction period method. MMI was used as a model of GSH. The results are also shown in Table I. The synergistic and antagonistic effects of these compounds were evaluated from the ratio of the IT values, i.e. the ratio of the IT for the eugenol-related compound/MMI mixture (1: 1 mole ratio) to that of a simple sum (eugenol-related compound + MMI). Antagonism (A), i.e. an IT ratio of <1, was observed for bis-eugenol and curcumin, and particularly the former. Weak but significant antagonism has also been observed previously for isoeugenol and curcumin (15). Conversely, eugenol showed an IT ratio of >1; indicating a synergistic (S) recycling effect. Such a recycling effect of eugenol has been reported previously (20, 21). A synergistic/antagonistic effect may be dependent on the ΔBDE value, i.e. the difference of BDE between O-H BDE and S-H BDE. When the O-H BDE value of phenolic antioxidants approximates the S-H BDE value of GSH, or when it is smaller than the S-H BDE value, synergism/antagonism can be explained by regeneration mechanisms (22). The ΔBDE, O-H BDE and S-H BDE values for eugenol-related compounds are shown in Table III. The ΔBDE value for eugenol was smaller than that for isoeugenol, and the ΔBDE of bis-eugenol (ΔBDE2nd) was smaller than those for other biphenols. The smallest ΔBDE2nd observed for eugenol-dimer may be related to the large antagonistic effect of bis-eugenol with GSH in biological systems. The curcumin/MMI mixture also showed an antagonistic effect, but was less marked than that of the bis-eugenol/MMI mixture. Phenoxyl radicals derived from peroxide-induced eugenol-related compound were reduced by interaction with MMI. In the last stage, eugenol-related compounds may be oxidized directly to quinone methides, possibly explaining their toxic effects and/or perhaps leading to formation of non-radical adducts. Isoeugenol also showed a relatively weak antagonistic effect, but greatly up-regulated Cox-2 and Nos2 gene expression at higher concentrations. We previously synthesized the reaction products of isoeugenol with ethanethiol, thiophenol, 2-mercaptothiazoline or 2-mercapto-1-methylimidazole in the presence of Lewis acid. Reaction products of isoeugenol bearing a SH group may tend to form via its radical-scavenging process because oxidation of isoeugenol produces a reactive benzyl radical as an intermediate (23). These findings suggest a possible interaction between biological macromolecules such as protein and DNA and isoeugenol, possibly resulting in allergens (24). In contrast, the O-H BDE2nd values of magnolol and honokiol were much higher than those of methoxyphenols such as curcumin and bis-eugenol, and also much higher than that of the S-H BDE for Cys-SH (Table III). Therefore, we did not study the antioxidant activity of the magnolol (or honokiol)/MMI mixture, as this would lead to possible additivity.

Stimulation of HO-1 gene expression by isoeugenol, eugenol and curcumin (A), magnolol, honokiol and bis-eugenol (B) in RAW264.7 cells. The cells were incubated for 3.5 h with each eugenol-related compound at a concentration of 1,000 μM or 0.1 μM, and then their total RNAs were prepared. Each cDNA was synthesized, and the expression level of HO-1 mRNA was determined by real-time PCR and standardized against the expression of 18s rRNA. The results are presented as means±standard error (SE) of three independent experiments, SE<15%. Significant differences between samples for each eugenol-related compound were observed for stimulation of HO-1 gene expression. *p<0.01 vs. control group. IsoE: Isoeugenol; E: eugenol; Cur: curcumin; Mag: magnolol; Hon: honokiol; bisE: bis-eugenol. 1,000:1,000 μM; 0.1: 0.1 μM.
The antioxidant activity of the eugenol-related compound/2-mercapto-1-methylimidazole, MMI mixture (1:1 molar ratio) determined by the induction period method for polymerization of methyl methacrylate (MMA) initiated by thermal decomposition of benzoyl peroxide (BPO). Effects of the antioxidant activity of eugenol-related compounds with or without MMI.
Phenolic O-H bond dissociation enthalpy (O-H BDE) and the difference between the O-H BDE of phenols and the S-H BDE of cysteine, ΔBDE=[O-H BDE] − [S-H BDE].
