


Abstract
Background/Aim: Uterine quiescence at the time of embryo transfer is a prerequisite for successful in vitro fertilization (IVF). This study assessed whether prostaglandin-induced contractions in the perfused swine uterus can be reduced by progesterone. Materials and Methods: Fifty-eight non-pregnant swine uteri were perfused using an established extracorporeal perfusion model. Intrauterine pressure changes during perfusion with prostaglandin (PG) administration (PGE1, PGE2, PGF2α) and progesterone (1 pg/ml, 10 pg/ml, 25 pg/ml, 50 pg/ml) were assessed using an intrauterine double-chip microcatheter. Results: The contraction-stimulating effect of PGs was clearly reduced by progesterone. Only PGE1 still triggered relevant contractions during continuous perfusion with progesterone solution, up to a concentration of 10 pg/ml. With PGE2 and PGF2α, a clear reduction of uterine contractility was observed even at at a progesterone concentration of 1 pg/ml. Conclusion: The extracorporal perfusion model of swine uteri shows that PG-induced contractions can be reduced in a dose-dependent manner by progesterone.
Adequate uterine contractility is involved in the transport of semen and gametes, and in successful embryo implantation, whereas inadequate uterine contractility may lead to ectopic pregnancies, miscarriages, retrograde bleeding and endometriosis (1-3). According to Mansour and Aboulghar (4), the key factors for improving embryo transfer technology include avoiding uterine contractions. It is known that a sufficiently relaxed myometrium at the time of embryo implantation is extremely important for successful reproduction (2, 5, 6) Evidently, not only initiating uterine contractions in order to assist directed sperm transport during the preovulatory phase, but also suppressing contractions in order to provide optimal conditions for implantation, may offer promising and innovative options for optimizing artificial reproduction (4-6). On the basis of the developments reported by Bulletti et al. (7), Dittrich (8) and Maltaris (9) in 2003 a model was established for the perfusion of swine uterus. Such a model is suitable for investigating the effects of a wide variety of medications in large numbers of organs under physiological conditions (8, 10). Although there are anatomic and physiological differences between human and swine uterus, the microscopic structures of the organs are similar. The regulatory control system for endocrine functions also allows comparison, particularly as progesterone concentration increases at the typical time for embryo implantation in human and swine.
The autonomic innervation of the swine myometrium is normally both parasympathetic and sympathetic. The smooth-muscle fibres of the uterus are described as being of the single-unit type, and excitation spreads across intercellular gap junctions. Pacemaker potentials are therefore able to create spontaneous contractions. Muscle tone in the myometrium is still myogenic in the model, so that although the organ is isolated, it is possible to generate spontaneous peristaltic contractions (11, 12). It is also possible to apply agents and medications such as oxytocin, tocolytics, spasmolytics, steroids, prostaglandins (PGs) and seminal plasma, and to examine their effects on the myometrium (8, 9, 11, 13-15).
The investigation of uterine contractility and the way in which it is regulated by PGs are of clinical interest not only in obstetrics–for example, to induce labour or to delay birth and prolong pregnancy–but also in gynaecology and reproductive medicine, especially in relation to in vitro fertilization (IVF) and embryo implantation (2). Although PGE and PGF receptor expression and myometrial sensitivity in different parts of the uterus and myometrium have been described in pregnancy and during labour, there is still a lack of comparable data for the human non-pregnant uterus (16). Yang et al. (17) reported expression of PG receptors primarily in the circular muscle of mouse myometrium during early pregnancy, and it has been suggested that this is the primary target for the PG-mediated uterine contractions required for transport and embryo implantation in the uterus. Müller et al. investigated the effects of different PGs on myometrial contractility, measuring intraluminal pressure changes in the complete uterus in a perfusion model for non-pregnant swine uterus independent of the stage of the oestrous cycle (13, 14).

A bilaterally cannulated swine uterus.
The aim of the present study was to assess whether it is possible to reduce contractions induced by PGE1, PGE2 and PGF2α in the perfused swine uterus using progesterone perfusion, as uterine quiescence at the time of embryo transfer is an important prerequisite for successful IVF (5).
Materials and Methods
Swine uterus. Non-pregnant swine uteri were collected from the abattoir in Erlangen early in the morning, immediately after killing (with electric shocks, 1.5 A, 400 V, 4 s). The organs were all from healthy swine aged 5-18 months. One uterus per day was selected for the experiments on the basis of size, weight, general condition and cannulability of the uterine arteries. The typical time of implantation post conception is 5-6 days in humans and 13-14 days in swine.
Cannulation. The selected uterus was dissected in a standardized way for cannulation. The uterine arteries were released from the surrounding connective tissue in the broad ligament of the uterus bilaterally as far as the adventitia. This was followed by cannulation of the arteries using 16-G Abbocaths, which were fixed in place using Vicryl 3-0 sutures (Ethicon Johnson & Johnson International Inc., Brussels, Belgium) (Figure 1). Correct positioning of the Abbocaths and patency of the uterine vascular system were checked using careful rinsing of the two arteries, each with 2.5 ml NaCl 0.9%. In addition, the tubes and the vascular plexus surrounding the ovary were ligated to prevent the perfusion solution from escaping at these points and to establish and maintain better uterine perfusion pressure. This was also intended to minimize any disturbing hormonal effects of oestrogens and gestagens from the ovaries.
