


Abstract
Patients with QT prolongation have delayed cardiac repolarization and may suffer fatal ventricular arrhythmias. To determine the role of cytokines in causing this syndrome, we reviewed reports on patients with rheumatoid arthritis, psoriasis and other inflammatory conditions. These patients frequently have prolonged QT, which correlates with increases in tumor necrosis factor alpha, and interleukin-1β and 6. Studies in experimental models have shown that these cytokines act through stimulation of reactive oxygen species. Our review of data on phospholipidosis and on QT-shortening agents suggests a key role in QT prolongation for the ceramide/sphingosine-1-phosphate rheostat. We conclude that the cause of prolonged QT in inflammatory conditions is cytokine induction of reactive oxygen species and then ceramides, and believe that QT-prolonging agents bypass initial steps of this pathway and directly affect ceramides. Since both pro-inflammatory cytokines and numerous medications cause QT prolongation and ventricular arrhythmias by this mechanism, extra caution is needed when using these agents in patients with inflammatory conditions.
- Prolonged QT
- repolarization
- ventricular arrhythmia
- hERG
- potassium channel
- inflammation
- cytokines
- TNFα
- interleukin-1β
- interleukin-6
- reactive oxygen species
- ceramide
- sphingosine-1 phosphate
- review
An increase in the levels of pro-inflammatory cytokines plays a role in the morbidity of numerous diseases. Marked elevations in levels of tumor necrosis factor-alpha (TNFα), and interleukins IL1β and IL6, in particular, have been found in diseases as diverse as diabetes (1), Alzheimer's disease (2), inflammatory bowel disease (3) and cancer (4). When elevation of these cytokines becomes massive, a ‘cytokine storm’ may develop and cause death of the patient (5-7). These pro-inflammatory cytokines are also found to be increased in autoimmune diseases, and blockers of these cytokines are now standard therapies for patients with rheumatoid arthritis (RA) and many other diseases (8-10).
In long QT syndrome, there is delayed repolarization secondary to abnormalities in the potassium, sodium and calcium ion channels in myocardial cells. This may be caused by a genetic abnormality, coexisting cardiac diseases, hypokalemia or hypomagnesemia, or by one of numerous pharmaceutical agents, including commonly used antihistamines, sedatives, antibiotics, antiarrhythmics and psychiatric drugs. Ventricular tachycardia and sudden death secondary to delay in repolarization may occur (11-15). There is now increasing evidence suggesting that overexpression of IL1β, IL6, TNFα, and other cytokines plays a role in the pathogenesis of the long QT syndrome.
Data from Patient Trials
It has been observed that patients with rheumatoid arthritis (RA) have a markedly increased incidence of QT prolongation and an increased mortality secondary to cardiac arrhythmias (16-19). Patients with RA are thought to have twice the risk of death from ventricular arrhythmias compared to the normal population (17), and a 50 ms increase in QT in patients with RA doubles the risk for all-cause mortality (18). This increase in mortality risk also correlates with increases in the levels of inflammatory markers (17). Patients with other autoimmune diseases, such as psoriasis (20), systemic lupus erythematosus (21-24) and inflammatory bowel disease (25), have also been shown to have an increased incidence of QT prolongation. Prolonged QT from myocarditis secondary to infectious diseases is also well-described (26-31).
In all of these inflammatory conditions, marked increases in IL1β, IL6, TNFα, in particular, are seen. Adlan et al. have shown that the levels of these three cytokines and of the anti-inflammatory cytokine IL10 in patients with RA correlate directly with the extent of prolongation of the QT interval (32); and that elevation of these cytokines correlates with increased QT prolongation even in patients without disease (33). In a study of 1200 patients with heart failure, Deswal et al. showed the chance of dying could be predicted by the extent of elevations of TNFα and IL6 (34). The ‘electrical storm’ of recurrent ventricular arrhythmia seen with cardiac disease is also associated with elevated levels of cytokines and other inflammatory markers (35, 36). Furthermore, prolongation of the QT interval has been reported as a side-effect of therapy in anticancer trials of the pro-inflammatory cytokines, IL18 and interferon-γ (IFNγ) (37, 38).
In a recent study, Lazzerini et al. noted that patients with RA treated with tocilizumab, an IL6 blocker, had shortening of their previously prolonged QT interval, and this shortening was associated with concomitant decreases in inflammatory markers. In this trial of 17 patients with severe RA, 76% (13/17) had QT intervals of greater than 440 ms (mean±SD=452.3±35.8 ms) before starting therapy. After six months of therapy, only 29% (5/17) had a QT interval greater than 440 ms (mean±SD=428.1±34.3 ms). Decreases in the levels of TNFα and C-reactive protein correlated with the decrease in QT (39).
