


Abstract
Background: The anti-inflammatory activity of magnolol and related compounds is currently a focus of interest. In the present study, the inhibitory effects of these compounds on cyclooxygenase (COX-2) expression and nuclear factor-kappa B (NF-κB) activation were investigated in RAW264.7 macrophage-like cells stimulated with the fimbriae of Porphyromonas gingivalis, an oral anaerobe. Materials and Methods: The cytotoxicity of magnolol, honokiol, eugenol and bis-eugenol against RAW264.7 cells was determined using a cell counting kit (CCK-8). The regulatory effect of these compounds on the expression of COX-2 mRNA, stimulated by exposure to the fimbriae was investigated by real-time polymerase chain reaction (PCR). NF-κB activation was evaluated by enzyme-linked immunosorbent assay (ELISA)-like microwell colorimetric transcription factor activity assay (Trans-AM) and western blot analysis. The radical-scavenging activity was determined using the induction period method in the methyl methacrylate-azobisisobutyronitrile (AIBN) polymerization system under nearly anaerobic conditions. The phenolic bond dissociation enthalpy (BDE) and orbital energy were calculated at the density functional theory (DFT) B3LYP/6-31G* level. Results: The cytotoxicity against RAW264.7 cells declined in the order bis-eugenol>eugenol> honokiol>magnolol, whereas the radical-scavenging activity declined in the order honokiol, bis-eugenol>magnolol> eugenol. Magnolol and honokiol significantly inhibited the fimbria-induced expression of COX-2 at non-cytotoxic concentrations. Both the fimbria-stimulated binding of NF-κB to its consensus sequence and phosphorylation-dependent proteolysis of inhibitor κB-α were markedly inhibited by magnilol and honokiol, whereas eugenol and bis-eugenol did not inhibit COX-2 expression and NF-κB activation. Magnolol and honokiol possessed a high electronegativity (χ) value. Conclusion: Magnolol and honokiol exhibit antioxidative activity, low cytotoxicity, and anti-inflammatory activity. These compounds may be capable of preventing chronic inflammatory diseases induced by oral bacteria.
Cyclooxygenase (COX)-2 is the key enzyme that catalyzes the two sequential steps responsible for the biosynthesis of prostaglandins (PGs) from arachidonic acid. COX-2, the inducible isoform of COX, plays a critical role in renal function, maintenance of gastrointestinal integrity, ovarian and uterine function, bone metabolism, various inflammatory responses, rheumatoid arthritis, and Alzheimer's disease, and its overexpression has been associated with several types of pathologies, including neurodegenerative diseases and various types of cancers (1). Transcription of its gene results in activation of cis-acting transcription factors, such as nuclear factor-kappa B (NF-κB), activator protein-1 (AP-1), and other effector elements in a variety of cells (2-4).
The stem bark of Magnolia officinalis REHD. et WILS. (Magnoliaceae) has long been used in Chinese and Japanese traditional medicine for the treatment of fever, headache, anxiety, diarrhea, asthma, and stroke, and has potent anti-inflammatory effects (5). Magnolol and honokiol are neolignane derivatives containing biphenolic structures extracted from the bark of M. officinalis. These two compounds are structural isoforms resembling the dimer of eugenol (4-allyl-2-methoxyphenol), which is a component of clover oil (6), and are responsible for the main pharmacological effects of the bark, which demonstrates antioxidant and anti-inflammatory effects (7-10). We have previously reported that the dimer of butylated hydroxyanisole (bis-BHA: 3,3’-di-tert-butyl-5,5’-dimethoxy-1,1’-biphenyl-2,2’-diol), a potent antioxidant, exerts a preventive effect against AP-1 activation and COX-2 expression in macrophages stimulated with fimbriae of Porphyromonas gingivalis, which is the major pathogen responsible for chronic periodontitis. In addition, we recently reported on the inhibitory effects of melatonin, the major secretory product of the pineal gland that is mostly associated with regulation of the circadian dark/light rhythm of the human body, on fimbria-stimulated COX-2 expression via suppression of NF-κB activation (11). This experimental model using macrophages has been shown to be reliable for evaluating the anti-inflammatory activity of phenolic antioxidants (12). A possible etiological link between Buerger disease and chronic infections such as those due to oral bacteria has been reported (13). In addition, the metabolites of this organism can promote oral cancer in epithelial cells and induction of epithelial–mesenchymal transition at sites of chronic inflammation (14).