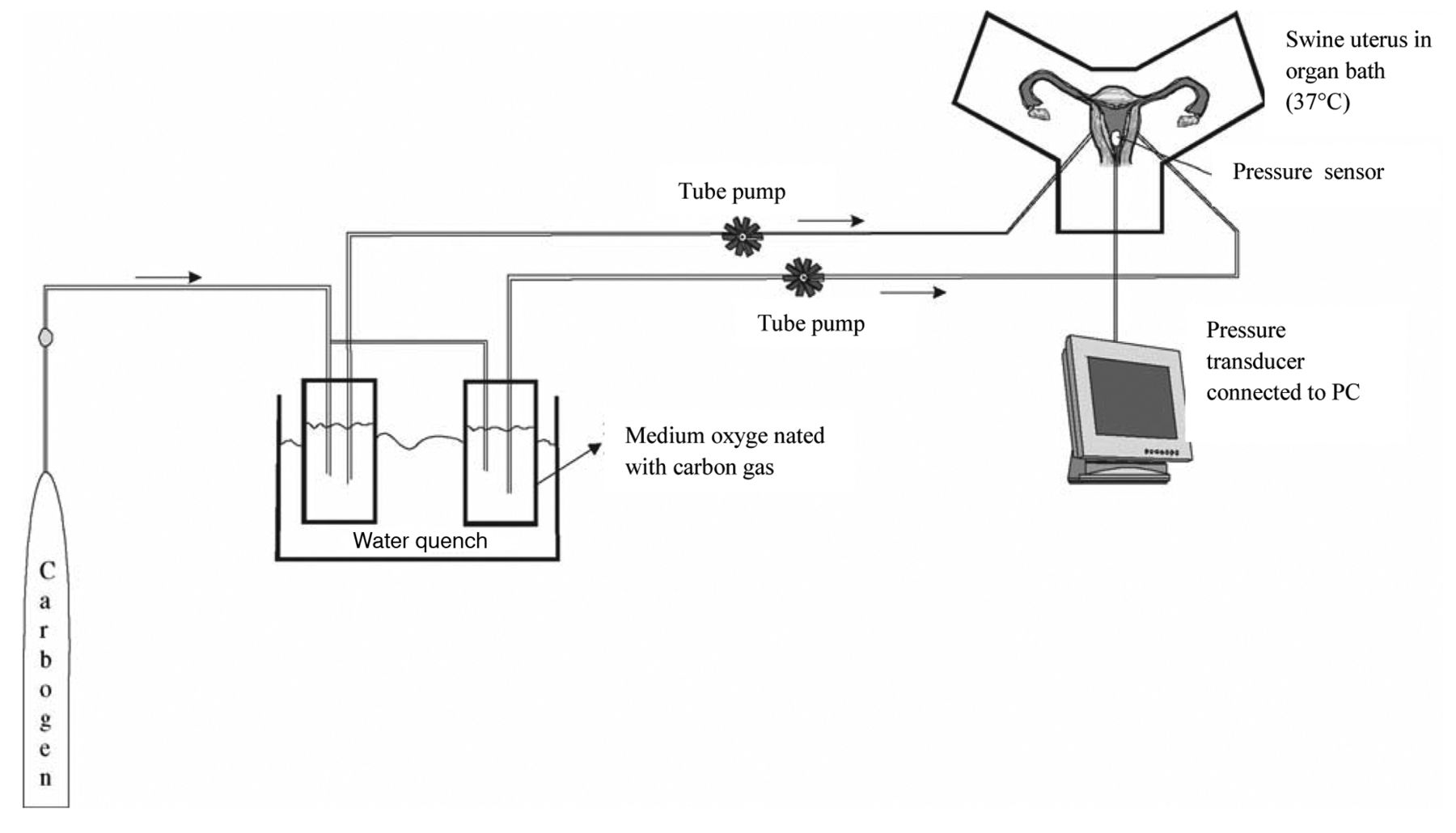
Perfusion system. Following cannulation, the organ that had been prepared for perfusion was placed in an organ bath at a set temperature. The organ bath initially consisted of 1 litre of perfusion medium warmed to 37°C. During the entire experiment, the temperature was kept constant at between 36.5°C and 37.5°C (temperature measurement probe, Raumedic; Rehau Ltd., Rehau, Germany). The perfusion medium was warmed to a constant 37°C in a shaking water-bath and oxygenated with carbogen (95% O2, 5% CO2; Linde Inc., Frankfurt, Germany). The Abbocaths were connected with a silicon tube system. A roller pump (Heidolph Ltd., Kelheim, Germany) transported the medium in the tube system at a constant flow rate into the organ's arterial system. The flow rate was increased gradually over 10-15 min to approximately 15 ml/min to ensure a perfusion pressure of approximately 80-100 mmHg. The perfusate flowed continuously through the uterine open vascular system into the organ bath – i.e. there was no re-circulation of the medium into the arterial system. The diagram in Figure 2 shows the experimental design.
Perfusion medium and drugs
Krebs–Ringer bicarbonate buffer (KRBB) solution. Physiological KRBB has been successfully used in our laboratory since 2003 for experiments in the perfused swine uterus (8, 13, 14). The solution is prepared in the institution in Erlangen in accordance with instructions from the manufacturers, Sigma-Aldrich Ltd. (Steinheim, Germany) (Table I).
Progesterone was used as a water-soluble dry substance (Sigma-Aldrich Ltd.) and diluted with KRBB so that when added to 1 litre of perfusion solution, a concentration of 1 pg progesterone per millilitre of medium was produced. The same procedure was used for the selected concentrations of 10 pg/ml, 25 pg/ml and 50 pg/ml of progesterone solution.
PGs were diluted with NaCl 0.9% at different concentrations in such a way to provide individual doses of 0.5 μg/ml, 2.0 μg/ml, 4.0 μg/ml, 6.0 μg/ml, 8.0 μg/ml and 10.0 μg/ml, which were administered as 1-ml boluses.

Diagram showing the experimental perfusion model used.

A double-chip catheter for intrauterine pressure measurement (Urobar 8 DS-F; Raumedic, Rehau Ltd., Rehau, Germany).
