


Abstract
Connective tissue growth factor (CCN2) regulates diverse cellular functions, including tooth development. In order to delineate the precise role of CCN2 in the epithelium during odontogenesis, we investigated how it is expressed and what roles it may have in primary cultures of epithelial cells derived from developing tooth germ of the bovine fetus. Ccn2 mRNA and protein were strongly expressed in the inner dental epithelium, which is consistent with the expression of transforming growth factor-β2 mRNA and proliferating cell nuclear antigen. Bone morphogenetic protein 4 (BMP4) and fibroblast growth factor 2 (FGF2) were also expressed in the inner dental epithelium, indicating that CCN2 functionally interacts with these factors in the epithelium. The stimulatory effects of FGF2 on cell proliferation and BMP4 on cell differentiation were additively up-regulated by CCN2 in a newly-established dental epithelium cell culture. Taken together, our data provide clear evidence that CCN2 is synthesized by inner dental epithelial cells, and appears to act as an autocrine factor, which regulates dental epithelial cell proliferation and differentiation in concert with growth factors.
The development of teeth is a multistage and multistep process involving proliferation and differentiation (1). Odontogenesis begins with the formation of the dental lamina, a localized thickening of the oral epithelium. The dental lamina then invaginates into the underlying mesenchyme to form tooth buds. The epithelial cells of each tooth bud then proliferate further to form a dental organ. The epithelial cell layer of the dental organ in close contact with the dental mesenchyme is known as the inner dental epithelium, which differentiates into ameloblasts (2).
Connective tissue growth factor (CCN2) is a heparin-binding multi-functional growth factor that regulates the development of organs by directly inducing cells to enter the cell cycle and to secrete a variety of cytokines, growth factors and extracellular matrix proteins (3, 4). It is a member of the CCN family of proteins, which is characterized by four distinct domains which contain the functional region for integrin recognition and proteoglycans (3, 