Therefore, it would be of interest to determine whether COX-2 expression and NF-κB activation could be suppressed by magnolol and honokiol and their related compounds eugenol and bis-eugenol in RAW 264.7 cells stimulated with P. gingivalis fimbriae. If this were possible, then magnolol and honokiol might contribute not only to the prevention of oral diseases, such as chronic infectious diseases, and neoplastic diseases such as oral squamous cell carcinoma (15), but also to the regeneration of alveolar bone through stimulation of type I collagen fiber production and modulation of osteoblastic and osteoclastic activity derived from cellular mediator proteins such as receptor activator of NF-κB ligand (RANKL) caused by P. gingivalis and related bacteria.
In the present study, we investigated whether magnolol, honokiol, eugenol and its dimer (bis-eugenol) are able to inhibit P. gingivalis fimbria-induced COX-2 expression and NF-κB activation. The radical-scavenging activity of these compounds was also investigated using the induction period method for polymerization of methyl methacrylate (MMA) initiated by thermal decomposition of azobisisobutyronitrile (AIBN) under nearly anaerobic conditions. Furthermore, theoretical parameters were investigated by the density fuction theory (DFT) B3LYP/6-31G* method. A possible link between COX-2 inhibition was investigated using chemical hardness (η) and electronegativity (χ) principles, in addition to the bond dissociation enthalpy (BDE) value.
Materials and Methods
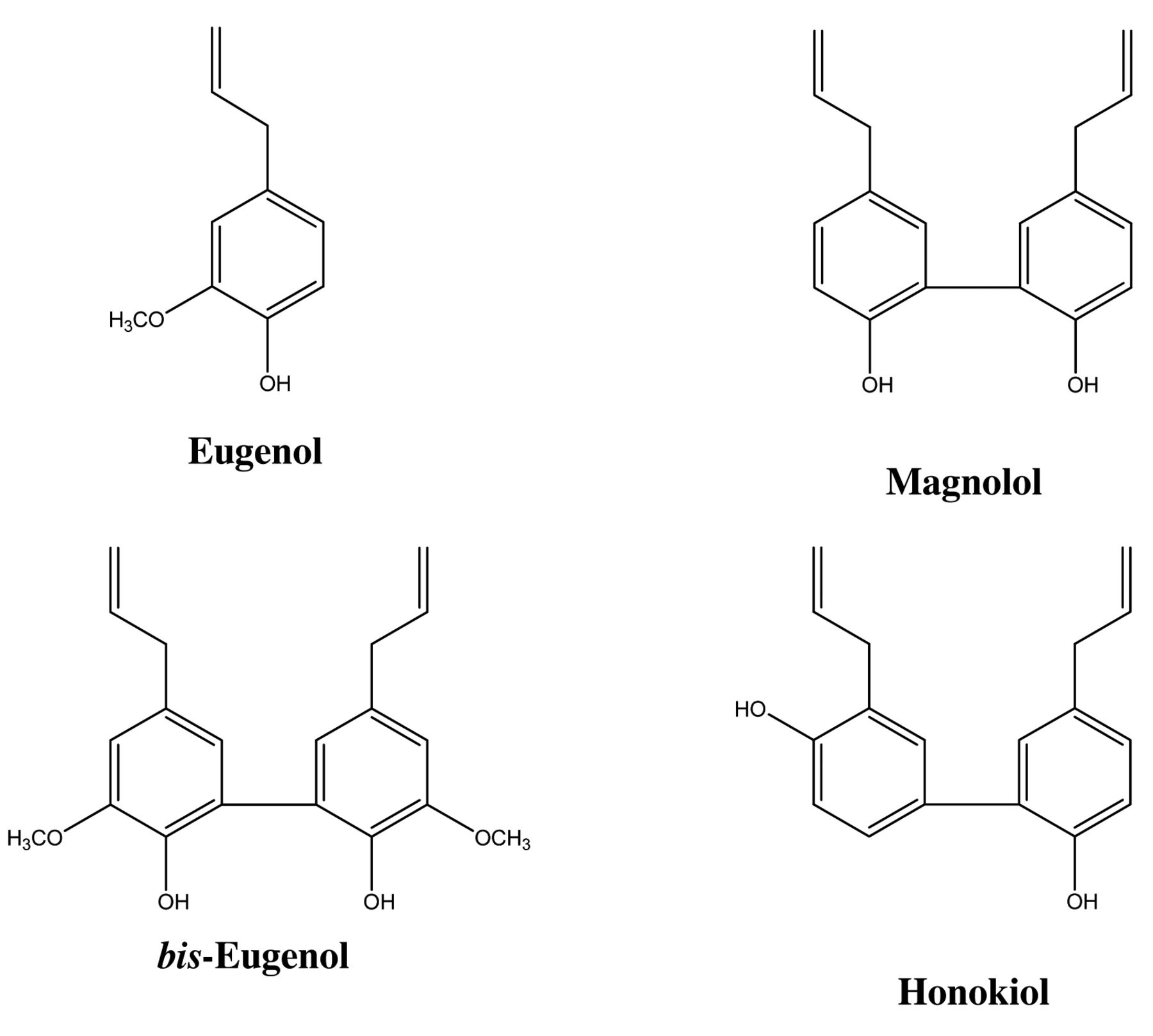
Materials. Magnolol (4-allyl-2-(5-allyl-2-hydroxy-phenyl) phenol) and honokiol (2-(4-hydroxy-3-prop-2-enyl-phenyl)-4-prop-2-enyl-phenol) were obtained from Kishida Chemical Industries, Ltd., Osaka, Japan. Eugenol (4-allyl-2-methoxyphenol) was purchased from Tokyo Kasei Co. (Tokyo, Japan). Bis-eugenol (3,3’-dimethoxy-5,5’-di-2-propenyl-1,1’-biphenyl-2,2’-diol) was synthesized from eugenol monomers by the CuCl(OH)-catalyzed ortho coupling reaction previously described (16). The chemical structures of these eugenol-related compounds are shown in Figure 1. Their solutions were made by dissolving each of them in dimethyl sulfoxide, and then they were diluted to the indicated concentrations using serum-free RPMI-1640 (Invitrogen