PGE1 derivative. Minprog 500 μg/ml; active agent: alprostadil (Pharmacia Ltd., Karlsruhe, Germany); PGE2 derivative: Nalador 500; active agent: sulprostone (Schering Deutschland Ltd., Berlin, Germany); PGF2α derivative. Enzaprost F5; active agent: dinoprost (Chinoin Ltd., Budapest, Hungary).
Intrauterine pressure measurement. Intrauterine pressure (IUP) was recorded using a double-chip microcatheter (Urobar 8 DS-F; Raumedic; Rehau Ltd.) (Figure 3). The measurement sensors were incorporated into the catheter at intervals of 8 cm, so that after careful introduction, the distal sensor (IUP1) was positioned without tension in one of the uterine horns and the proximal sensor in the body of the uterus (IUP2). The pressure catheter was then fixed in place with a single button suture at approximately 11 cm to prevent it from sliding out during the measurements. After this, the pressure catheter was connected with the monitoring device (Datalogger, MPR1; Raumedic, Rehau Ltd.).

Intrauterine pressure changes (IUP) during PGE1 administration and KRBB perfusion compared to perfusion with 1 pg/ml progesterone; n=3 (blue bar=uterine horn; red bar=body of the uterus).
The Datalogger allowed simultaneous and continuous recording of pressure changes at both electrodes (IUP1, IUP2) and continuous temperature measurement. A pressure value in millimetres of mercury was recorded by the Datalogger for each second. Pressure conditions in the vascular and tube system were displayed using a central venous pressure measurement module.
The measurement electrodes IUP1 and IUP2 and central venous pressure were calibrated to 0 mmHg at the start of each experiment and the corresponding recording. The course of the experiment was observed on a graphic display and finally evaluated with the help of the corresponding software (Datalogg; Raumedic, Rehau Ltd.).
Vitality parameters. Perfusate samples were taken at 1-hour intervals for measurement of pH (7.35-7.45), pO2 (65-100 mmHg), pCO2 (35- 45 mmHg), HCO3 (21-26 mmol/l), lactate (0.55-2.2 mmol/l) and oxygen saturation (95-98%). The perfusate samples were analysed with an i-STAT portable clinical analyser (Abbott Diagnostics, Lake Forest, Illinois, USA).
Data processing and statistics
Data processing. For each uterus, data for pressure changes were recorded and analysed continuously. Absolute maximum pressure values (IUP) for each uterine contraction were counted and the mean values were analysed. Additional evaluation of the data was carried out using Windows Excel.
Statistical analysis. A paired Student's t-test was used to evaluate differences in pressure changes at IUP1 and IUP2, with significance set at p<0.05.
Experimental procedure. Test uteri (n=9): To demonstrate the contraction-stimulating effect of the PGs and their interaction with progesterone, three test uteri for each PG derivate were prepared. These test uteri were perfused for 1h with physiological KRBB, followed by administration of a 6-μg/ml bolus of PGE1, PGE2, or PGF2α. After this, these test uteri were each perfused with 1 pg/ml progesterone solution for a further hour, and finally 6 μg/ml of PGE1, PGE2 or PGF2α was again administered as a bolus.
Experimental series (n=49): Each series was started with a concentration of 1 pg/ml progesterone as the perfusion solution. After 1 hour of perfusion, the first bolus of PGE1, PGE2, or PGF2α was injected. For each uterus and for each drug concentration, a time interval of 10 minutes before and after drug administration was recorded and evaluated. After continuous rinsing with perfusion solution for 20 minutes, the next drug concentration was administered, if the contractions have returned to the baseline, which was the case in all uteri tested. Altogether each PG investigated was administered with increasing dosages (0.5 μg/ml, 2.0 μg/ml, 4.0 μg/ml, 6.0 μg/ml, 8.0 μg/ml, 10.0 μg/ml) every 30 min. The same procedure was then used for the other progesterone concentrations of 10 pg/ml, 25 pg/ml and 50 pg/ml.
Results
A total of 58 non-pregnant swine uteri were perfused. The mean weight of the organs was 123.3 g (range=65-188 g). All perfused uteri initially showed spontaneous rhythmic contractions. The vitality parameters remained physiological during the perfusion period [data not shown; for details, see Dittrich et al. 2003 (8)].