4). We have reported that CCN2 is expressed during odontogenesis, and appears to be needed for proliferation and differentiation along both ameloblasts and odontoblast cell lineages in a mouse in vivo model (5). In the present study, we examined the expression of CCN2 and its effect on dental epithelial cells in a primary culture system obtained from developing bovine fetus tooth germs, a system that has the advantage of closely mimicking the physiological state of cells in vivo.
Materials and Methods
Immunohistochemistry. Mandibular or maxillary fragments containing tooth germs isolated from bovine embryos were fixed overnight in buffered 4% paraformaldehyde and embedded in paraffin. Continuous 5-mm-thick sections were prepared and treated with phosphate buffered saline (PBS) containing 2 mg/ml glycine for 10 min. After blocking with 10% normal goat serum in PBS for 1 h, the sections were reacted with different dilutions of antibodies to CCN2 (5) and proliferating cell nuclear antigen (PCNA, Boehringer-Mannheim, Mannheim, Germany). As a control, companion sections were incubated with preimmune rabbit IgGs or with blocking solution alone. After rinsing with 3% normal goat serum in PBS, the sections were incubated with a 1:250 dilution of biotin-conjugated goat anti-rabbit IgG (Boehringer-Mannheim) in 10% normal goat serum in PBS for 2 h, rinsed again, incubated with 1:500 dilution of streptavidin β-galactosidase conjugate (Boehringer-Mannheim) for 1 h, rinsed, and slides were analyzed with Nikon microscope (Nikon Inc., Melville NY, USA)

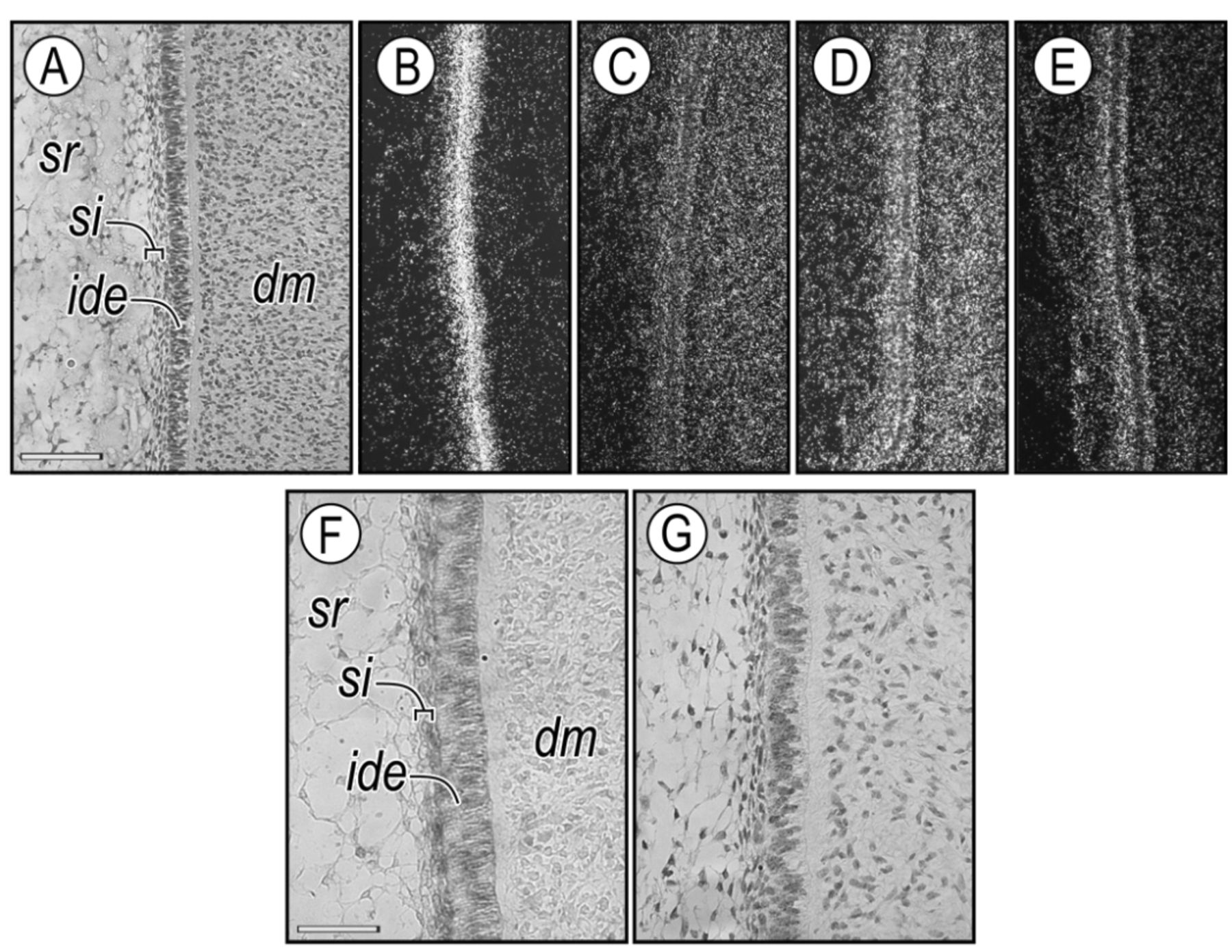
In situ hybridization and immunohistochemical analysis of bovine fetus bell-stage molar tooth germs. Tissue sections from tooth germs in the bell stage were hybridized with radiolabeled antisense Ccn2 (B), Bmp4 (C), Tgfβ2 (D) and Fgf2 (E). Panel A shows a high-magnification image of a tooth germ with a characteristic inner dental epithelium (ide), dental mesenchyme (dm), stratum intermedium (si), and stellate reticulum (sr). Companion serial sections of bell-stage tissue were processed for immunohistochemical localization of CCN2 protein (F) and PCNA (G). Scale bars, A: 100 μm (also applies to B-E); F, G: 40 μm.
