


Abstract
Background/Aim: Peyronie's disease (PD) is a fibrotic entity for which the pathogenetic mechanism remains unclear and if resulting in severe deformity, its treatment is only surgical. In this study we investigated the possible role of insulin-like growth factor 1 (IGF1) expression in the pathogenesis of PD. Materials and Methods: Tissue samples were obtained from plaques of 24 patients with PD. The expression of IGF1 isoforms was investigated using quantitative real-time polymerase chain reaction and immunofluorescence. Results: All IGF1 isoform gene expression (Ea, Eb and Ec) were found significantly decreased in the affected tunica albuginea, compared to normal tunica albuginea, with Ec showing the greatest decrease. Staining of tissue sections with an antibody against IGF1Ec confirmed greater expression of IGF1Ec isoform in normal tunica albuginea. Conclusion: The expression of all IGF1 alternative spliced isoforms is decreased in patients with PD, suggestive of its possible participation in the pathophysiology of PD.
Peyronie's disease (PD) is a fibrotic entity involving the tunica albuginea and the adjacent tissue of penile corpora cavernosa. It is characterized by the formation of a fibrotic plaque, which usually leads to penile curvature and, if severe, subsequently to inability for vaginal penetration (1). Despite the fact that PD was well described more than 250 years ago and is a relatively common disease, affecting 0.4 to 8.9% of the male population (1, 2), its pathogenetic mechanism remains enigmatic. The current hypothesis suggests that repetitive microvascular injury or trauma to the tunica albuginea triggers an inflammatory process (3, 4). The most commonly evaluated factors regarding the pathogenesis of PD are fibrin and transforming growth factor-beta 1 (TGFβ1) (5), that is overexpressed in PD plaque (6). It has been established that differentiation of tunica albuginea fibroblasts into myofibroblasts under the influence of TGFβ1 is the cornerstone of PD pathogenesis, resulting in excess collagen synthesis and plaque formation. In the typical wound-healing process, myofibroblasts are led to apoptosis and there is normal healing of tissue, but in PD, myofibroblasts escape apoptosis, leading to fibrosis, collagen accumulation and tissue contraction. Unfortunately, these models do not reflect the natural history of the disease, thus leaving unanswered questions regarding the pathogenesis of PD. These gaps in the pathogenetic cascade and the fact that all available medical treatment options have been proven ineffective (7), with the exception of promising intralesional distribution of collagenase (8), leave surgical correction of PD as the last hope for all patients with severe deformation. Therefore, it is mandatory to fill these blanks in the knowledge of PD pathogenesis in order to provide suffering patients a satisfactory treatment along with a better understanding of the disease.
Insulin-like growth factor 1 (IGF1) is an important regulator of growth, survival and differentiation in many organs and tissues (9). IGF1 gene contains six exons, which in humans give rise to three mRNA isoforms by alternative splicing, namely IGF1Ea, IGF1Eb and IGF1Ec (10, 11), with expression of the latter shown to be enhanced during regeneration and repair of both skeletal and heart muscles (12-14). Since the IGF1 isoforms have been implicated in tissue-repair mechanisms, we investigated the possible role of the IGF1 isoforms in the pathogenesis of PD.
Materials and Methods
Patients. A total of 24 patients, aged 36 to 72 (mean=66.7) years with PD undergoing surgical correction of their curvature were enrolled in this study. All of them gave their written consent to this study, which was approved by the Ethics Committee of the National and Kapodistrian University of Athens School of Medicine (approval number 3725/16-12-2009), and all experimental procedures conformed to the Declaration of Helsinki.
Primer sequences used in quantitative real-time polymerase chain reactions.
Specimens were obtained directly from the palpable plaque during surgery after degloving of the penis. As control, specimen from normal tunica albuginea was obtained from the same patient. Additional tissue specimens from every patient were harvested, both from plaque and normal tunica albuginea to confirm histologically the clinical diagnosis of PD and normal tunica albuginea, respectively. Biopsy samples were immediately transferred in Ambion RNAlater (Thermo Fisher Scientific Inc., Waltham, MA, USA), frozen at site at at −80°C and the histological specimens were placed in formaldehyde solution for fixation.