Cytokines: Data from Experimental Models
Transgenic mice which overexpress TNFα have a markedly prolonged action potential duration and experience recurrent ventricular arrhythmias (40). Studies in animal models or in cultured cardiomyocytes support the hypothesis that elevations in cytokines can cause prolonged QT. Wang et al. studied the effects of TNFα on canine cardiomyocytes using whole-cell patch clamp techniques. Marked inhibition of the rapid delayed-rectifier K+ current (IKr) was seen. TNFα was also found to reduce the human ether-à-go-go-related gene (hERG) current in HEK 293 cells expressing hERG. The suppressive effect of TNFα was concentration-dependent, and could be blocked by co-administration of an antibody to TNFα (41). The effect was thought to be due to TNFα stimulation of reactive oxygen species (ROS) since it was also blocked by vitamin E and by the superoxide dismutase mimic manganese (III) tetrakis (4-benzoic acid) porphyrin (MnTRAP). Kawada et al. reported that rat cardiomyocytes incubated with TNFα showed a decreased transient outward potassium current (Ito), and reductions in Kv4.2 and K+ channel-interacting protein-2 (42). Fernandez-Velasco found similar results, and also suggested that TNFα affects cardiac K+ channels through stimulation of ROS. Again, the effect of TNFα on QT prolongation was blocked by vitamin E and by MnTRAP (43). Petkova-Kirova et al. studied mice overexpressing TNFα in the heart, using whole-cell voltage-clamp recording of K+ current, and observed a 50% decrease in Ito, as well as a major decrease in the slow component of the delayed rectifier potassium current (IKs) (44). In another study, Grandy and Fiset reported that ventricular myocytes from mice chronically treated with TNFα to achieve blood levels equivalent to those seen in patients with congestive heart failure or HIV exhibited significantly reduced Ito and IKr (45). Li and Rozanski reported that IL1 significantly increased the duration of the action potential and the effective refractory period in excised papillary muscle, and increased the L-type calcium current (ICaL) in isolated cardiomyocytes. The increase in action potential duration in papillary muscle was blocked by aspirin and by indomethacin (46). Hagiwara et al. used whole-cell patch clamp recordings of the ventricles of mice exposed to IL6, and found a marked increase in ICaL (47).
Phospholipidosis and Ceramides
Another clue to the mechanism by which prolonged QT may occur is the similarity between the list of medications that cause QT prolongation and the list of those that cause phospolipidosis (PLD), the excess accumulation of intracellular phospholipids due to abnormalities in lysosomal lipid metabolism. Sun et al. screened 4,090 approved and investigational agents and found that 209 induced PLD in Hep G2 cells. Seventy-seven percent of these compounds were hERG channel blockers, and when steroidal drugs were excluded, 87% of PLD-inducing drugs were found to be hERG channel blockers (48). Abnormalities in cytokine production have been seen in phospholipidotic cells. We noted a study by Reasor et al. that showed marked enhancement of IL6 and TNFα release by rat phospholipidotic alveolar macrophages after lipopolysaccharide (LPS) stimulation (49). Likewise, Reinhart et al. reported that amiodarone, an agent which causes both PLD and prolonged QT, stimulated the release of TNFα and transforming growth factor (TGF)-β from pulmonary alveolar macrophages (50). Similar results have been reported by Masubuchi et al. (51). On the other hand, Munic et al. reported that the phospholipidotic potential of various macrolide antibiotics correlated with a reduction in IL6 levels in LPS-stimulated cells. Small increases in TNFα levels were seen with increasing PLD (52).
More importantly, although the exact biochemical mechanism by which PLD occurs is not known, it is clear that abnormalities in ceramide metabolism play a role in this disease entity. Several investigators have shown that PLD is associated with marked abnormalities in ceramide metabolism (53, 54). Ceramides are lipid molecules consisting of sphingosine and fatty acids. They are formed by hydrolysis of sphingomyelin by sphingomyelinases, but may also be synthesized through other pathways (55). It has been suggested that PLD results in increased levels of ceramides within cells and that this is part of the mechanism of drug-induced PLD (56). Other investigations support the hypothesis that ceramide metabolism plays a key role in PLD. Exposure of human macrophages to oxidized low-density lipoprotein results in both PLD and an increased content of acid sphingomyelinase and ceramides (57). The potential importance of the ceramide transacylase lysosomal phospholipase A2 (LPLA2) in alternate pathways of ceramide metabolism has been emphasized by Shayman et al. (53). Hiraoka et al. studied mice made deficient in LPLA2 by deletion of exon 5 of the (Lpla2) gene. These mice develop increases in ceramides and PLD of alveolar macrophages, peritoneal macrophages and spleen (58). Abe et al. studied the effects of three compounds: D-threo-1-phenyl-2-decanoylamino-3-morpholino-propanol (PDMP), a glucosylceramide synthase inhibitor and PLD-inducing agent; the strongly phospholipidotic agent, amiodarone; and tetracycline, which does not cause PLD, on Madin Darby canine kidney cells. Both PDMP and amiodarone inhibited LPLA2 and the transacylation of ceramide. This effect was concentration-dependent, and no effect was seen with tetracycline (59). Lecommandeur studied rat HEP G2 liver cells after chloroquine-induced PLD and found changes in the fatty acyl chain lengths of the ceramides, as well as changes in ceramide synthases, and suggested these changes might affect membrane morphology (60).