In situ hybridization. For in situ hybridization, paraffin-embedded serial tissue sections of bovine fetus tooth germs were pre-treated with 10 mg/ml proteinase K (Sigma, St. Louis, MO, USA) for 10 min at RT, postfixed in 4% paraformaldehyde, washed with PBS containing 2 mg/ml glycine, and treated with 0.25% acetic anhydride in triethanolamine buffer. The sections were then hybridized with antisense or sense 35S-labeled probes (approximately 1×106 disintegrations (DPM)/section) at 50°C for 16 h. The probes have been described previously (6, 7). After hybridization, the slides were washed with 2 standard saline citrate (SSC) containing 50% formamide at 50°C, treated with 20 mg/ml RNase A for 30 min at 37°C, and washed three times with 0.1×SSC at 50°C for 10 min/wash. The sections were dehydrated with 70%, 90%, or 100% ethanol for 5 min/step, coated with Kodak NTB-3 emulsion (Estman Kodak Company, Rochester, NY, USA) diluted 1:1 with water, and exposed for 10 to 14 days. The slides were developed with Kodak D-19 at 20°C and stained with hematoxylin, and analyzed and photographed with Nikon microscope (Nikon Inc.) equipped by for dark- and bright-field optics.
Cell separation and cell culture of odontogenic epithelial cells. Molar tooth germs from 3-month-old embryos were obtained from a heifer from a local slaughterhouse. The stages of bovine embryogenesis are usually described in terms of the longitudinal length of the fetuses. The embryos used here were about 15.0 cm in length (corresponding to approximately day 90 of gestation). Jaw fragments were placed into Hank's balanced salt solution (GIBCO BRL, Grand Island, NY, USA) containing antibiotics (50 U/ml penicillin, 50 mg/ml streptomycin, and 0.5% fungizone). Early bell-stage tooth germs isolated from day-90 bovine embryos were dissected free of surrounding tissues, and were briefly exposed to 1.2 U/ml dispase grade II (Boehringer-Mannheim), and then the mesenchymal cells were microsurgically separated from the epithelial cell layers. The epithelial tissue contained in the inner and outer enamel epithelium, stratum intermedium and stellate reticulum, which comprised a heterogeneous population of cells, was minced and incubated in a fresh mixture of 0.25% trypsin (Sigma, St. Louis, MO, USA) and 0.1% collagenase1 (Sigma) for 3 h, at which point the tissue was completely digested. The freshly isolated epithelial cells were plated at a density of 2×104/well in the wells of a 96-well micro-well plate or in culture dishes at 1×106 cells/60-mm dish in Dulbecco's modified Eagle's medium (DMEM, GIBCO BRL) containing 15% fetal bovine serum (FBS, GIBCO BRL) and incubated at 37°C in an atmosphere of 5% CO2/air. Recombinant human CCN2 (rCCN2) was purified by using the baculovirus expression system as described previously (5). Recombinant human FGF2, TGFβ and BMP4 were purchased from R&D Systems (Minneapolis, MN, USA) for in vitro experiments for the in vitro experiments.
Northern blot analysis. Total RNA was prepared from quiescent monolayers of odontogenic epithelial cells isolated from bovine tooth germs by using TRIZOL reagent (Life Technologies Inc. Rockville, MD, USA). Northern blot analysis was performed by the standard procedure. Briefly, 10 mg of total RNA was denatured by glyoxalation, size-fractionated by gel electrophoresis in 1% agarose gels at 10 μg/lane, and transferred to a membrane by capillary blotting, as described previously (8). The blots were stained with 0.04% methylene blue to verify that each sample had been transferred efficiently. Next, the blots were hybridized for 16 h to 32P-labeled DNA probes at of 2.5×106 cpm/ml of hybridization solution containing 50% formamide, 6×SSC, 1% sodium dodecyl sulfate (SDS, GIBCO BRL), 200 mg/ml sheared denatured salmon sperm DNA, and 10×Denhardt's reagent. The hybridization temperature was 42°C. After hybridization, the blots were rinsed several times at room temperature with 2×SSC and 0.5% SDS; and a final high-stringency rinse was done with 0.1×SSC and 0.5% SDS at room temperature. Finally, the blots were exposed to X-ray films (Estman Kodak Company) at −70°C.