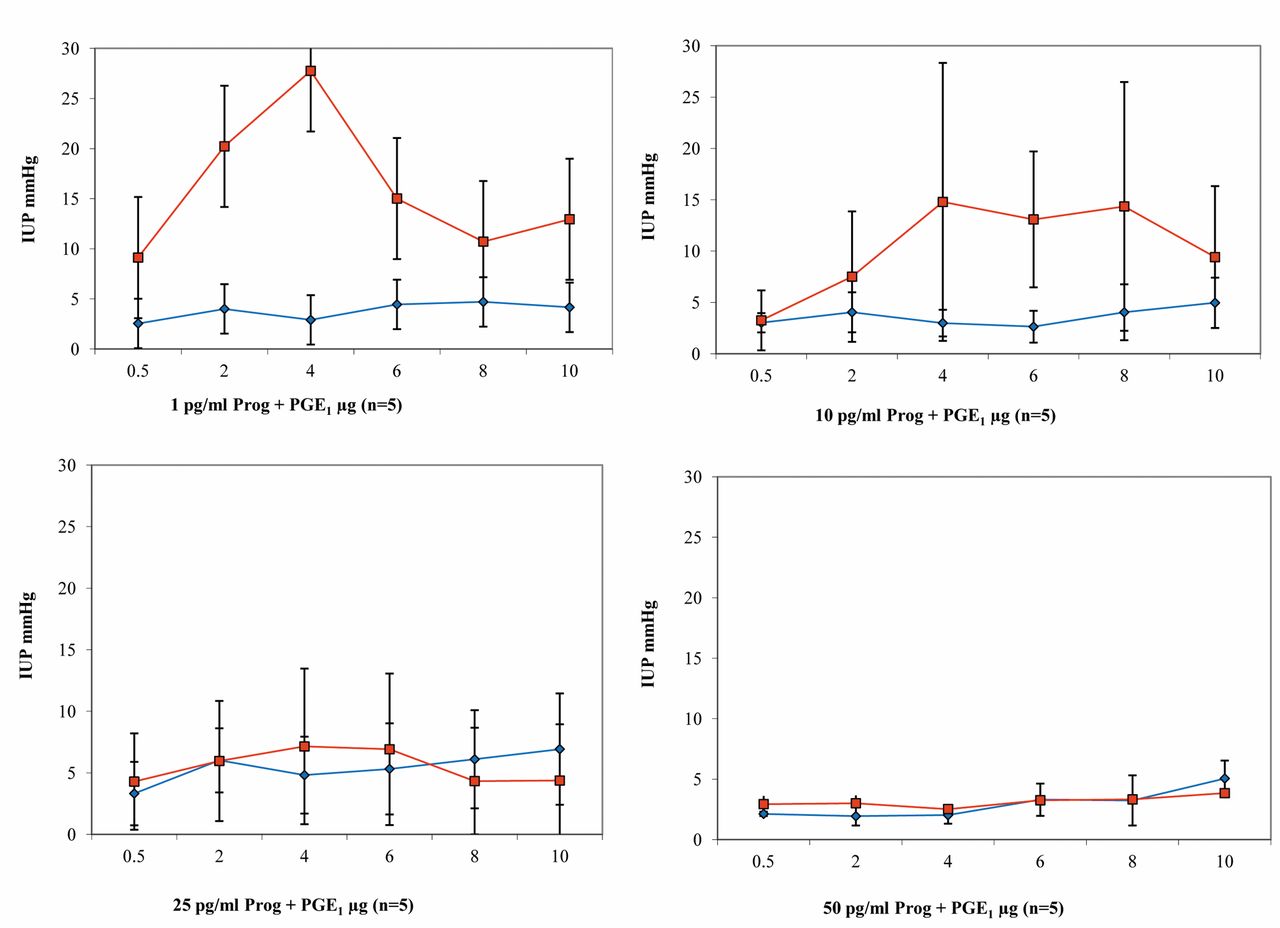
Results of perfusion with progesterone and PGE1. Test uteri (n=3). During perfusion with physiological KRBB, a marked increase in pressure was observed after administration of contraction-stimulating PGE1. The contraction-inhibiting effects of the progesterone nutrient solution then became clear both at the uterine horn and in the body of the uterus (Figure 4).
Progesterone and PGE1 (n=18): During perfusion with 1 pg/ml progesterone solution, a dose-dependent increase in IUP was observed in the uterine body after administration of PGE1. Intrauterine pressure reached a peak in the body of the uterus of up to 27.8 mmHg after bolus administration of 4 μg/ml PGE1. During the entire series of measurements, much higher IUP values were generally measured in the body of the uterus than in the uterine horn (Figure 5). With a progesterone nutrient solution of 10 pg/ml and a PGE1 dosage of 4-8 μg/ml, a plateau was reached in the body of the uterus. Much lower pressure values were again observed in the uterine horn than in the body (Figure 5). With a progesterone perfusion solution of 25 pg/ml, clear inhibition of the PG effect was observed. The IUP values were stable at a low level at both measurement points, independently of the PG dosage, and did not exceed 7.5 mmHg during the entire measurement period (Figure 5). This inhibitory effect of progesterone became even clearer at the highest concentration of 50 pg/ml progesterone. The uteri hardly reacted at all any more to the contraction-stimulating effect of the PG studied. The pressure values at both measurement points did not exceed the 5-mmHg mark during the entire measurement period (Figure 5).

Results of intrauterine pressure changes (IUP) during PGE1 administration and perfusion with increasing dosages of progesterone (blue line/rhombus=uterine horn; red line/quadrat=body of the uterus).
Results of perfusion with progesterone and PGE2. Test uteri (n=3). During perfusion with KRBB, a contraction-stimulating effect of PG was observed after administration of PGE2, particularly in the uterine body. Higher pressure values were noted in the body than at the uterine horn during PGE2 administration. During progesterone perfusion, there was inhibition of contractility in the uterine body, with a trend towards increased pressure at the uterine horn (Figure 6).