Co., Carlsbad, CA, USA) as test samples. COX-2 goat polyclonal antibody, β-actin rabbit polyclonal antibody, and horseradish peroxidase (HRP)-conjugated mouse anti-goat IgG were from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Phospho-specific antibody to inhibitory kappa B alpha (IκB-α) (recognizing phospho-Ser32) and anti-IκB-α, both rabbit polyclonal antibodies, as well as HRP-conjugated goat anti-rabbit IgG, and a Phototope-HRP western blot detection kit were obtained from Cell Signaling Technology, Inc. (Beverly, MA, USA). RPMI-1640 was purchased from Invitrogen Corp. Fetal bovine serum (FBS) was from HyClone (Logan, UT, USA). MMA and AIBN were purchased from Tokyo Kasei Co. (Tokyo, Japan).
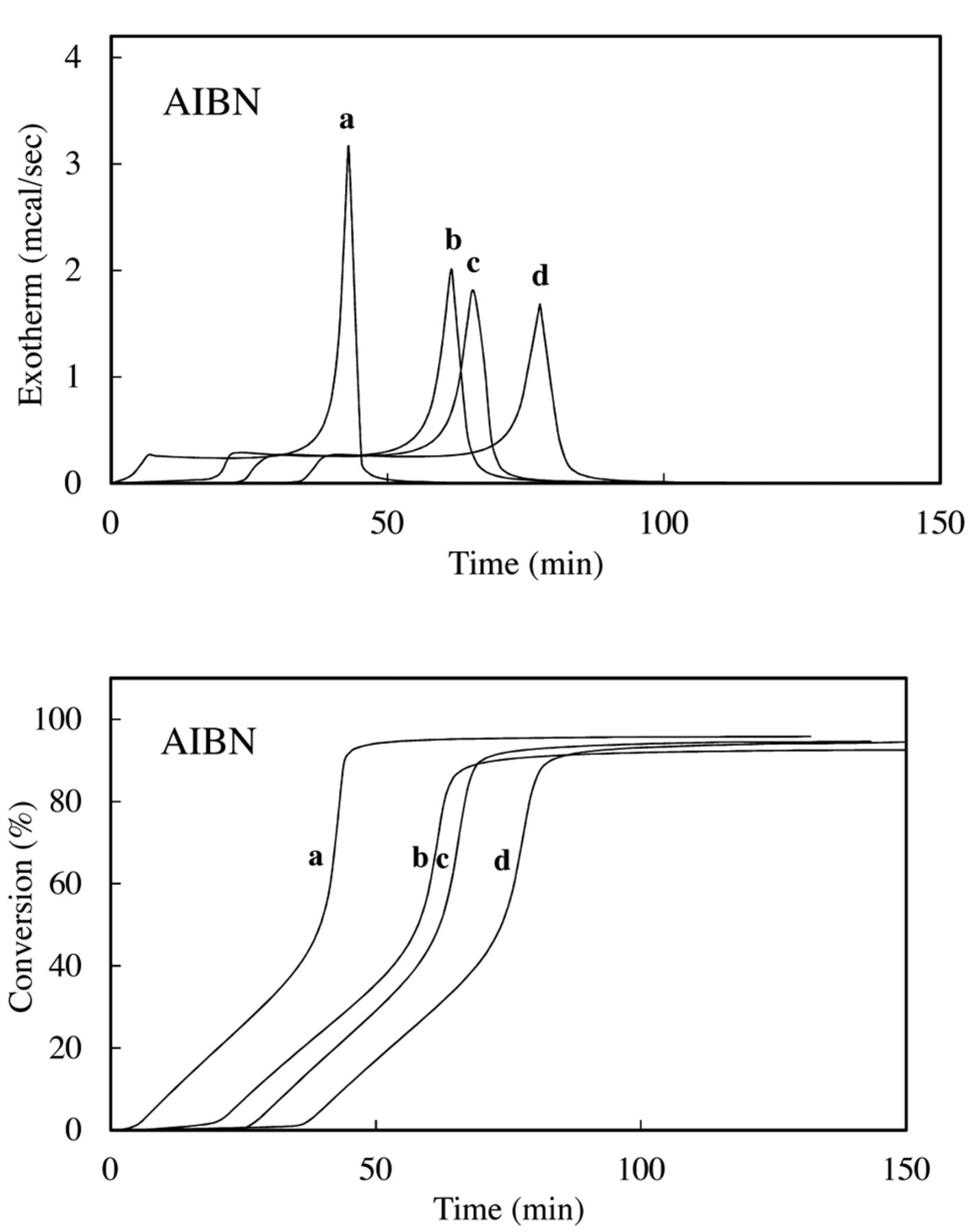
Radical-scavenging activity. The induction period (IP) and initial rate of polymerization in the presence and absence of an antioxidant were determined by the method reported previously (17). In brief, the experimental resin consisted of MMA and AIBN with or without additives. AIBN was added at 1.0 mol%, and the additives were used at 0.05 mol%. Approximately 10 μl of the experimental resin (MMA=9.15-9.30 mg) was loaded into an aluminum sample container and sealed by applying pressure. The container was placed in a differential scanning calorimetry (DSC) (model DSC 3100; Mac Science Co., Tokyo, Japan) kept at 70°C, and the thermal changes induced by polymerization were recorded for the appropriate periods. The heat due to the polymerization of MMA was 13.0 kcal mol−1. The conversion of all samples (%) was calculated from the DSC thermograms using the integrated heat evoked by polymerization of MMA. The value was 93.5-96.7%. Time-exotherm and time-conversion curves are shown in Figure 2. Time-conversion curves break when an inhibitor is consumed. These breaks are sharp and provide a reliable measure of the induction time of the inhibitor. The IP was calculated from the difference between the IP of inhibitors and that of controls.
Measurement of the stoichiometric factor (n). The n value in Equation 1 can be calculated from the IP (time) in the presence of inhibitors:
 (Eq.1)
where IP is the induction period in the presence of an inhibitor (IH). The number of moles of AIBN radicals trapped by the antioxidant was calculated with respect to 1 mole of the inhibitor moiety. The initiation rate (Ri) for AIBN at 70°C was 6.24×10−6 mol l−1s−1, calculated on the basis of n=2.0 for 2,6-di-tert-butyl-4-methoxyphenol.