Characteristics of bovine fetus dental epithelial cells in primary culture. A: Morphology of the primary dental epithelial cells in culture. The cells were cultured in Dulbecco's modified Eagle's medium (DMEM) containing 15% fetal bovine serum (FBS) with or without 5 μg/ml ascorbic acid or 1 mM β-glycerophosphate (β-GP) for three days (left panel). Aliquots of culture medium from [35S] methionine-labeled odontogenic epithelial cells with or without ascorbic acid or β-GP were analyzed by 8-16% Tris-glycine sodium dodecyl sulphate-polyacrylamide gel electrophoresis under reducing conditions (right panel). The arrows indicate procollagen bands. B: Histochemical detection of alkaline phosphatase (ALP) activity and alizarin-red staining in the epithelial cells after nine days of culture. C and D: Quantitation of ALP activity (C) and Ca accumulation (D) in epithelial cells.
Alkaline phosphatase (ALP) activity. For estimation of ALP activity, bovine dental epithelial cells were plated at a density of 1×105 cells/well in 24-well multiplates with DMEM containing 15% FBS and cultured for three days. The medium was replaced every three days with DMEM containing 15% FBS. After the incubation, the cells were rinsed with ice-cold PBS, homogenized in 0.5 M Tris (pH 9.0) containing 0.9% NaCl and 0.1% Triton X-100 on ice and centrifuged at 12,000 g for 15 min. ALPase activity in the resultant supernatants was determined by the method of Bessey et al. (9) with some modifications. The enzyme reaction was initiated with 0.5 mM p-nitrophenyl phosphate (p-NPP) in 0.5 M Tris-HCl (pH 9.0) containing 0.5 mM MgCl2 at 37°C and terminated by the addition of a quarter volume of 1 M NaOH. The concentration of p-NP generated was determined by spectrophotometry at 410 nm. The enzyme activity is expressed as nanomoles per p-NPP cleaved per minute per well.
Labeling of cell cultures. Bovine dental epithelial cells in culture dishes were washed with PBS, incubated at 37°C for 2 h in methionine-free medium (GIBCO BRL), and then pulse-labeled for 2 h with [35S] methionine (100 μCi/ml; NEN Life Science Products, Inc., Boston, MA, USA) with or without 5 mg/ml ascorbic acid (Sigma) or β-glycerophosphate (β-GP, Sigma). Following the pulse period, the cells were washed twice with complete DMEM containing 1% FBS and incubated for 2 h. The media from the 2-h chase period were collected. Finally, protease inhibitors were added, and the media were dialyzed against three changes of water, the aliquots of culture medium were analyzed by 8-16% Tris-glycine SDS-polyacrylamide gel electrophoresis under reducing conditions.
Cell proliferation assay. Bovine dental epithelial cells proliferation was determined with a proliferation enzyme-linked immunosorbent assay BrdU kit according to the manufacturer's protocol (Boehringer-Mannheim). The experiments were repeated at least three times, and triplicate samples were assayed.
Statistical analysis. Data were analyzed by using the unpaired Student's t-test for the analysis of 2 groups or one-way ANOVA for the analysis of repeated multiple group comparisons, and repeated measures ANOVA for the analysis of repeated multiple group comparisons. The results were expressed as the mean±S.D. and values of *p<0.05 and **p<0.01 were considered statistically significant.
Results
CCN2 expression patterns in developing bovine tooth germs. In bell-stage tooth germs from the bovine fetus, intense Ccn2 gene expression was detected in the inner dental epithelium and stratum intermedium (Figure 1A and B), and Bmp4 (Figure 1C), Tgfβ2 (Figure 1D) and Fgf2 (Figure 1E) mRNA were highly expressed in the inner dental epithelium and stratum intermedium. Bmp4 and Tgfβ2 mRNA were also detected in the dental mesenchyme cells (Figure 1C and D). CCN2 protein was expressed in the inner dental epithelium and stratum intermedium (Figure 1F), which were composed of proliferating PCNA-positive cells (Figure 1G).