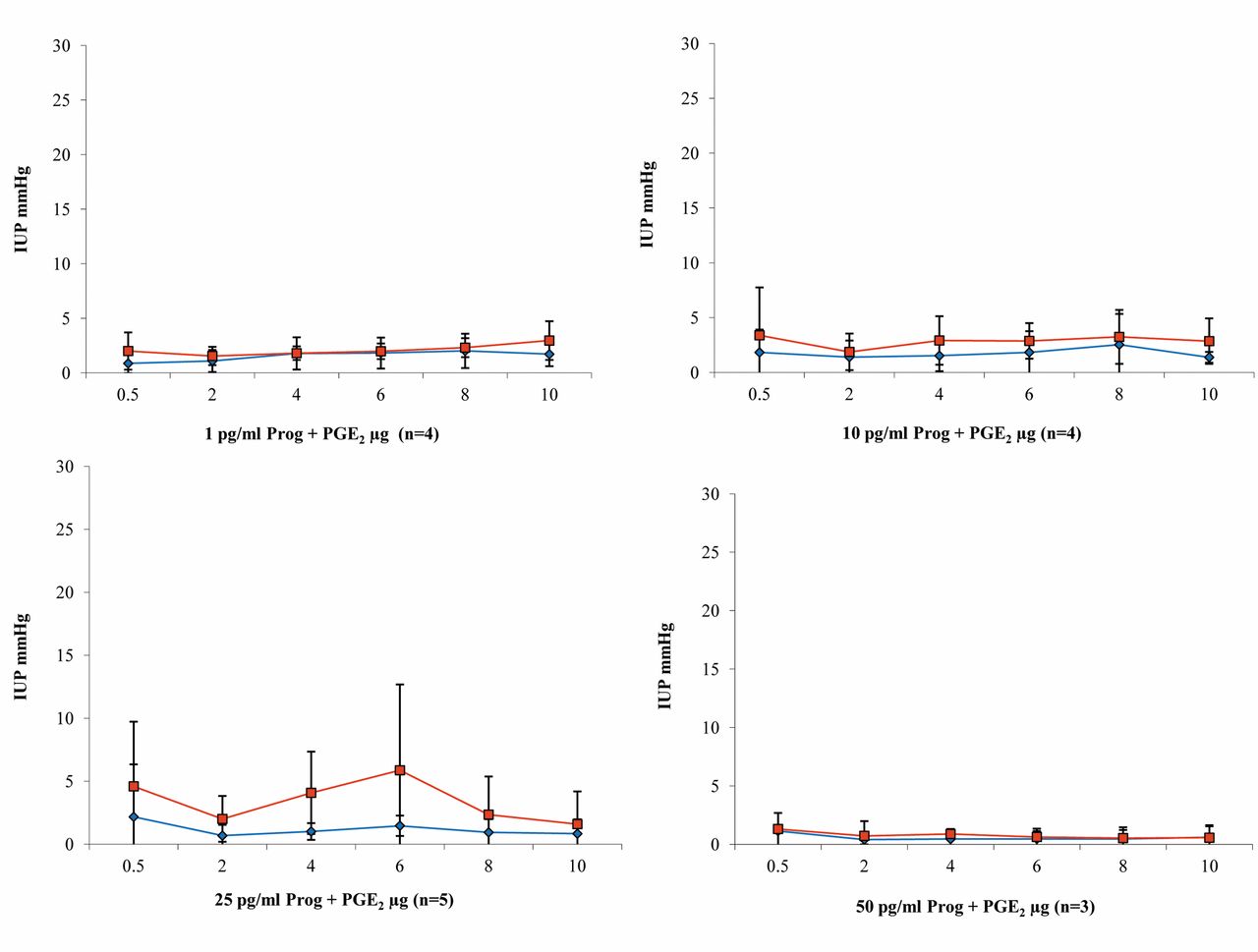
Progesterone and PGE2 (n=16). Relevant increases in pressure were only seen during perfusion with 25 pg/ml progesterone nutrient solution, with a maximum pressure of 5.87 mmHg in the uterine body after administration of 6 μg/ml PGE2. No relevant increases in contractility due to the addition of PGE2 were observed with any of the other progesterone concentrations. Overall, pressures in the uterine body were slightly higher than at the uterine horn (Figure 7).

Intrauterine pressure changes (IUP) during PGE2 administration and KRBB perfusion compared with perfusion with 1 pg/ml progesterone; n=3 (blue bar=uterine horn; red bar=body of the uterus).
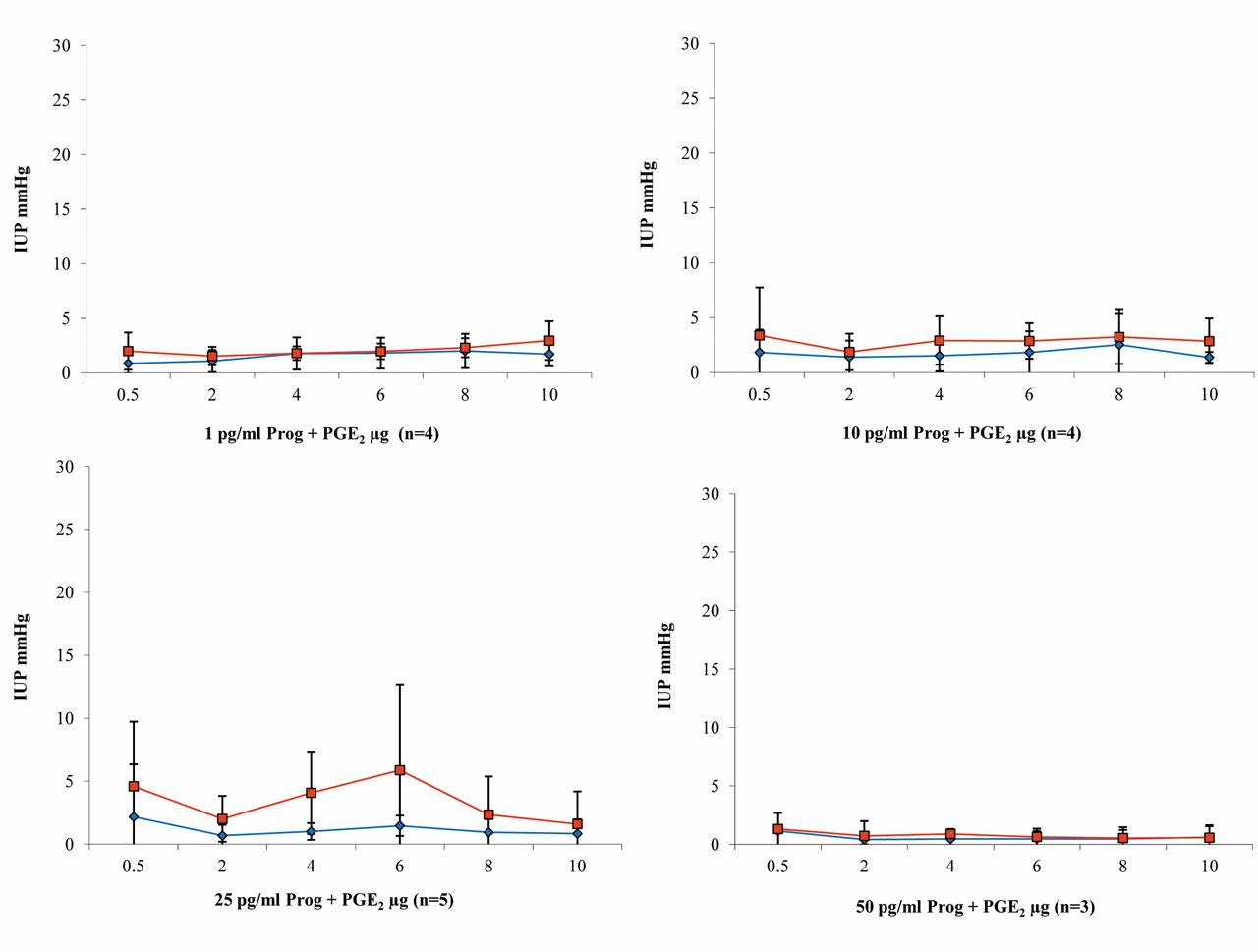
Results of intrauterine pressure changes (IUP) during PGE2 administration and perfusion with increasing dosages of progesterone (blue line/rhombus=uterine horn; red line/quadrat=body of the uterus).