It is known that ceramides are important in the induction of multiple diseases, including atherosclerosis (61), diabetes (62) and cancer (63), as well as in growth arrest and cell apoptosis (64). It is also known that ceramides play a key role in TNFα and IL1 signal transduction (65-68). Studies have shown that IL1, IFNγ and TNFα can stimulate sphingomyelinase activity, resulting in ceramide accumulation (61, 69-72), and ceramide has frequently been referred to as the mimic of TNFα because of its similar effects on tumor-cell apoptosis and proliferation, and its role in many other disease processes (73-77). Lopez-Marure et al. showed that treatment with C8-ceramide had identical effects to those of TNFα on endothelial-cell proliferation and DNA synthesis (74). Similarly, Dbaibo et al. showed that TNFα and C2-ceramide had identical effects on the growth of Jurkat T-cells (76). Raines et al. showed that C2-ceramide and C6-ceramide mimicked the effects of TNFα on mitogen-activated protein kinase (MAPK) enzymatic activity, and on tyrosine phosphorylation (77). It is known that ceramides down-regulate the hERG K+ channel (78-80). Ceramides are metabolized to sphingosine and fatty acids, and sphingosine is phosphorylated by sphingosine kinases to form sphingosine-1 phosphate. The ceramide and sphingosine-1 pathways have opposite effects, the ceramide pathway causing cell death, and the sphingosine-1 phosphate pathway promoting cell survival (81-83). It is the balance of these two pathways that determines the fate of the cell (81). Since it has been postulated that ceramide metabolism is the common pathway to the development of atherosclerosis, perhaps through stimulation by TNFα (61), it is not unreasonable to ask whether the ceramide pathway might also be key to the mechanism of QT prolongation.
Agents that Reduce the Prolonged QT Interval
It has been long known that female sex is a risk factor for the long QT syndrome (84). However, QT intervals are shorter in the luteal phase of the menstrual cycle, when progesterone levels are higher, than in the follicular phase (85). This suggests that progestins have a protective effect against long QT (86). Kadish et al. studied 34,378 post-menopausal women who used unopposed estrogen, combined estrogen and progesterone, or used no therapy. A mild QT-prolonging effect was seen with estrogen, which was reversed with progesterone (87). Nakamura et al. investigated the effects of progesterone on guinea pig ventricular myocytes using patch clamp techniques. Progesterone shortened action potentials by lengthening IKs and inhibiting ICaL (88). Odening et al. studied ovariectomized transgenic LQT2 rabbits, which were exposed to estradiol, progesterone or dihydrotestosterone. In the estrogen-treated rabbits, ICaL increased, while in the progesterone-treated animals, ICaL decreased. Major cardiac ventricular events occurred in five out of seven estradiol-treated rabbits, but in none of the progesterone- or dihydrotestosterone-treated animals (89). Progesterone is known to have anti-inflammatory actions in many diseases. Aisenberg et al. showed that the anti-inflammatory effects of progesterone were essential in preventing pregnancy loss after LPS administration in mice (90). Loudon emphasized the importance of progesterone suppression of IL8 and cyclooxygenase-2 in maintaining myometrial quiescence in pregnancy and avoiding premature labor (91). Multiple studies have suggested that progesterone can reduce secondary damage from inflammation after traumatic brain injury by reducing levels of TNFα, IL1β and IL6 (92-94). Giannoni et al. showed that progesterone dramatically reduced secretion of TNFα and IL6 from mononuclear cells in umbilical cord blood from newborn children (95). Progesterone is known to induce sphingosine kinase, and the S1P pathway, the counteracting pathway to ceramides (96). Recent studies have suggested that non-genomic progesterone signaling may be a counteracting force to the effects of ceramides (97-103). Kupchak et al. have shown that non-genomic progesterone receptor activity can be antagonized by TNFα, and by 1(S),2(R)-D-erythro-2-(N-myristoylamino)-1-phenyl-1-propanol, a ceramidase inhibitor (101). Moussatche and Lyons have suggested that progesterone receptors act as the ‘fulcrum’ of the ceramide/S1P rheostat, determining whether ceramide synthase or ceramidase activity is greater, and that progestins and TNFα play opposing roles in the actions of these enzymes (98). Thomas and Pang suggested the neuroprotective effects of progestins are mediated by this mechanism (100). This suggests that the QT-shortening action of these agents may occur because of their opposing actions to TNFα and ceramides. It is also interesting that both estrogens and anti-estrogens, such as tamoxifen and toremifene, cause QT prolongation, and that both estrogens and anti-estrogens increase production of ceramides, although by different mechanisms. Tamoxifen is a potent inhibitor of glucosylceramide synthase and of acid ceramidase (104, 105). Estrogens increase the activity of ceramide synthases (106, 107).