Characterization of primary bovine dental epithelial cells in culture. To study the effect of CCN2 on the dental epithelial cells in vitro, we first established a conventional cell culture system using dental epithelial cells from bovine tooth germs. Dental epithelial cells were isolated from bell-stage tooth germs, and analyzed for their ability to further differentiate in an in vitro setting. When the cells were cultured with 5 μg/ml ascorbic acid, abundant bundles of extracellular matrix (ECM) appeared after three days of culture (Figure 2A, left panel), which was reflected by the relative intensities of radioactive procollagen bands separated on 8-16% Tris-glycine gels (Figure 2A, right panel, arrow). Ascorbic acid increased ALP staining (Figure 2B) and ALP activity for nine days of culture (Figure 1C). The combination treatment of 5 μg/ml ascorbic acid and 1 mM β-GP yielded only weak ECM formation in cells (Figure 2A left panel), procollagen expression for three days of culture (Figure 2A right panel), and low ALP activity (Figure 2B and C). On the other hand, ascorbic acid and β-GP treatment increased the deposit of calcium in the ECM in a time-dependent manner (Figure 2D). To examine the mineral composition in greater detail, mineral samples from ascorbic acid and β-GP cultures were processed for Fourier Transform Infrared (FT-IR) spectroscopy and determined to be biological hydroxyapatite (data not shown). Interestingly, control cells cultured in 15% FBS exhibited accumulation of calcium after 15 and 21 days of culture (Figure 2D).
Expression and role of CCN2 in dental epithelial primary culture. Firstly, we determined whether these dental epithelial cells expressed CCN2 in the proliferative phase, which occurs when the cells are cultured in DMEM containing 15% FBS. The proliferative dental epithelial cells expressed Ccn2 mRNA at a 5-fold higher level than the confluent stationary cells (Figure 3A). The Tgfβ mRNA expression level in these proliferating cells was slightly higher than that in the stationary cells (Figure 3A). On the other hand, the mRNA levels of Alp and type I collagen were not changed between the proliferating and stationary phase (Figure 3A). Because the tissue reconstitution experiments showed that CCN2 expression in the dental epithelium required an interaction with the dental mesenchyme but was maintained by treatment of the epithelium with TGFβ (5), we asked whether the TGFβ signal could regulate Ccn2 gene expression in a primary dental epithelial cell culture system. To answer this question, we treated dental epithelial cells with 10 ng/ml TGFβ at the indicated times, and found that TGFβ achieved a significantly greater stimulation of the Ccn2 mRNA expression at 6 h and 24 h of culture compared to the level in vehicle-treated control cells (Figure 3B). We then asked whether CCN2 would affect the proliferation and differentiation of dental epithelial cells in a primary culture system. Primary dental epithelial cells were treated with various amounts of CCN2 with or without 10 ng/ml FGF2 for 24 h, and pulse-labeled with BrdU during the last 2 h of culture. CCN2 did not have an effect on the proliferation and ALP activity when administered alone, but did significantly enhance the proliferation of the epithelial cells when added together with 10 ng/ml FGF2 (Figure 3C), and significantly enhanced the differentiation of the cells when added together with 500 ng/ml BMP4 (Figure 3D).

Northern blot analysis of the proliferative phase of developing dental epithelial cells in primary culture. A: Bovine dental epithelial cells were inoculated at a density of 1×106/dish into 6-cm dishes and cultured in Dulbecco's modified Eagle's medium (DMEM) containing 15% fetal bovine serum (FBS) for 48 h. The medium was changed to 3 ml of DMEM containing 15% FBS, and the cells were cultured for 12 h, after which time they were actively proliferating (proliferative phase). After 36 h, the medium was replaced with 3 ml of DMEM containing 15% FBS, and the cells were cultured for 12 h until they reached confluence (stationary phase). B: Proliferating cells and those in the stationary phase were examined by Northern blotting for their expression of Ccn2, transforming growth factor-β (Tgf-β), alkaline phosphatase (Alp), and type I collagen (Coll-1). The gene expression was analyzed by computer-assisted densitometric analysis of band intensities. Induction of Ccn2 mRNA expression by treatment with TGFβ was also examined (right panel). C: The effect of recombinant CCN2 on the proliferation of dental epithelial cells treated with or without 100 ng/ml FGF2. D: The effect of CCN2 on 500 ng/ml BMP4-induced ALP activity in bovine dental epithelial cells. *Statistical significance defined as p<0.05.