Histology. For histological analysis, tissue specimens from both the affected site of plaque and normal tunica albuginea of the same patient were obtained, fixed overnight in 10% neutral buffered formalin and embedded in paraffin. De-waxed sections (4 μm) were stained using hematoxylin and eosin (H&E) and photographed using a Nikon Eclipse 80i camera (Nikon, Shinagawa, Tokyo, Japan).
RNA isolation and cDNA synthesis. Once all tissues were collected, RNA extraction was performed. The RNAlater reagent was removed and 1 ml of TRIzol Reagent (Invitrogen, Carlsbad, CA, USA) was added to each tissue. The samples were left at room temperature for 5 min and were subsequently homogenized using a portable homogenizer (IKA, Staufen, Germany). Following homogenization, the samples were left for 5 more min at room temperature and 200 μl of chloroform (Applichem GmbH, Darmstadt, Germany) was added. The the samples were then shaken vigorously for 2 min. Afterwards, an equal volume of ethanol was added to each sample that was then thoroughly mixed. The whole volume was loaded into spin columns (Ultra Clean Tissue and Cell RNA isolation kit; MoBio Laboratories, Inc., Carlsbad, CA, USA) and RNA was eluted in 20 μl final volume of nuclease-free water according to the instructions of the manufacturer. The quantity and quality of the RNA was assessed using a biospectrophotometer (Shimadzu, Kyoto, Japan).
In total, 1 μg of RNA was amplified in order to create cDNA using the Protoscript II first strand cDNA synthesis kit (New England Biolabs, Ipswich, MA, USA) and according to the instructions of the manufacturer. Following the reverse transcription reaction, samples were stored at −20°C until further processing.
Quantitative real-time polymerase chain reaction (qPCR). All qPCR reactions were performed at 20 μl final volume using 50 ng as a cDNA template. Briefly 50 ng of cDNA was added to each well of a 96-well PCR plate, along with Biorad universal IQ SYBR mix 2x (Bio-Rad Laboratories, Inc., Hercules, CA, USA) gene-specific forward and reverse primers (Integrated DNA Technologies, Inc., Coralville, IA, USα) at 500 nM final concentration and nuclease-free water. Primers used are displayed in Table I. All genes were amplified using the Bio-Rad IQ5 real-time PCR cycler under the following conditions: one cycle for 10 min at 95°C, 45 cycles of 15 s at 95°C 30 s at 62°C and 30 s at 72°C followed by 51 cycles of 0.5°C gradient heating for melting-curve analysis. Amplicon specificity was confirmed by melting-curve analysis for specific peaks and by agarose gel electrophoresis. Results were analyzed using the DDct method for relative quantification using glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as reference gene.
Immunofluorescence microscopy. Tissues were harvested and stored in paraffin blocks. Sections of 10 μm were cut and fixed into glass cover slides. The samples were then deparaffinized by use of xylene and slides were rehydrated in decreasing concentrations of ethanolic solutions. Once rehydrated, the slides were incubated for 15 min in 0.1 M glycine, washed twice with phosphate-buffered saline (PBS) followed by 10 min incubation with 0.1% triton-X in PBS for membrane permeation. Then the slides were blocked for 30 min at room temperature with 5% goat serum and subsequently incubated with 1:1,000 dilution of a IGF1 Ec-specific antibody overnight at 4°C (15). The following day, the slides were washed three times with PBS and incubated for 1 h with 1:1,000 dilution of Anti-rabbit IgG (H+L), F(ab’)2 Fragment Alexa Fluor® 488 conjugate (Thermo Fisher Scientific Inc.). Slides were once again washed three times with PBS followed by the addition of 20 μl of SlowFade® Gold Antifade Mountant (Thermo Fisher Scientific Inc.) with 4’,6’-diamidino-2-phenylindole (DAPI) and coverslipped. The same procedure was followed using antibody to TGFβ1. Slides were then viewed under a Nikon Eclipse fluorescent microscope (Nikon, Shinagawa, Tokyo, Japan).