3-Hydroxy-3-methylglutaryl coenzyme A reductase inhibitors (statins) may reduce ceramides by a number of mechanisms. Statins have also been shown to reduce levels of pro-inflammatory cytokines in many conditions, and discontinuation of these medications in patients being treated for hypercholesterolemia may result in a marked increase of cytokines and other inflammatory markers (108-110). In animal models of traumatic brain injury, the anti-cholesterol agents, atorvastatin, simvastatin and lovastatin, reduce brain and cerebrospinal fluid levels of IL1β, IL6 and TNFα (111-113). Iwata et al. showed that statins reduced IL6 and IL8 expression from LPS-stimulated human bronchial epithelial cells (114), and statin use is associated with decreased mortality in patients with pneumonia (115-117) and sepsis (118, 119). Thomsen et al., for example, reviewed the case histories of 29,900 patients hospitalized with pneumonia. Mortality among statin users was 10.3% at 30 days and 16.8% at 90 days, compared to 15.7% and 22.4% among non-statin users. These investigators also noted that statin users had lower blood levels of the inflammatory marker, C-reactive protein (116). Statins are known to reduce the QT interval (120, 121), and to reduce the chance of ventricular arrhythmia (122-125), and the reduction in ventricular arrhythmias has been linked to decreased inflammation (121, 124). Several investigators have argued that the primary underlying mechanism for the pleiotropic effects of statins is activation of sphingosine-1 phosphate-1 (S1P1) signaling (126, 127). Sugiura et al. showed that treatment with rosuvastatin led to an increase in the plasma level of S1P1 (128), and it has been reported that both atorvastatin and pitavastatin cause an up-regulation of S1P1 receptors (127). Furthermore, addition of pravastatin has been shown to prevent ceramide-induced death of mouse cerebral endothelial cells (129). Wei et al. showed that pravastatin and simuvastatin blocked oxidized LDL-induced acid sphingomyelinase activity and ceramide production in coronary artery endothelial cells (130).
Other agents have been shown to shorten the QT. We showed that liposomal curcumin, as well as empty liposomes, can attenuate the prolongation of the QT interval caused by crizotinib, nilotinib and other QT-prolonging agents (131-133). Liposomal curcumin and empty liposomes suppress TNFα, IL1β, IL6, monocyte chemoattractant protein 1 (MCP1), macrophage inflammatory protein 1-alpha (MIP1α) and Rantes, in both in vitro and in vivo models (134). Curcumin, which has both pro- and antioxidant effects, has been reported to protect against amiodarone pulmonary toxicity [a precursor to pulmonary PLD (135)] by reducing TNFα and TGFβ (136). Curcumin itself may cause prolonged QT because of its pro-oxidant effects, or because it can stimulate de novo ceramide synthesis (104), yet QT prolongation is blocked by liposomes. It has been suggested that liposomes, besides their anti-inflammatory activities, may compete with sphingomyelin for the enzyme sphingomyelinase, thus reducing the production of ceramides, or act by direct interaction with the cell membrane (137-139).
We have noted that in experimental models, the inducing effect of TNFα on QT prolongation can be blocked by antioxidants (41, 43), and other studies in animal models (140) and in patients (141, 142) have confirmed this. Zhang et al. showed that antioxidants restored IKr and hERG in the diabetic rabbit model of diabetes (140). Bednnarz et al. showed antioxidant vitamins corrected QT dispersion after exercise in patients post myocardial infarction (141). Kuklinski et al. reported that in patients treated with coenzyme Q and selenium, prolonged QT intervals were normalized in all, while 40% of controls continued to demonstrate prolonged QT. None of the treated patients died of re-infarction, while 20% of control patients had died of re-infarction by one year (142). Prolonged QT in diabetic patients has been associated with reduced plasma concentrations of vitamin C (143). Numerous studies have shown that ROS cause activation of sphingomyelinases (144-150). Vitamin E has been shown to reduce neutral sphingomyelinase activity and prevent neuronal death in rats after cerebral ischemia (151), although it may have contrary effects in cancer cells (152). Vitamin E has also been shown to prevent the induction of PLD in U937 cells exposed to 7-ketocholesterol (153). Navas et al. reported that coenzyme Q regulates the release of ceramides by non-competitive inhibition of neutral sphingomyelinase (154). The antioxidants 4,5-dihydroxy-1,3-benzenedisulfonic acid disodium salt monohydrate (Tiron) and N-acetylcysteine have been shown to block acid sphingomyelinase activity and the formation of ceramide-enriched membrane platforms that are stimulated by TNF-related apoptosis-inducing ligand (104, 155).