Results of perfusion with progesterone and PGF2α. Test uteri (n=3). Both with physiological KRBB perfusion and also during perfusion with progesterone solution, PGF2α produced the lowest pressure values among all three PGs. During perfusion with KRBB, pressure at the uterine horn was higher than in the body, in contrast to the findings with PGE1 and PGE2. During progesterone perfusion, basically no contractions were triggered by the addition of PGF2α. The values at both measurement points were practically zero (Figure 8).
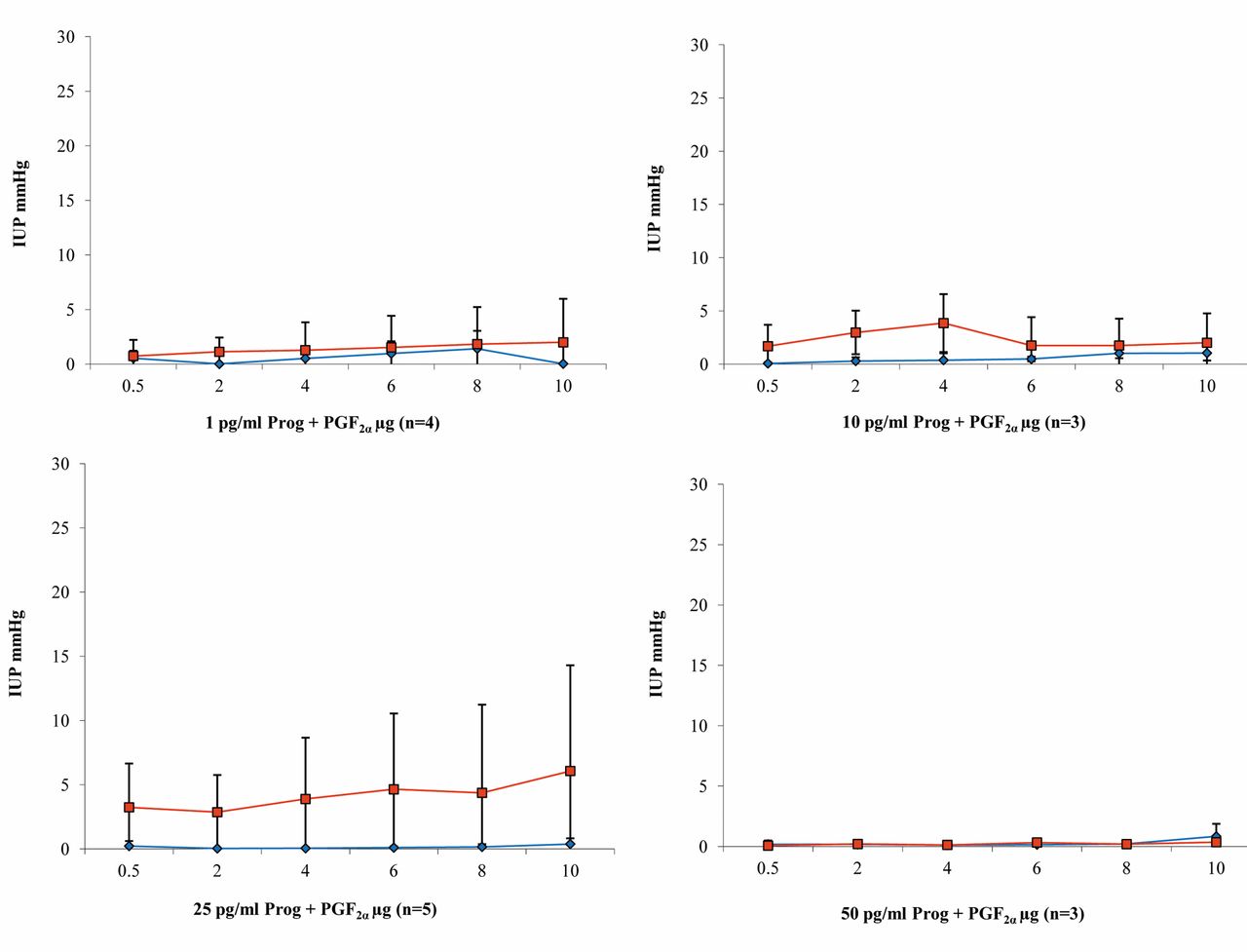
Progesterone and PGF2α (n=15). Pressures higher than 2.5 mmHg were never achieved either in the uterine body or in the horn during perfusion with 1 pg/ml progesterone solution (Figure 9). With 10 pg/ml progesterone, pressure in the body rose to 3.85 mmHg at 4 μg/ml PGF2α and then declined again with further increases in the PGF2α dosage up to the end of the measurements and no longer exceeded 2.5 mmHg. Lower pressures were always observed in the uterine horn than in the body (Figure 9). The effect of PGF2α at the uterine horn was completely inhibited by perfusion with the 25 pg/ml progesterone solution. In the uterine body, the pressure values increased slowly with increasing concentrations of PGF2α and reached a maximum of 6.07 mmHg at 10 μg/ml PGF2α (Figure 9). The uteri did not react to any administration of PGF2α during perfusion with 50 mg/ml progesterone (Figure 9).