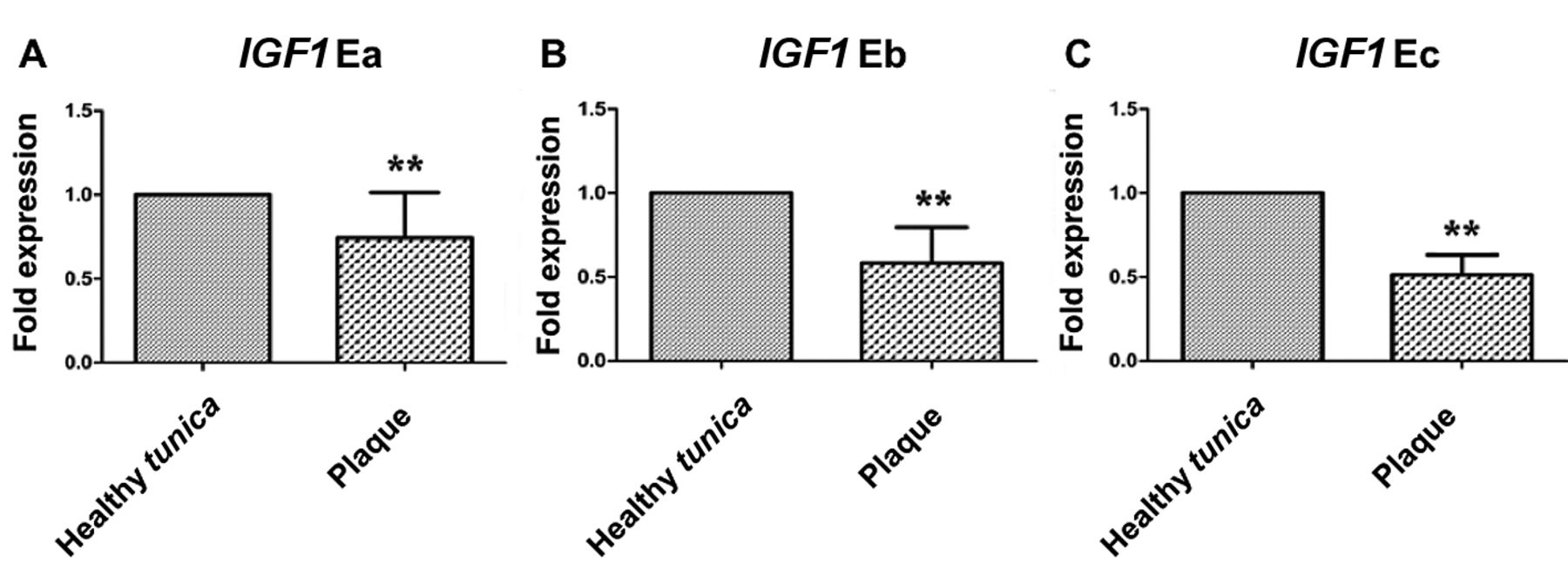
Relative mRNA expression of Insulin-like growth factor 1 (IGF1) splice variants in specimens from plaque and normal tunica albuginea. Specimens from plaque had significantly lower transcript expression than normal tunica albuginea (**p<0.01).
Representative melting curves of the quantitative real-time polymerase chain reaction specific products of Insulin-like growth factor 1 (IGF1) Ea (A) IGF1Eb (B), IGF1Ec (C) and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (D).
Statistical analysis. For statistical analysis, Graphpad Prism v5.0 was used (GraphPad Software, Inc., San Diego, CA, USA). All results were submitted to normality tests followed by paired t-test statistical analysis. Statistical significance was considered at p<0.05.