(Eq.1)
where IP is the induction period in the presence of an inhibitor (IH). The number of moles of AIBN radicals trapped by the antioxidant was calculated with respect to 1 mole of the inhibitor moiety. The initiation rate (Ri) for AIBN at 70°C was 6.24×10−6 mol l−1s−1, calculated on the basis of n=2.0 for 2,6-di-tert-butyl-4-methoxyphenol.
The initiation rate (Ri) for AIBN at 70°C was 6.24×10−6 mol l−1s−1, calculated on the basis of n=2.0 for 2,6-di-tert-butyl-4-methoxyphenol (17).

The chemical structures of eugenol, bis-eugenol, magnolol and honokiol.
Cell culture. The murine macrophage-like cell line RAW264.7, obtained from Dainippon Sumitomo Pharma Biomedical Co. Ltd. (Osaka, Japan), was used. The cells were cultured to a subconfluent state in RPMI-1640 medium supplemented with 10% FBS at 37°C and 5% CO2 in air, washed, and then incubated overnight in serum-free RPMI-1640. They were then washed again and treated with the test samples.
Cytotoxicity. The relative number of viable cells was determined using a Cell Counting Kit-8 (CCK-8) (Dojindo Co., Kumamoto, Japan) (18). In brief, RAW264.7 cells (3×104 per well) were cultured in NUNC 96-well plates (flat-well-type microculture plates) for 48 h, after which the cells were incubated with test samples for 24 h. CCK-8 solution was added to each well and then the absorbance was measured at 450 nm with a microplate reader (Biochromatic, Helsinki, Finland). The 50% cytotoxic concentration (CC50) was determined from the dose–response curves. Data are expressed as means of three independent experiments. Statistical analyses were performed using the Student's t-test.
Preparation of P. gingivalis fimbriae. P. gingivalis ATCC33277 fimbriae were prepared and purified from cell washings by the method of Yoshimura et al. (19). As documented previously, purified fimbria-induced biological activities were not attributable to lipopolysaccharide (LPS) contaminants in the preparation (20, 21). Viability of the cells after exposure to the fimbriae at the concentrations used was over 90% by CCK-8. The protein content of the fimbriae was measured by the method of Smith et al. (22).