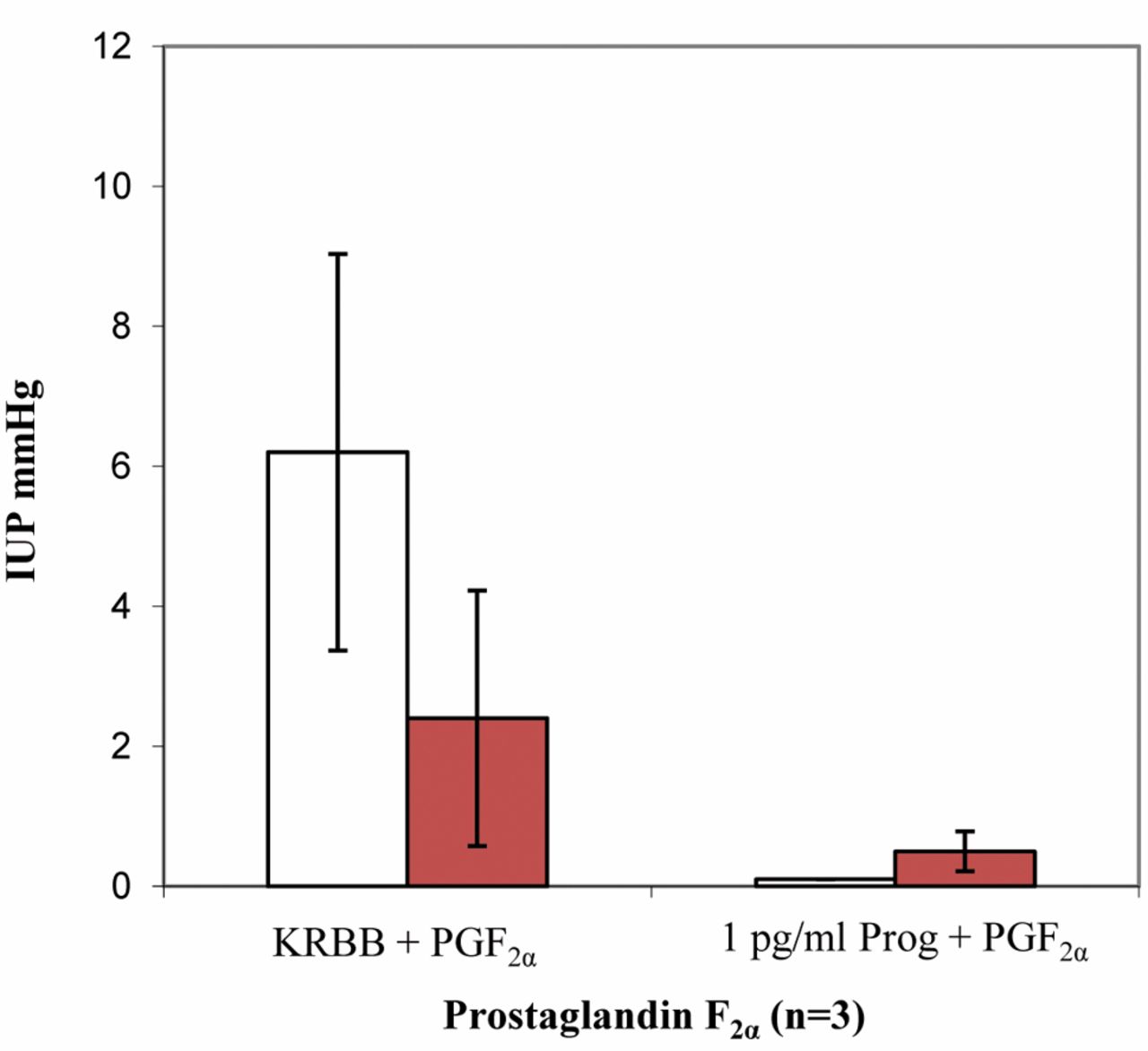
Intrauterine pressure changes (IUP) during PGF2α administration and KRBB perfusion compared to perfusion with 1 pg/ml progesterone; n=3 (blue bar=uterine horn; red bar=body of the uterus).

Results of intrauterine pressure changes (IUP) during PGF2α administration and perfusion with increasing dosages of progesterone (blue line/rhombus=uterine horn; red line/quadrat=body of the uterus).