Discussion
According to Fukui's Frontier Orbital Theory (25), adduct formation occurs when a soft nucleophile denotes its high-energy electron to the empty lowest energy orbital (LUMO) of a soft electrophile. Therefore, the most relevant frontier orbital for electrophiles is the LUMO, whereas the HOMO is most important for nucleophiles (26). Softness is defined as the ease with which electron redistribution takes place during covalent bonding, and thus the softer the electrophile (more negative LUMO energy and higher σ value), the more readily it will form an adduct by accepting an outer shell electron from a soft nucleophile such as a sulfur atom. With respect to covalent reactions, relative softness (σ) reflects the ease with which electron redistribution occurs during adduct (covalent bond) formation. The softer the electrophile (large σ and more negative LUMO value), the faster it will accept electron density from a donating nucleophile (27). Our results suggested that the cytotoxicity of eugenol-related compounds for RAW264.7 cells was linearly correlated with their σ, ELUMO and ω values. Therefore, the cytotoxicity of these compounds may be due to their interaction with cellular GSH and macromolecules with sulfhydryl groups such as DNA and proteins. We previously reported that the cytotoxicity of eugenol-related compounds was linearly correlated with their η value (σ value) (28, 29). Also, the rank of the 50% effective concentration, E50, for the anti-inflammatory activity of methoxyphenols appeared to be dependent on their ω value. Similarly, we have previously reported that the anti-inflammatory activity of eugenol-related compounds was related to their ω value, the former increasing as the ω value increased (30). Also, magnolol and honokiol have been shown to exert anti-inflammatory activity, the rank order being much lower than curcumin but higher than bis-eugenol, judged by the estimation of COX-2 inhibitory activity by basing the ω value of these compounds (3).
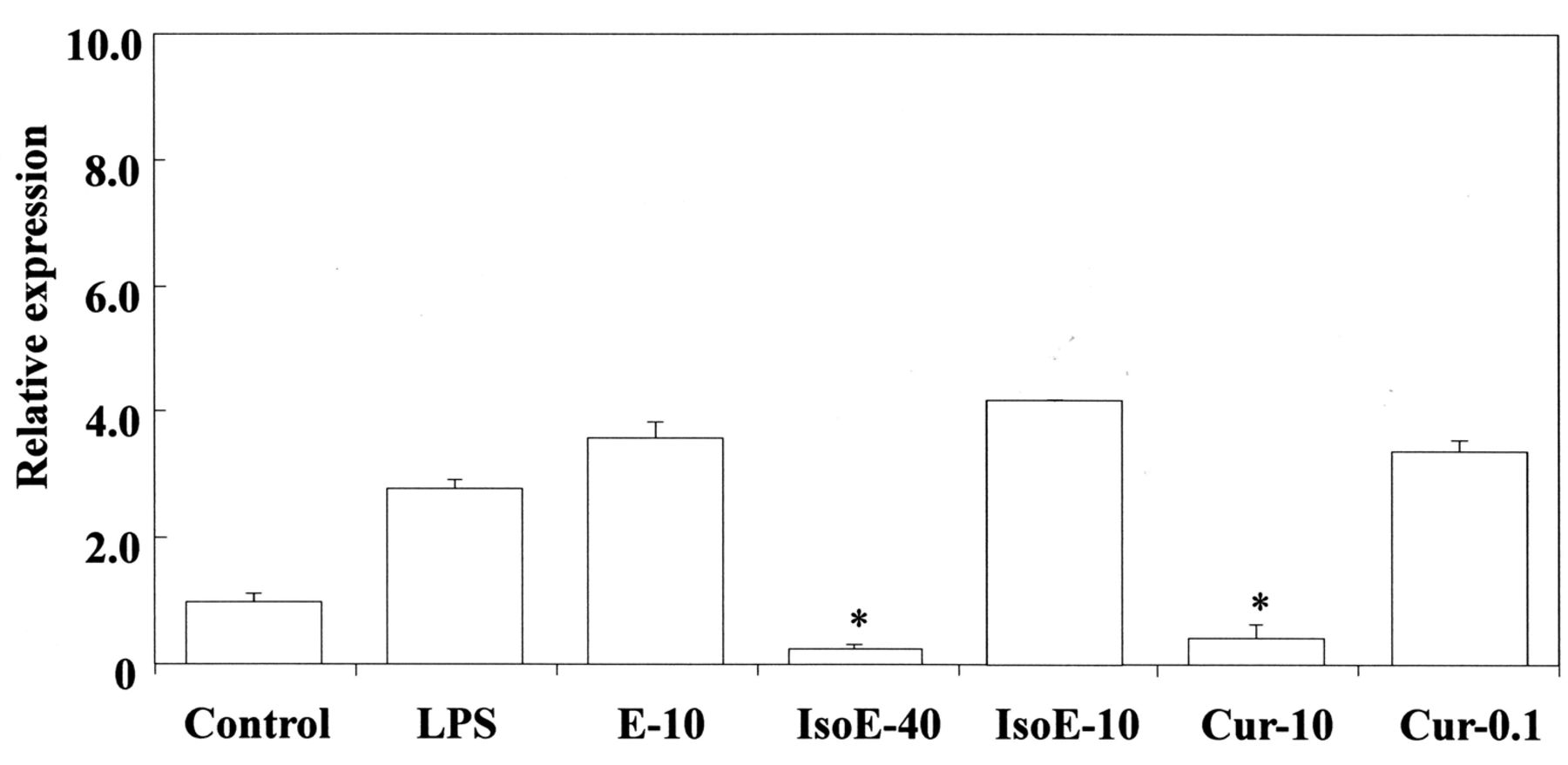
Inhibitory effects of isoeugenol and curcumin on LPS-stimulated Cox-2 gene expression in RAW264.7 cells. The cells were pretreated for 30 min with the indicated doses of isoeugenol, eugenol and curcumin. They were then incubated for 3 h with or without LPS at 100 ng/ml, and their total RNAs were prepared. Each cDNA was synthesized, and the expression level of Cox-2 mRNA was determined by real-time PCR and standardized against the expression of 18s rRNA. The results are presented as means±standard error (SE) of three independent experiments, SE<15%. Significant differences between samples for isoeugenol, eugenol and curcumin were observed for inhibition of Cox-2 gene expression. *p<0.01 vs. control group. IsoE: Isoeugenol; E: eugenol; Cur: curcumin. 40:40 μM; 10:10 μM; 0.1: 0.1 μM.