Western blot analysis. Cells in 5-cm-diameter Falcon dishes (106 cells per dish) were treated with test samples. The cells were then solubilized with lysis buffer [20 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1 mM EGTA, 1 mM EDTA, 1% (v/v) Triton X-100, 2.5 mM sodium pyrophosphate, 2 mM Na3VO4, 10 mM NaF, 1 mM β-glycerophosphate, 1 μg/ml aprotinin, 1 mM phenylmethylsulfonyl fluoride (PMSF)]. Protein concentrations were measured by the method of Smith et al. (22). Each sample (10 μg of protein) was subjected to sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis (PAGE) in 12.5% polyacrylamide gel, and the separated proteins were transferred to a polyvinylidene difluoride membrane (Millipore Co., Bedford, MA, USA). The blots were then blocked with 5% skim milk, washed and incubated with anti-COX-2 antibody, phospho-specific anti-IκB-α antibody and anti-IκB-α, as the primary antibody, diluted 1:2000 in working solution [5% bovine serum albmim, 1× TBS (50 mM Tris-HCl, pH 7.4, containing 150 mM NaCl) and 0.1% Tween 20] at 4°C. β-Actin antibody was used at 0.1 μg/ml after dilution with working solution. After incubation, the blots were treated at room temperature with HRP-conjugated secondary antibody diluted 1:4000. Proteins were detected with a Phototope-HRP western blot detection kit (Cell Signaling Technology, Inc.), and the blots were exposed to Kodak X-ray film for 10 min. β-Actin was used as a loading control in each lane of the gel.

Exothermic and time-conversion curves for the polymerization of methyl methacrylate (MMA) with azobisisobutyronitrile (AIBN) in the presence of 0.05 mol% additives. Exotherm (left panel) and time-conversion curves (right panel) for the polymerization of MMA with AIBN in the presence of eugenol (b), magnolol (c) and honokiol (d); a: Control.
Preparation of total RNA and real-time polymerase chain reaction (PCR). Cells in NUNC 96-flat-well-type microculture plates (105 cells per well) were treated with test samples. The total RNA was isolated by using an RNeasy Plus Micro Kit (Qiagen Japan Co. Ltd., Tokyo, Japan), in accordance with the instruction manual. cDNA was synthesized from total RNA (2 μg) of each sample by random priming using a High Capacity RNA-to-cDNA Kit (Life Technologies Japan, Tokyo, Japan). Reaction mixtures without the RT were also used as a negative control. An aliquot of each cDNA synthesis reaction mixture was diluted and used for real-time PCR quantification. An equal-volume aliquot of each cDNA was mixed, serially diluted, and used as a standard. TaqMan probes/primers for COX-2 and 18s rRNA and the PCR enzyme mix for real-time PCR were purchased from Life Technologies Japan. The real-time PCR quantification was performed in triplicate using GeneAmp Sequence Detection System 5700 software (Life Technologies Japan) in accordance with the instruction manuals. The relative amount of target was calculated from standad curves generated in each PCR, and quantification data with a co-efficient of variance (CV) less than 10% were used for further analyses. Each calculated mRNA amount was standardized by reference to 18s rRNA.