Cytokine-induced Prolongation of QT Likely Occurs Through ROS and Ceramides
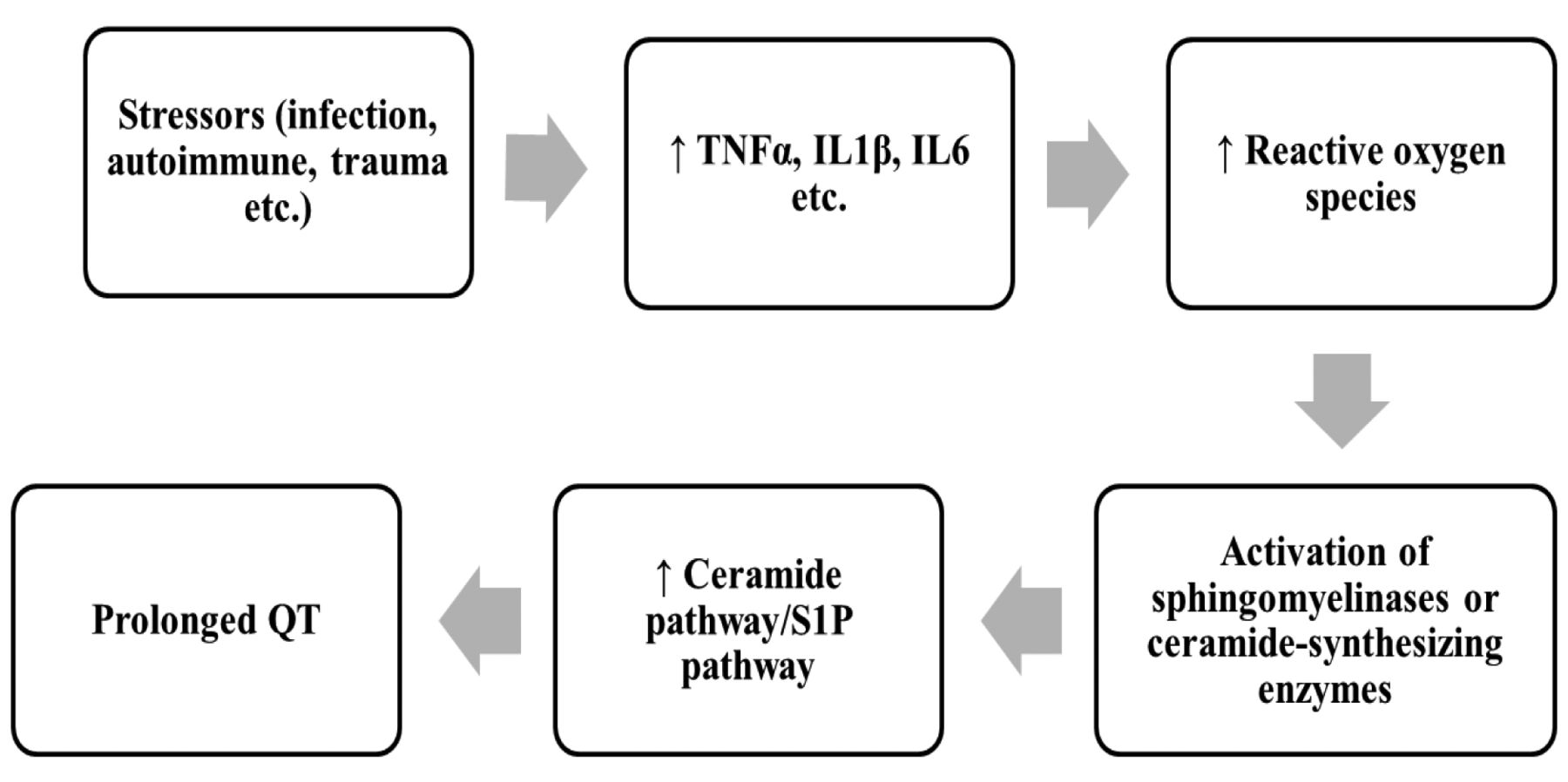
The information from these studies suggests the pathway by which cytokines cause prolonged QT. After stress, cytokines are released, and in turn, stimulate production of ROS. ROS cause activation of sphingomyelinases, which stimulate the ceramide pathway, and alter the ceramide pathway/sphingosine-1-phosphate pathway balance (Figure 1). The first step, concerning the role of stressors in the induction of pro-inflammatory cytokines, is well documented. As noted above, Wang et al. (41) and Fernandez Velasco et al. (43) both found that TNFα caused IKr, IKs and Ito suppression in experimental models, and that this effect was reversible by antioxidants such as vitamin E, and by a superoxide dismutase mimic, suggesting that TNFα caused this suppression by stimulating ROS. Pro-inflammatory cytokines (TNFα, IL1β, IL6, and IFNγ) have been shown to stimulate ROS in other experimental models (156-159). As noted, studies have shown that ROS can cause the activation of sphingomyelinases, causing an increase of ceramide levels (144-150). The mechanism of histone deacetylase inhibitor-induced leukemia cell death, for example, is thought to be secondary to acid sphingomyelinase-dependent generation of ceramides. This process does not occur in the absence of ROS stimulation of sphingomyelinase, and is blocked by antioxidants (146). ROS can also stimulate the salvage and de novo pathways of ceramide synthesis (160, 161), and this is likely the mechanism of TNFα stimulus of the de novo pathway (162). An increase in ROS appears to be the major mechanism of the cytokine effect on these enzymes, particularly on sphingomyelinases, although ROS-independent pathways may also play a role (163). ROS also inhibit sphingosine kinase-1, and thus further tilt the balance between the ceramide pathway (destructive) and the S1P pathway (protective) (81, 164, 165). The ceramide pathway is known to regulate numerous potassium channels, including hERG (78-80, 166-169). Ganapathi et al. studied the effects of C6-ceramide on HEK-293 cells and found that ceramide inhibited hERG channel current, and recruited hERG channels within caveolin-enriched lipid rafts (169). Chapman et al. reported an up to 30% decrease in hERG current with C6-ceramide, which they attributed to a reduction in the total number of hERG channels (78). Bai et al. found that C2-ceramide reduced hERG current, an effect they concluded was moderated by ROS (80). Wu et al. showed that C2-ceramide inhibited IKr in neuroblastoma IMR-32 cells and in rat pituitary GH(3) cells, and that this effect could be duplicated by TNFα (79).

A simplified model: Cytokine causation of the prolonged QT syndrome.