Summary of the results. During perfusion with KRBB, PGE1 and PGE2 showed a contraction-stimulating effect, which was stronger in the uterine body (Figures 4 and 6); the opposite was true with PGF2α (Figure 8). The strongest contractions were caused by PGE2 (Figure 6). Overall, the effect of all of the prostaglandins studied was markedly reduced by progesterone (Figures 4, 6 and 8). Only PGE1 was capable of triggering relevant contractions in the uterine musculature during continuous perfusion with progesterone solution, up to a concentration of 10 pg/ml (Figure 5). In the other two test series with PGE2 and PGF2α, clear reduction of uterine contractility was already observed at a progesterone concentration of 1 pg/ml (Figures 7 and 9). The inhibitory effect of progesterone concerning PG induced contractions did not reach statistical significance, but Figures 5, 7 and 9 illustrate a stable and clear tendency towards a dose dependency.
Discussion
Prostaglandin is secreted by the seminal vesicle and prostate gland and is present in human and boar seminal plasma (18-20). In addition, the human and swine endometrium itself is able to generate different PGs, which may play a significant role in placentation and in regulating the lifespan of the corpus luteum (21). Wasielak et al. (22) evaluated the effect of the embryo on uterine PG release and synthesis during the peri-implantation period in the pig and they suggested a local effect of the embryo on PG release pathways but also a more systemic effect within the whole uterus with regard to PG synthesis and accumulation in uterine tissues. Cyclooxygenases (COX1 and COX2) are the crucial enzymes responsible for the synthesis of various PGs. The unique expression pattern of Cox1 and Cox2 genes in pre-implantation mouse uterus suggests an important role for PGs in embryo implantation (23). Recently, St. Louis et al. reported that COX2 expression is regulated by steroid hormones that play a crucial role in embryo implantation through PG synthesis (24). Achache et al. identified defective endometrial PG synthesis in patients with repeated implantation failure undergoing IVF and suggested that reduced PG synthesis in the human endometrium may lead to poor endometrial receptivity (25). Blockade of PG synthesis before or during the time of implantation causes complete inhibition, a delay in implantation, or a reduction in the number of implantation sites with diminished decidual tissue (23).
In summary, in the literature there is considerable evidence regarding the role of PGs in human pregnancy and in particularly at the ‘window of implantation’, whereas their possible role in the regulation of uterine contractility in non-pregnant uteri and its implications for successful IVF are still enigmatic and are not completely understood.
In addition to oestrogen and progesterone, as well as oxytocin, PGs are also believed to modulate myometrial contractility in a characteristic way not only at the time of labour, but also at the time of human reproduction (26). In a study of myometrial PG receptor expression, a region-dependent, heterogeneous distribution of contractile and relaxant prostanoid receptors and cyclooxygenase was reported in the non-pregnant porcine myometrium (27). In a perfusion experiment in non-pregnant swine uteri by Müller et al. (13), a dose-dependent increase in intrauterine pressure after the administration of PGF2α was demonstrated, which reached a plateau after further stimulation. A dose-dependent increase in IUP was also observed after the administration of PGE1 and PGE2, with a plateau in IUP in the middle concentration range and a decrease during the further course of stimulation. The authors stated that PGs may play an important role in the regulation of contractility in non-pregnant uteri. Chiossi et al. (28) investigated the effects of PGE1 and PGE2 on in vitro myometrial contractility. Treatment with PGE1 significantly increased myometrial contractions in comparison with PGE2.
Progesterone is a hormone naturally produced by the corpus luteum and the placenta in early pregnancy and is essential for maintaining pregnancy. For the genomic pathway, progesterone interacts with its nuclear progesterone receptor, which decreases expression of contraction-associated genes and maintains uterine quiescence. For the non-genomic pathway, progesterone interacts with membrane-associated receptors to modulate directly intracellular calcium and cyclic adenosine monophosphate levels (29). Details of immune aspects and myometrial action of progesterone have been reviewed by Vrachnis et al. (30) An experimental study on isolated porcine myometrial cells by Franczak et al. (31) reported that the effects of oxytocin on PG secretion and Ca2+ accumulation are completely inhibited by progesterone during pregnancy.
Bulletti et al. (6) studied the hormonal regulation of the menstrual cycle and concluded that uterine quiescence and the dominance of convergent uterine contractility prevailing at the time of implantation are linked to progesterone. These findings were confirmed in the swine uterus perfusion model by Müller et al. (13, 14). Fanchin et al. (5) showed that high-frequency uterine contractions on the day of embryo transfer hinder the outcome during IVF, possibly by expelling embryos out of the uterine cavity.
The results of the present experimental study show a distinct tendency for PG-induced contractions of the swine myometrium to be reduced in a dose-dependent manner by continuous progesterone perfusion. Progesterone, as well as PGs, affects intracellular calcium metabolism of the uterus. Although there is a lack of data concerning the interactions between PGs and progesterone on uterine contractions, there is some evidence that the inhibitory effect of progesterone in this study may be mainly non-genomic and secondary due to decrease of intracellular Ca2+ accumulation (29, 31). Observations by Fanchin's (32) group confirm the uterus-relaxing effects of progesterone in the non-pregnant uterus and support the administration of progesterone before embryo transfer to increase tissue concentrations and improve the outcome of IVF.
Conclusion
The extracorporal perfusion model of swine uterus shows that PG-induced contractions can be reduced in a dose-depend manner by progesterone. Further research is required to clarify the cellular effects of the interactions between progesterone and PGs, but the observed inhibitory effect of progesterone concerning PG induced contractions supports the current practice of administering progesterone to achieve uterine quiescence at the time of embryo transfer.
Footnotes
-
This article is freely accessible online.
-
Conflicts of Interest
All Authors hereby state that there were no actual or potential conflicts of interest, including any financial, personal or other relationships with other people or organizations within three years of beginning the submitted work that could inappropriately influence, or be perceived to influence their work.
- Received February 5, 2014.
- Revision received March 4, 2014.
- Accepted March 5, 2014.
- Copyright © 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}