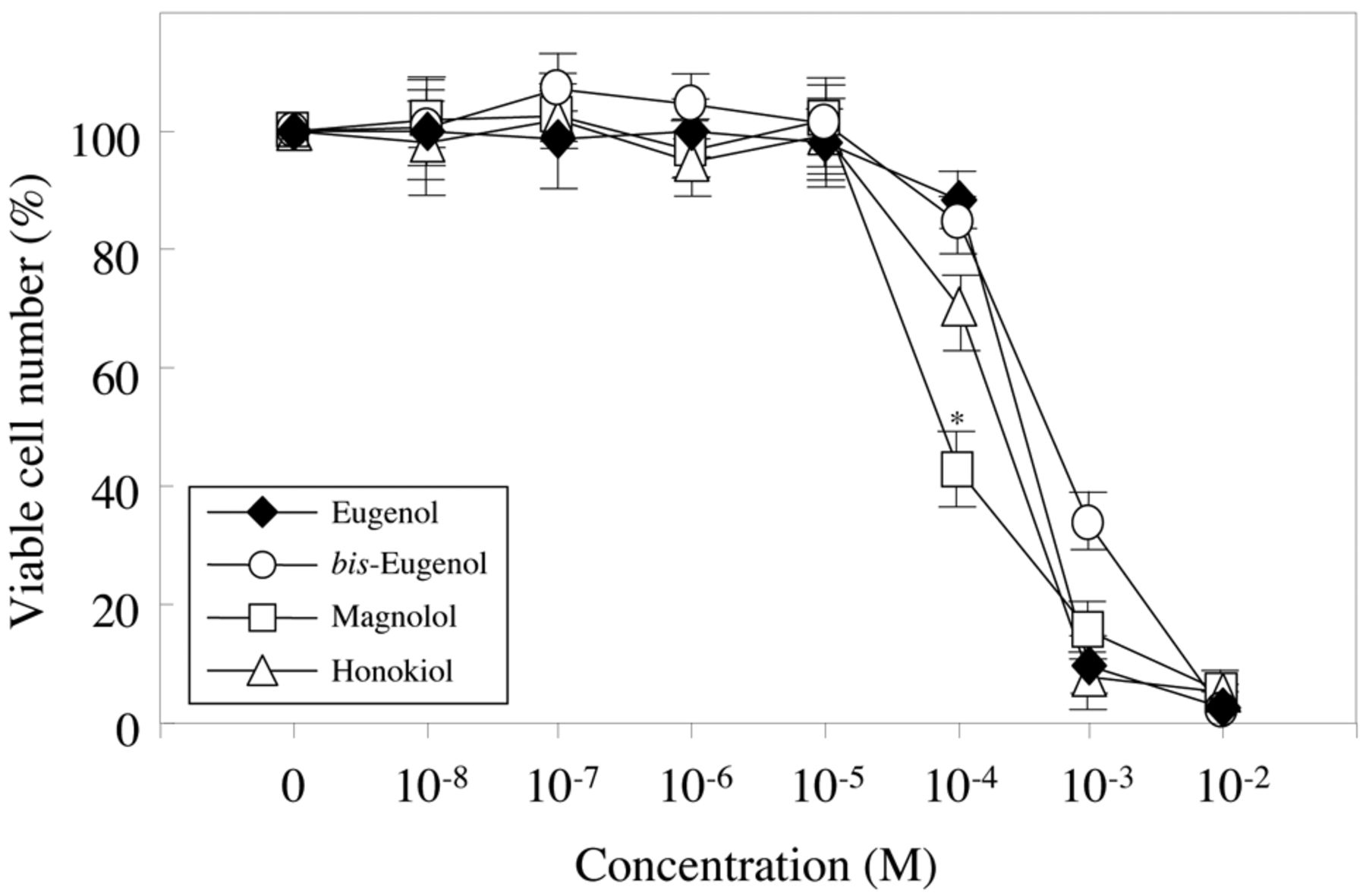
The cytotoxicity of eugenol, bis-eugenol, magnolol and honokiol towards RAW264.7 cells. The procedures employed are described in the Materials and Methods. The results are presented as the means±standard error (SE). SE<15%. There was a significant difference between magnolol and honokiol at 100 μM. *p<0.001.
Preparation of nuclear extract and microwell colorimetric NF-κB assay. Nuclei were extracted and prepared for the microwell colorimetric NF-κB assay. In brief, the cells in 5-cm-diameter Falcon dishes (106 cells per dish) were pretreated for 30 min with or without the indicated doses of the compounds and then treated with fimbriae at 4 μg/ml for 1 h. Thereafter, nuclear extracts were prepared using a Nuclear Extraction kit (Active Motif Co., Carlsbad, CA, USA) in accordance with the manufacturer's protocol. The microwell colorimetric NF-κB assay was performed, as described previously (23), using the Trans-AM NF-κB family transcription factor assay kit (Active Motif Co.). Briefly, cell extracts were incubated in a 96-well plate coated with an oligonucleotide containing the NF-κB consensus binding site (5’-GGGACTTTCC-3’). Activated transcription factors from extracts that bound specifically to the respective immobilized oligonucleotide were detected in an ELISA-like assay using antibodies against the NF-κB p50, p52, p65 and RelB subunits, followed by a secondary antibody conjugated to HRP. Optical density was measured at 450 nm with a microplate reader (Biochromatic). The specificity of the assay was validated by including both the wild-type and mutated oligonucleotides in the reaction. Raji cell nuclear extract was used a positive control.
BDE and orbital energy calculation. BDE was calculated as follows. Firstly, the lowest and second lowest energy conformers of both the phenol derivatives and their phenoxyl radical species were identified as candidates for geometry optimization using a conformer search procedure by Merck Molecular Mechanics force fields (MMFF) calculation. The tentative conformers were them optimized for geometry by the restricted or the unrestricted hybrid Hartree-Fock DFT calculation for the phenols and the phenoxyl radicals in vacuo using B3LYP functional on the 6-31G* basis set level, to afford the energetic minimized structures, respectively. BDE=Hr+Hh−Hp, where Hr is the enthalpy of phenoxyl radical generated by H-abstraction, Hh is the enthalpy of the hydrogen radical, and Hp is the enthalpy of the parent phenol.