Results
IGF1 expression. Expression of all IGF1 isoforms (Ea, Eb and Ec) were found to be significantly decreased in the affected tunica albuginea, where plaque had been formed, compared to normal tunica albuginea, as assessed by qPCR methods (Figures 1 and 2).
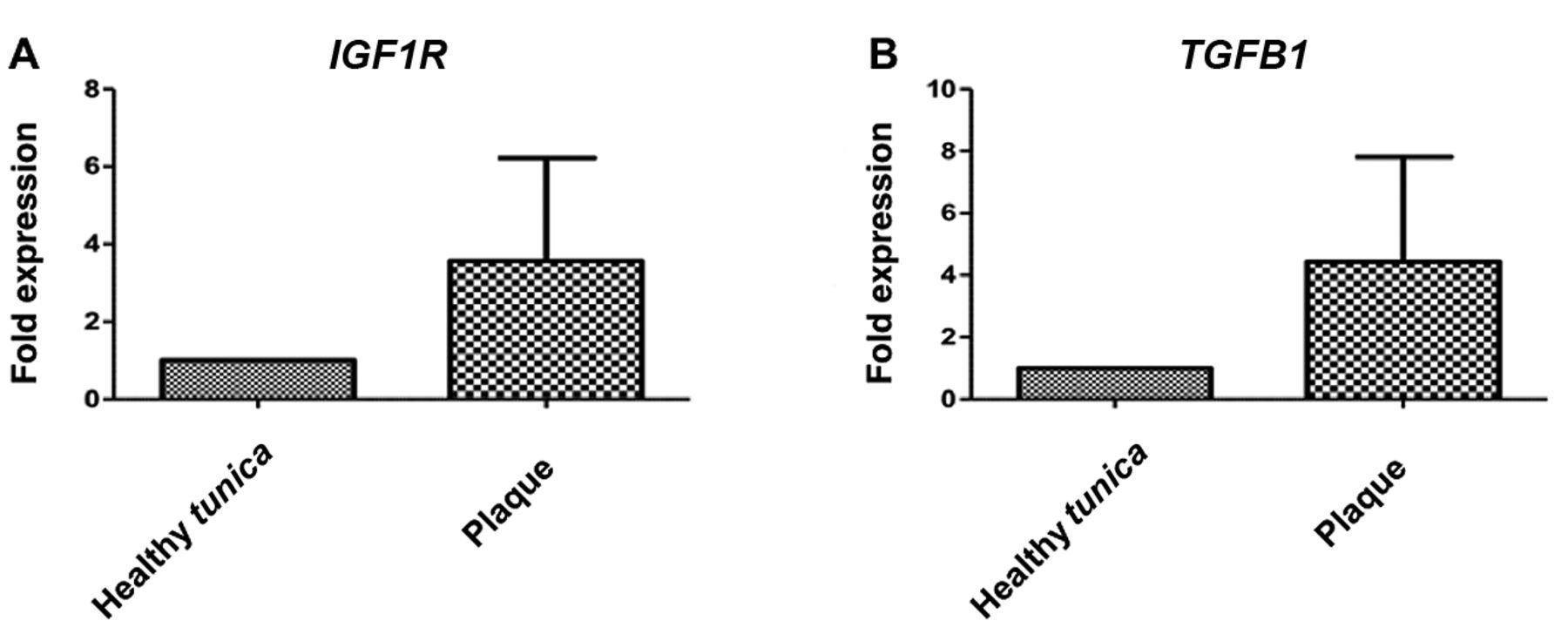
IGF1R expression. Real-time PCR showed that IGF1R mRNA was overexpressed in plaque samples, but expression was not significantly different from that of normal tunica albuginea, mainly due to high variation among samples (Figure 3).
TGFB1 expression. TGFB1 gene expression was found to be increased, as previously documented, but the increase was not statistically significant, due to high variation among samples (Figure 4).