Discussion
In this study, we demonstrated that CCN2 is strongly detected in PCNA-positive bovine dental epithelial cells in the early bell stage. Our in vitro data suggest that CCN2 positively regulated the proliferation of dental epithelial cells that were isolated from bovine developing tooth germs and placed in primary culture. We found that intense Ccn2 mRNA expression was well-correlated with the proliferating phase of the dental epithelial cells and that once the cultures had become confluent, Ccn2 expression decreased. These in vitro data are consistent with our in vivo data (Figure 1B and G) and our previous in vivo study showing that E14 mouse tooth germs injected with CCN2-neutralizing antibodies were smaller in overall size than those in controls and that cell proliferation was severely reduced (5). We also showed that CCN2 positively regulated the proliferation of dental epithelial cells in the presence of FGF2. FGF2 signaling is known to be involved in tooth development (10, 11). CCN2 can activate intracellular FGF2 signaling, resulting in the stimulation of cells and matrix metalloproteinase production by chondrocytes (12). These findings suggest that CCN2 may regulate the proliferation and matrix degradation of dental epithelial cells by forming a complex with FGF2 as a novel modulator of FGF2 functions. By utilizing this conventional dental epithelial cell culture system, we also showed that CCN2 induces ALP activity by co-treatment with BMP4. The data suggest that CCN2 may regulate the differentiation of ameloblasts in the presence of BMP signaling in vivo. It has been reported that CCN2 can antagonize BMP4 activity by preventing its binding to BMP receptors in Xenopus embryos (13). On the other hand, CCN2 directly interacted with BMP2, and CCN2–BMP2 stimulated proteoglycan synthesis via the suppression of extracellular signal-regulated kinase1/2 (ERK1/2) phosphorylation in cultured chondrocytes (14). These findings suggest that CCN2 may regulate dental epithelial cell differentiation via reduced ERK1/2 phosphorylation by forming a complex with BMP4 as a novel modulator of BMP signaling. Taken together, the results suggested that CCN2 acts as an autocrine factor during odontogenesis and regulates dental epithelial cell proliferation and differentiation in cooperation with FGF2 and BMP4. Further investigation into the molecular mechanisms of CCN2 and FGF or the BMP signaling responsible for dental epithelial cells will be crucial in order to find an effective therapy for tooth regeneration.
We used a primary bovine dental epithelial cell culture system in order to study the process of differentiation while maintaining the characteristics of cells in vivo. Cells in primary culture have an advantage of very closely mimicking the physiological state of cells, and rodent odontogenic epithelial cells are conventionally used for in vitro studies. However, the preservation of the primitive cell structure, cell-specific metabolism, and cell–cell interactions in vitro are often limited by the restricted number of cells available. On the other hand, a primary culture system using bovine odontogenic epithelial cells has a great advantage in terms of the stability and maintenance of the primitive structure, because it yields a large number of high-grade, freshly isolated cells without any passage. The epithelial cells are able to differentiate into ameloblasts and are initiated to express the differentiation marker alkaline phosphatase. Alkaline phosphatase is the earliest and most reliable molecule for the differentiation of ameloblasts in the presence of ascorbate and of ascorbate and β-GP. These data correlate well with our previous study using a bovine dental mesenchyme primary culture system, in which type I collagen and dentin sialophosphoprotein mRNAs were up-regulated under treatment with ascorbate and β-GP (15). We thus suggest that this culture system may have good potential for investigating the developmental stages of the dental epithelium.
By utilizing this conventional dental epithelial cell culture system, we have demonstrated that the effects of CCN2 in the inner dental epithelial cells are important for the intricately orchestrated processing of cellular proliferation and differentiation along with FGF and BMP signaling for the proper early tooth germ development.
Footnotes
-
Conflicts of Interest
None.
- Received November 11, 2014.
- Revision received November 23, 2014.
- Accepted November 26, 2014.
- Copyright © 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.