Drug-induced Prolonged QT
We have argued that pro-inflammatory cytokines cause prolonged QT by stimulating the ceramide pathway. If this hypothesis is accurate, we must account for the mechanism of action of QT-prolonging drugs, many of which are actively associated with the suppression of cytokines. We believe that although elevated levels of TNFα, IL1β and IL6 cause QT prolongation through the ceramide pathway and that this is the explanation for the high incidence of prolonged QT in inflammatory conditions, agents inducing QT prolongation may bypass the initial steps in this model and cause prolonged QT by directly affecting the ceramide pathway and disrupting the ceramide pathway/S1P pathway balance.
An important class of agents that cause QT prolongation is the tyrosine kinase inhibitors (TKIs), which are now used extensively for the treatment of chronic myelogenous leukemia (CML), renal cell carcinoma, hepatoma, gastrointestinal stromal tumors and many other types of cancer. One of the most prominent toxicities of nilotinib, a second-generation TKI which targets the breakpoint cluster region–ABL proto-oncogene 1 (BCR–ABL) receptor on CML cells, is QT prolongation. It has been shown that nilotinib induces apoptosis of CML cells by up-regulating ceramide synthase and down-regulating sphingosine kinase (170, 171). Addition of exogenous ceramides increased the apoptotic effect of nilotinib. In another study, El-Agamy showed that nilotinib caused a decrease in inflammatory cytokines and ROS generation and ameliorated LPS-induced acute lung injury in rats, presumably secondary to its stimulatory effect on ceramides and subsequent reduction in TNFα (172). Similarly, dasatinib, another second-generation TKI which is similar to nilotinib and which also causes QT prolongation, also causes apoptosis of CML cells by up-regulating ceramide synthase and down-regulating sphingosine kinase, thus shifting the ceramide/S1P balance toward ceramide (173). Likewise, alterations in the ceramide/S1P rheostat toward excess S1P have been associated with resistance to imatinib in patients with CML. The efficacy of imatinib has been linked to its generation of ceramides, particularly C18-ceramide, whereas resistance is thought to be caused by increased sphingosine kinase-1 activity and increased levels of S1P (174). Resistance has also been correlated with increased activity of glucosylceramide synthase, which converts ceramide to glucosylceramide. PDMP, a glucosylceramide synthase inhibitor and inducer of PLD, increased CML cell kill after treatment with imatinib (175). Huang et al. showed PDMP sensitizes resistant T315I mutant CML cells to a BCR–ABL inhibitor (176). Gao et al. reported that resistance to sunitinib, a tyrosine kinase inhibitor which causes prolonged QT and which is commonly used for treatment of renal carcinoma, gastrointestinal stromal tumors and pancreatic neuroendocrine tumors, is caused by activation of sphingosine kinase-1 and stimulation of the S1P pathway (177). Salas et al. showed that sphingosine kinase-1 and S1P mediate BCR–ABL stability and cause resistance to therapy, and that the efficacy of nilotinib depended on suppression of S1P, consistent with the view that all these agents work by tilting the ceramide/S1P balance toward ceramide (178). Many other investigators have found similar results (179-181).
While it is well-known that pro-inflammatory cytokines can stimulate the production of ceramides, it is also well-documented that exogenous ceramides, and the stimulation of the ceramide pathway, can cause a feedback inhibition of cytokines (182-188). For example, Jozefowski et al. showed that treatment with C8-ceramide reduced production of TNFα and MIP2 in murine peritoneal macrophages and in J774A.1 macrophage-like cells after LPS stimulation, and concluded that ceramide acted as a negative regulator of cytokine production. Inhibition of sphingomyelinase increased cytokine production (182). Rozenova et al. found similar results. They studied acid sphingomyelinase-deficient mice and peritoneal macrophages derived from the animals, and found that both the animals and the isolated macrophages produced 10- to 15-fold higher levels of TNFα when stimulated with LPS (183). Walton et al. reported that C6-ceramide inhibited LPS-induced IL8 synthesis in human aortic endothelial cells (184). The negative regulation of cytokines by ceramides may explain why medications which bypass the initial steps in the inflammatory pathway to ceramides and cause prolonged QT directly, may also cause cytokine suppression (189).
Other agents that cause QT prolongation are known to stimulate aberrant ceramide metabolism. For example, chlorpromazine, a commonly used antipsychotic drug, has been shown to reduce the level of acid ceramidase, which degrades ceramides to sphingosine and fatty acids, causing increased levels of ceramides (190-192). It is also well-known that chlorpromazine can induce inflammatory conditions, such as drug-induced lupus (193, 194). Hieronymus et al. have shown that the addition of S1P, which shifts the ceramide/S1P balance away from ceramide, can block chlorpromazine-induced apoptosis of peripheral blood mononuclear cells, and have suggested that aberrations of the ceramide pathway are the cause of drug-induced lupus (195).
Tricyclic antidepressants have been shown to have effects on ceramide/S1P balance. Desipramine inhibits acid ceramidase (190-192), although it also has an inhibitory effect on sphingomyelinase (196-198). Because of their inhibitory effects on sphingomyelinase, tricyclic antidepressants have been studied extensively as possible suppressors of ceramides, and it has been speculated that the antidepressant effects of these agents may be due to their effects on this enzyme. However, Elojeimy et al. treated DU145 prostate carcinoma cells with desipramine and found an increase in ceramide levels, suggesting that in some cases, the effects of desipramine on acid ceramidase may outweigh its effects on sphingomyelinase (192). That sphingomyelin hydrolysis and ceramidase activity may produce ceramides with different fatty acid chain lengths, which is discussed below, may be another explanation why tricyclic antidepressants cause prolonged QT despite inhibiting sphingomyelinase.