Immunofluorescence microscopy. Qualitative analysis with immunofluorescence revealed that IGF1Ec expression was lower in the affected tunica albuginea compared to control tissue. Staining of placenta samples was used as a control. Furthermore, staining for TGFβ1 was stronger in the affected tunica albuginea (Figure 5).
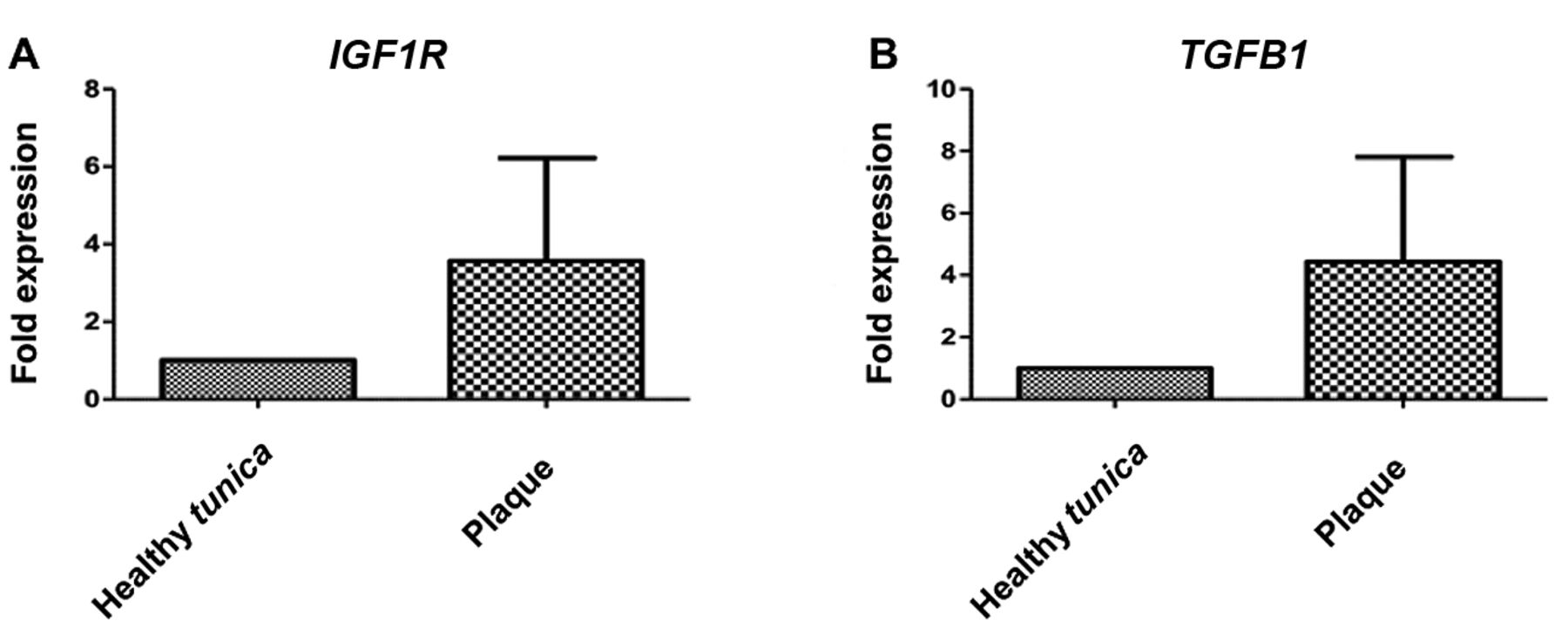
Relative mRNA expression of insulin-like growth factor 1 receptor (IGF1R) (A) and transforming growth factor beta 1 (TGFB1) (B) in specimens from plaque and normal tunica albuginea.

Hematoxylin-eosin staining of tissue. A: Normal tunica albuginea, normal stroma without evidence of collagen alterations (×10). B, C: Dense hyperplastic collagen bundles with evidence of early ossification (B: ×10, C: ×40).