On the other hand, it is well known that free radicals such as the superoxide anion, hydroxy, alkyl, alkoxy, peroxy and nitric oxide radicals can emerge spontaneously and/or be generated through enzyme catalysis or in biological systems; eugenol-related compounds, which are chain-breaking antioxidants, preferably scavenge free radicals. Therefore, much interest has been focused on the antioxidant activity of eugenol-related compounds for prevention of radical-mediated inflammatory activity and diseases (3). The kinetic antioxidant activity of phenolic compounds was analyzed previously in terms of their ability to inhibit the oxidation of styrene by proxyl radicals generated from styrene by azo-bis-isobutyronitrile, and these inhibitory effects were similar to those occurring in cells (31). We have also studied the kinetic antioxidant activity of methoxyphenols and tert-butylphenols using the induction period method for polymerization of MMA initiated by thermal decomposition BPO, and found a possible link between cytotoxicity in human gingival cells and the kinh/kp values of these compounds; a certain correlation was evident between the logarithm of kinh/kp and the O-H BDE value (29). These findings suggested that phenol-induced cytotoxicity may be due to radical-mediated mechanisms.
In the present study, eugenol-related compounds, particularly isoeugenol, eugenol and curcumin, up-regulated the expression of Cox-2 and Nos2 mRNA at the high cytotoxic concentration of 1,000 μM. At 50 μM, magnolol, honokiol, curcumin and bis-eugenol, and particularly magnolol, up-regulated Cox-2 and Nos2 gene expression to a greater degree than the monophenols, isoeugenol and eugenol. As biological systems are heterogeneous environments, hydrophobicity is a more desirable property than hydrophilicity. Therefore, biphenols with high hydrophobicity may have a more pronounced effect on gene expression than monophenols. The high expression of the Cox-2 and Nos2 genes elicited by magnolol, which has an intramolecular hydrogen bond between the di-ortho-hydroxy groups, may have been caused by formation of its semiquinone radical (-OH-O*) and subsequent quinone methide formation (32). The rank order of O-H BDE1st (eV) was honokiol (345) > curcumin (344) > bis-eugenol (337) > magnolol (328) (Table III). Magnolol may be preferentially oxidized due to its small O-H BDE1st value and its n value was 1, possibly resulting in the high expression of the Cox-2 and Nos2 genes. Curcumin and bis-eugenol, and particularly the latter, showed a small BDE2nd=[O-H BDE] −[S-H BDE], possibly due to antagonistic effects occurring between the phenolic antioxidant and GSH. For GSH, co-antioxidant effects may up-regulate HO-1 gene expression, although the mechanism remains unknown. GSH is well known to be an abundant thiol present in mammalian cells, performing a number of functions; it maintains the essential thiol-states of proteins by preventing oxidation of sulfhydryl groups or by reducing disulfide bonds induced by oxidative stress by scavenging free radicals (33, 34). The complex nature of electrophiles, which are reactive eugenol-related compounds, is illustrated by the fact that GSH is depressed by quinone methides, which are formed by radical oxidation of phenol (21). The toxic effects of eugenol are mediated through its quinone methide metabolite. Quinone methide formation by phenolic compounds is associated with both cytoprotective as well as cytotoxic effects (35). The biological targets of quinone methides derived from radical-oxidation of phenols could exert cytotoxic effects through covalent modification of biological macromolecules such as proteins and/or DNA, as well as depletion of GSH leading to an altered redox balance within cells (36). Alternatively, cytoprotection could occur due to formation of quinone methide through covalent modification of Kelch-like ECH-associated protein 1 (Keap1), leading to activation of nuclear factor erythroid 2-related factor 2 (Nrf2) and subsequent induction of detoxification enzymes such as nicotinamide adenine dinucleotide phospho (NAD(P)H):quinone oxidoreductase (NQO1) and heme oxygenase (36).