Antimalarials, such as mefloquine and chloroquine, have profound effects on the ceramide pathway. Mefloquine inhibits sphingomyelin synthase and glucosylceramide synthase, leading to an increased intracellular ceramide concentration, and this is thought to be its mechanism of action against Plasmodium falciparum (199). Mefloquine also increases sphingomyelinase (200) and reduces sphingosine kinase (201). Chloroquine inhibits acid ceramidase (190).
Cyclooxygenase-2 inhibitors, such as celecoxib, are known to cause prolonged QT (202), and to increase ceramide levels by activating sphingomyelinase (203-205). In one study, addition of C6-ceramide duplicated the effects of celecoxib on the cell cycle and cell growth inhibition (204). Schiffmann et al. showed that the toxic effects of celecoxib occur by activation of ceramide synthase 6 and the salvage pathway (206).
Fingolimod, a sphingosine analog with both agonist and antagonist effects on the S1P1 and other S1P receptors, and which sequesters lymphocytes in lymph nodes, is used to treat patients with relapsing multiple sclerosis. Fingolimod causes QT prolongation and inhibition of the hERG current (207-209), and sometimes fatal arrhythmia (210). This drug also can cause profound suppression of cytokines, including TNFα, IL1β, MCP1, interferon γ, chemokine (C-X-C motif) ligand 1 and TGFβ (211, 212).
In the mouse model of influenza-induced cytokine storm, modulation of the S1P1 receptor was identified as the primary pathway for initiation of the storm (213). The storm was reversed in this model through feedback inhibition by a S1P1 receptor modulator, with marked reductions in TNFα, MCP1, IFNγ, MIP1α, IL6 and other cytokines (214, 215). In this study, the S1P1 receptor modulator was a much more effective therapy than the antiviral agent, Tamiflu: 82% of mice treated with the receptor modulator survived compared to 50% with Tamiflu and to 20% of controls, 96% (27/28) of mice treated with both the receptor modulator and Tamiflu survived (213). Ceramides are composed of sphingosine and fatty acids. Sphingosine kinase, the enzyme which catalyzes sphingosine to S1P, is the crucial enzyme in the balance between the opposing ceramide to S1P pathways, and sphingosine has important ceramide-like effects (216). In addition, fingolimod, like tricyclic antidepressants, appears to inhibit acid ceramidase (217). Other sphingosine analogs, such as siponimod, have also been reported to cause prolonged QT (207).
Many other agents that are known to cause prolonged QT have been shown to affect the ceramide pathway and disrupt the ceramide/S1P balance, including aminoglycosides (218-221), antiestrogens such as tamoxifen (222), quinolones (221-224), monoclonal antibodies such as rituximab (225), platinum-containing antineoplastics (226-228), calcium channel blockers (229, 230), anthracyclines (231, 232), arsenic trioxide (233, 234), antihistamines (229), taxanes (235), cyclosporines (235) and imidazole antifungals such as ketoconazole (235). A list of QT-prolonging drugs and their effects on ceramide signaling is given in Table I.
Ceramide Fatty Acid Chain Length
The production of specific ceramides may be as important as the total amount of cytokines in the causation of the prolonged QT interval (236-238). Senkal et al. showed that treatment with the anthracycline doxorubin, in combination with gemcitabine, against human head and neck carcinoma cells, resulted in a 3.5-fold increase in ceramide synthase activity. When these cells were heterotransplanted into mice, a 7-fold increase in C-18 ceramide was seen after treatment, despite no increase in total ceramide levels (236). Veret et al. showed that treatment of pancreatic β-cells in culture with high doses of glucose resulted in apoptosis through stimulation of ceramide synthase-4, and increases in the levels of C18:0, C22:0 and C24:1 ceramides, without an increase in total ceramides. Down-regulation of ceramide synthase-4 by short-interfering RNA reduced apoptosis (238). C16:0 ceramide appears to have an important role in the induction of glucose intolerance. Turpin et al. reported that obese patients had elevated levels of ceramide synthase-6 mRNA and C16:0 ceramide in their adipose tissue. In addition, they showed that ceramide synthase-6-deficient mice had reduced levels of C16:0 ceramide and did not develop glucose intolerance or obesity after a high-fat diet (239). In patients with Parkinson's disease, ceramide levels are generally elevated. However, levels of C16:0, C18:0, C20:0, C22:0 and C24:1 ceramides are especially high, particularly in those who have significant cognitive impairment (240). There is also a great deal of data showing that elevated levels of C18:0 and C24:0 ceramides contribute to disease severity in patients with Alzheimer's disease (241). Such patients have greatly increased levels of C24:0 ceramide in the middle frontal gyrus, and the levels of C18:0 and C24:0 ceramides there correlate with disease severity. Addition of amyloid β-peptide-42 to cultured hippocampal neurons has been shown to result in increased C18:0 and C24:0 ceramide levels, and this effect can be reversed by an inhibitor of serine palmitoyltransferase (242). Numerous investigators have shown that C16:0, in particular, is crucial for apoptosis, both of malignant and normal cells (241-245). In cancer, and other diseases, it has been suggested that very long chain ceramides interfere with the toxic effects of C16:0 ceramide, and that what is crucial in many diseases is the balance between C16:0 and C24:0 ceramides, with C16:0 having toxic and C24:0 having protective effects (241, 242, 247).