Immunofluoresence staining with antibody to insulin-like growth factor 1 receptor (IGF1R) Ec and and transforming growth factor beta 1 (TGFβ1) (green staining) in tissue segments of placenta, affected tunica albuginea and normal tunica albuginea. Nuclear counterstain with 4’,6’-diamidino-2-phenylindole (blue staining) (magnification ×20).
Discussion
In this study, we demonstrated a significant suppression of the expression of IGF1 isoforms in tunica albuginea derived from Peyronie's plaque as compared to healthy tunica albuginea.
IGF1 plays a pivotal role in numerous developmental processes, such as embryonic development, postnatal growth, maintenance of homeostasis and tissue repair (16, 17). It mainly acts as a mitogen, differentiation factor and survival factor, inhibiting apoptosis (18-20). IGF1 facilitates wound healing by stimulating fibroblast proliferation and enhancing collagen synthesis (21), while reduced expression of IGF1 and its receptors has been associated with impaired wound healing (22, 23). IGF1 is implicated in other fibrotic diseases, including liver and pulmonary fibrosis, muscle damage and skin scar formation (24, 25). Data supporting the role of IGF1 in wound healing are quite conflicting, since a causative role of increased IGF1 expression has been suggested in the pathogenesis of hypertrophic scars (26), while it is differentially expressed in acute and chronic wounds. In fact, IGF1 is overexpressed in pulmonary fibrosis and acute lung injury (27-29), while it is down-regulated in chronic injury (30). The development and progression of chronic wounds is associated with excessive inflammation, which fails to resolve (31). Improved burn-wound healing has been associated with IGF1-induced attenuation of pro-inflammatory cytokines, which may improve wound healing by reducing prolonged local inflammation. Moreover, IGF1 contributes to normal tissue repair and remodeling in damaged skeletal (24, 32) and cardiac muscle (25, 33), while it was recently proposed that local IGF1 administration would be beneficial for chronic wounds.
Our findings are in accordance with the natural progression of the disease, that includes transformation of fibroblasts into myofibroblasts, avoiding apoptosis, thus leading to collagen deposition and scar formation. It seems that as in other chronic wounds, normal wound healing process is impaired in PD and low expression of all IGF1 isoforms, especially IGF1 Ec, explains partially why plaque formation is favored. Previous findings from our laboratory suggest that IGF1 isoforms play a distinct role in several pathophysiological processes (13, 14, 34), nevertheless the role of IGF1 in PD has never been investigated to date.
Our study has certain important limitations. We used control specimens from the same patient, since on the one hand we could not harvest normal tunica albuginea from healthy individuals and on the other, we could not use specimens from patients undergoing penile prosthesis implantation or patients with penile cancer undergoing penile amputation, due to underlying conditions which could influence the results. During specimen harvesting, we did not have the opportunity of obtaining enough material in order to perform further tests at the protein level, but we used immunofluorescence in order to verify our PCR findings. We found IGF1 Ec expression was lower in the affected tunica albuginea compared to normal, while TGFβ1 was increased. TGFβ1 is one of the most potent profibrotic stimuli (35, 36), which transforms the process of scarless wound healing into fibrotic scar formation (37, 38). Specimens from human placenta were used as control samples for immunofluorence since IGF1 is present in syncytiotrophoblast and cytotrophoblast at all stages in gestation (39). Since therapeutic administration of IGF1 has been proposed in other fibrotic entities (40), the next logical step is to create cell cultures of myofibroblasts derived from plaque sites of patients with PD or to design a new animal model of induced PD and administer IGF1 locally in order to define whether IGF1 advances normal wound healing, hence proposing a novel therapeutic agent which can be added to our armamentarium.
Acknowledgements
The Authors would like to thank Dr. Rados Djinovic, Chairman of the Sava Perovic foundation and President of ESGURS for his valuable contribution in collecting tissue samples.
- Received December 21, 2015.
- Revision received March 22, 2016.
- Accepted March 23, 2016.
- Copyright © 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}