COX-2 is selectively induced by proinflammatory cytokines at sites of inflammation, and NOS2 is involved in the inflammatory process (37). HO-1 is the inducible isoform of heme oxygenase that catalyzes the degradation of heme into biliverdin, iron, and carbon. HO-1 has also been shown to be an oxidative stress-responsive gene that may exert important roles in cytoprotection and the resolution of inflammation to maintain tissue homeostasis (12, 38). In most cell types, HO-1 is inducible by heme, radiation, cytokines, inflammatory stimuli, and oxidative and heat stress (39). Recent studies have demonstrated that overexpression of HO-1 prior to stimulation with LPS markedly inhibits the production of subsequent inflammatory mediators such as nitric oxide (NO), interleukin-6, and monocyte chemoattractant protein (MCP-1) (40, 41). Although, unlike nitric oxide (NO•), CO only weakly activates soluble guanylate cyclase, CO has been found to broadly suppress LPS-induced immediate-early genes that initiate and propagate inflammation involving pivotal transcription factors (e.g. ECG family members, Activating Transcription Factor 3) and cytokines (e.g. interleukin (IL)-1β, IL-8 and tumor necrosis factor (TNF)-α) (42).
Our present study demonstrated that bis-eugenol down-regulated the expression of the Nos2 gene in RAW264.7 cells. bis-Eugenol was able to suppress production of nitric oxide. It has also been reported that CO derived from HO-1 may reduce the production of ROS and the function of NAPDH oxidase, as well as being a potent inhibitor of iNOS and Nf-ĸB (43). Many previous studies have indicated that curcumin affects HO-1 expression in hepatoma cells (44) and fibroblasts (45), whereas magnolol does so in RAW264.7 cells (46). In the present study, however, none of the eugenol-related compounds except for bis-eugenol induced HO-1 expression in RAW264.7 cells. The signaling mechanisms involved in HO-1 expression may vary according to cell type and the inducers involved. LPS-stimulated expression of the Nos2 gene in RAW264.7 cells was potently inhibited by bis-eugenol, but not by eugenol, whereas that of Tnf-α mRNA was not affected by 100 μM bis-eugenol (data not shown). LPS-stimulated expression of Cox-2 mRNA was inhibited by eugenol-related compounds, particularly curcumin, which had the highest ω value. This may also be related to the structure of biphenols and the α,β-unsaturated carbonyl group of curcumin. Compounds like bis-eugenol, such as curcumin, magnolol and honokiol, preferentially suppress Nfĸ-B activation, whereas bis-eugenol itself elicits a weak suppressive effect (5). In the present study, bis-eugenol required HO-1 mRNA expression. Magnolol has a bis-eugenol-like structure and has been reported to exhibit beneficial effects of treating bone diseases associated with excessive osteoclastogenesis (46); magnolol can increase HO-1 expression and Nrf2 activation in receptor activator of Nf-ĸB ligand (RANKL)-stimulated cells.
Our result strongly suggested that bis-eugenol may have therapeutic applications against inflammatory and anticancer activity via regulation of HO-1 mRNA. Despite the large disparity between experimental conditions and natural biological systems, studies of the kinetic and synergistic/antagonistic antioxidant effects of eugenol-related compounds may help to explain the biphasic mechanism of the antioxidant/prooxidant activity. The toxicity of antioxidative eugenol-related compounds may be both intrinsic, and also attributable to metabolites such as quinonemethides, which have antioxidant-derived radicals (3). These phenolic compounds can exert multiple pharmacological functions through a single structure, involving radical-mediated dimer formation and quinonemethide formation. Therefore, theoretical parameters such as the σ, ω and O-H BDE values may be useful for predicting the anti-inflammatory/pro-inflammatory activity of eugenol-related compounds in biological systems. Further studies will be needed to clarify the mechanisms involved in the anti-inflammatory and pro-inflammatory activity of eugenol-related compounds.
Acknowledgements
This work was supported by a grant-in-aid for scientific research (No.15K11266) from the Ministry of Education, Science, and Culture of Japan. The Authors would also like to thank Kadoma Y. and Ito S. for measurement of the induction time and calculations of theoretical parameters, respectively.
Footnotes
This article is freely accessible online.
- Received May 19, 2017.
- Revision received June 5, 2017.
- Accepted June 6, 2017.
- Copyright© 2017, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}