The role of ceramides with specific fatty acid lengths in the pathogenesis of the prolonged QT syndrome and other abnormalities of cardiac function is not known. However, as noted, the production of different cytokines by different enzymes in the various ceramide pathways could be part of the reason why tricyclic antidepressants cause prolonged QT despite their tendency to inhibit sphingomyelinase. Di Paola et al. showed that C2-ceramide stimulated, while C16-ceramide inhibited, cytochrome c oxidase activity in rat heart mitochondria. Short-chain ceramides inhibited the generation of ROS associated with membrane potential-dependent reverse electron flow from succinate to complex 1, while long-chain ceramides had no effect. The investigators in this study concluded that short-chain ceramides caused a collapse in membrane potential, leading to an increase in permeability (248). Monette et al. reported that the impaired electron transport and increase in ROS in the aging rat heart after chronic inflammation was caused by increased levels of C16:0, C18:0 and C24:1 ceramides. The increased ceramide levels and mitochondrial abnormalities were reversed by (R)-α-lipoic acid (249). Russo et al. reported that mice which were fed a diet high in myristate (C14:0) developed marked increases in ceramide synthase-5 and C14-ceramide through the de novo pathway, resulting in cardiac hypertrophy and dysfunction (250). No effect was seen with a high palmitate (C16:0) diet.
QT-Prolonging agents and their effects on ceramide/S1P signaling
The significance of these findings is unclear, but there are hints that C16:0 ceramide may be especially important in the causation of QT prolongation, just as it is in other diseases. In a series of elegant experiments, Schiffmann et al. reported that celecoxib activation of ceramide synthase-6 induced C16:0 ceramide selectively, and showed that the toxic effects of celecoxib on cell growth were solely due to C16:0 ceramide production (206). As noted, Lecommandeur reported that drug-induced PLD is characterized by an increase in C16:0 ceramide, even when the total ceramide content is not changed (60). Bock et al. showed that treatment of Jurkat T-lymphocytes with C16:0 ceramide causes clustering of the voltage-gated potassium channels Kv1.3 within ceramide-enriched membranes, resulting in inhibition of Kv1.3 activity (251). It is also known that many of the toxic effects of TNFα occur specifically through generation of C16:0 ceramide, and that these toxic effects can be blocked by reduction of this ceramide (252, 253). Likewise, it has been shown that the toxic effects of IFNγ in models of autoimmune disease occur specifically through ceramide synthase-6 and C16:0 ceramides, and that these effects are reversed by inhibition of this ceramide (254).
Conclusion
We conclude that excess levels of pro-inflammatory cytokines play a role in causing prolongation of the QT interval, and that the likely mechanism of action is through ROS and the ceramide pathway. We also believe that QT-prolonging drugs may bypass the initial steps and cause prolonged QT by direct effects on the ceramide and S1P pathways and alterations of the ceramide/S1P rheostat. Elevated levels of cytokines may predispose a patient to QT prolongation, and this may be the reason that the QT-prolonging potential of these agents may not be observed in initial studies in human volunteers. Recent studies have suggested that it may be the stimulation of ceramides with particular fatty acid chain lengths, as much as the increase in total ceramides, which is important in the pathogenesis of some diseases. The potential role of individual ceramides, particularly C16:0 ceramide, in the causation of QT prolongation needs to be investigated. That pro-inflammatory cytokines and QT-prolonging medications may cause QT prolongation by the same mechanism means extra caution is needed when using these agents in patients with inflammatory conditions.
Footnotes
This article is freely accessible online.
Conflicts of Interest
The Authors acknowledge a potential conflict of interest. Dr. Peter Sordillo is a member of the Scientific Advisory Board of SignPath Pharma, which makes several formulations of intravenous curcumin. Dr. Helson is CEO of SignPath Pharma.
- Received July 28, 2015.
- Revision received September 9, 2015.
- Accepted September 25, 2015.
- Copyright © 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
Jump to section
- Article
- Abstract
- Data from Patient Trials
- Cytokines: Data from Experimental Models
- Phospholipidosis and Ceramides
- Agents that Reduce the Prolonged QT Interval
- Cytokine-induced Prolongation of QT Likely Occurs Through ROS and Ceramides
- Drug-induced Prolonged QT
- Ceramide Fatty Acid Chain Length
- Conclusion
- Footnotes
- References
- Figures & Data
- Info & Metrics
Related Articles
Cited By...
- Pirarubicin, an Anthracycline Anticancer Agent, Induces Apoptosis Through Generation of Hydrogen Peroxide
- The Relationship Between Cardiac Conduction Times, Cardiovascular Risk Factors, and Inflammation in Patients with Early Arthritis
- In Utero Particulate Matter Exposure Produces Heart Failure, Electrical Remodeling, and Epigenetic Changes at Adulthood
- Sphingosine Kinase Inhibitors as Maintenance Therapy of Glioblastoma After Ceramide-Induced Response