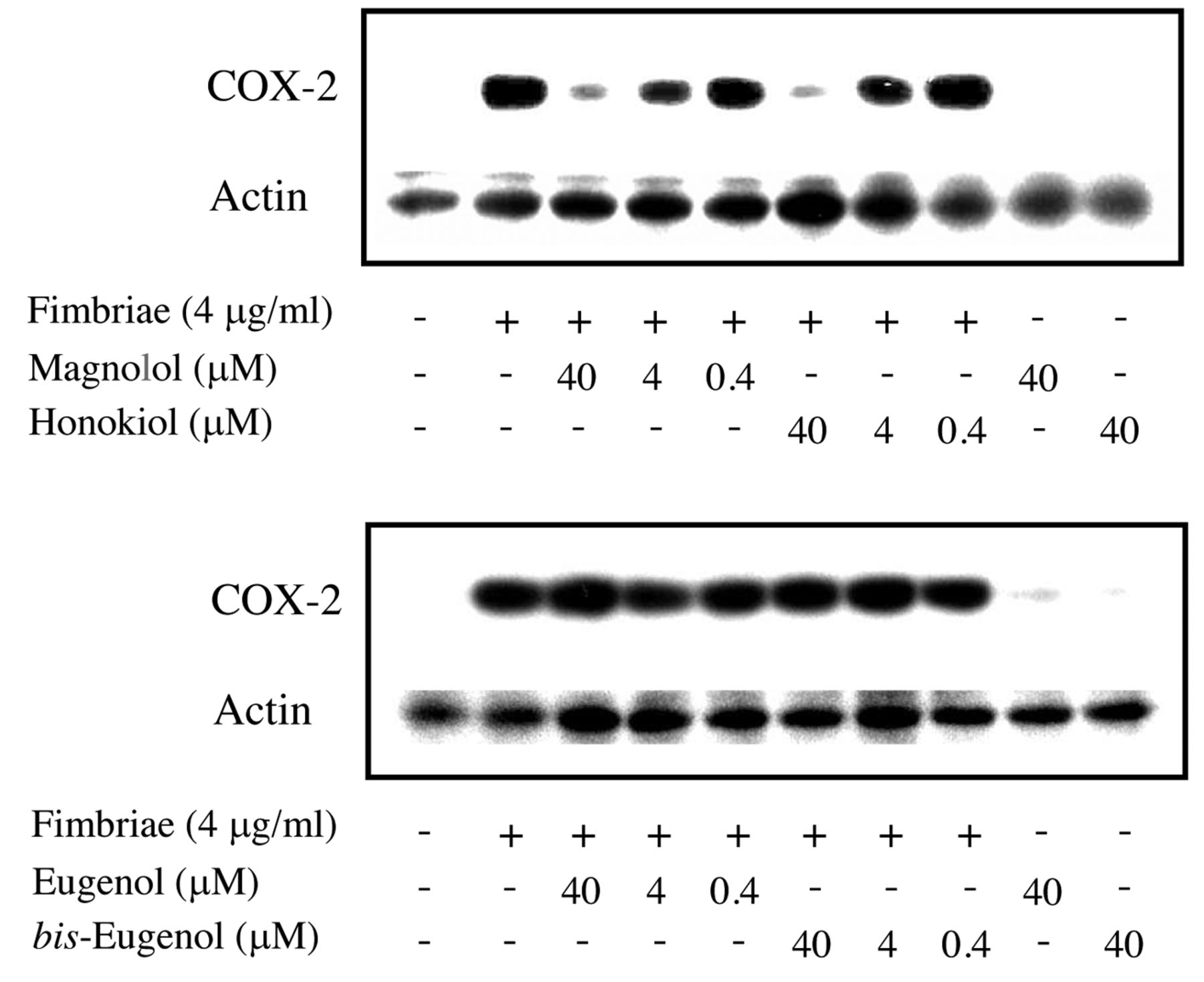
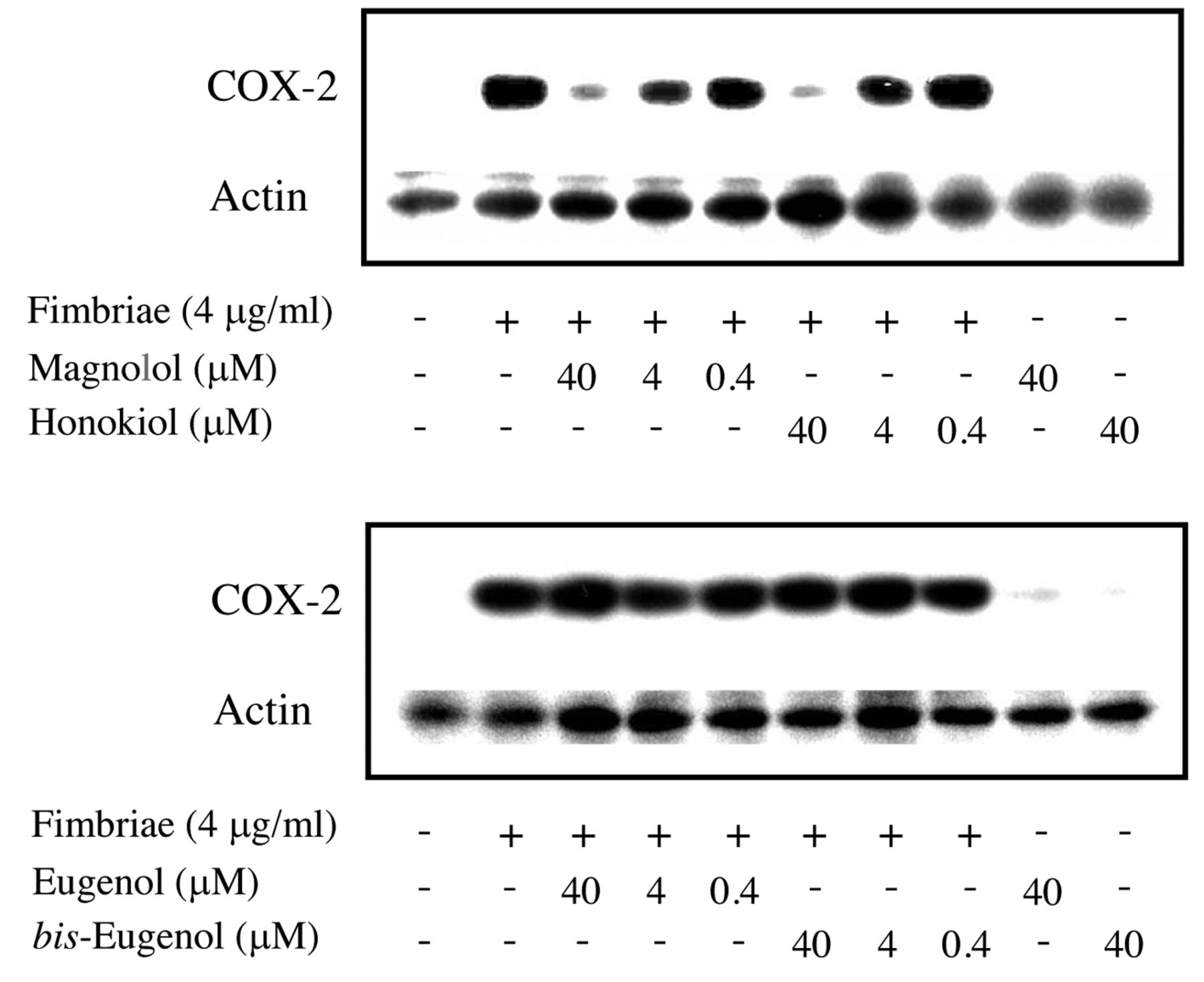
Regulatory effects of magnolol and honokiol on Porphyromonas gingivalis fimbria-stimulated expression on cyclooxygenase (COX)-2 protein in RAW264.7 cells. The cells were pre-treated for 30 min with or without the indicated doses of magnolol or honokiol (upper panel), eugenol or bis-eugenol (lower panel), and then incubated with or without the fimbriae at 4 μg/ml. The cells were solubilized with lysis buffer at 6 h after the start of treatment. Equal amounts of cell lysates were analyzed by western blotting after sodium dodecyl sulfate-polyacrylamide gel electrophoresis with anti-COX-2 antibody. Blotting analysis was performed to determine the levels of COX-2 protein. Three independent experiments were performed, and similar results were obtained.

Regulatory effect of magnolol and honokiol on Porphyromonas gingivalis fimbria-stimulated expression of the cyclooxygenase (COX)-2 gene in RAW264.7 cells. The cells were pre-treated for 30 min with 40 μM eugenol, bis-eugenol, magnolol or honokiol, respectively. They were then incubated for 3 h with or without fimbriae at 4 μg/ml, and their total RNAs were then prepared. Each cDNA was synthesized, and the expression levels of COX-2 mRNA were determined by real-time PCR and standardized against the expression of β-actin mRNA. The results are presented as means±standard error (SE) of three independent experiments. SE<15%. Significant differences between samples for eugenol, bis-eugenol, magnolol and honokiol were observed for inhibition of COX-2 gene expression. *p<0.0001.
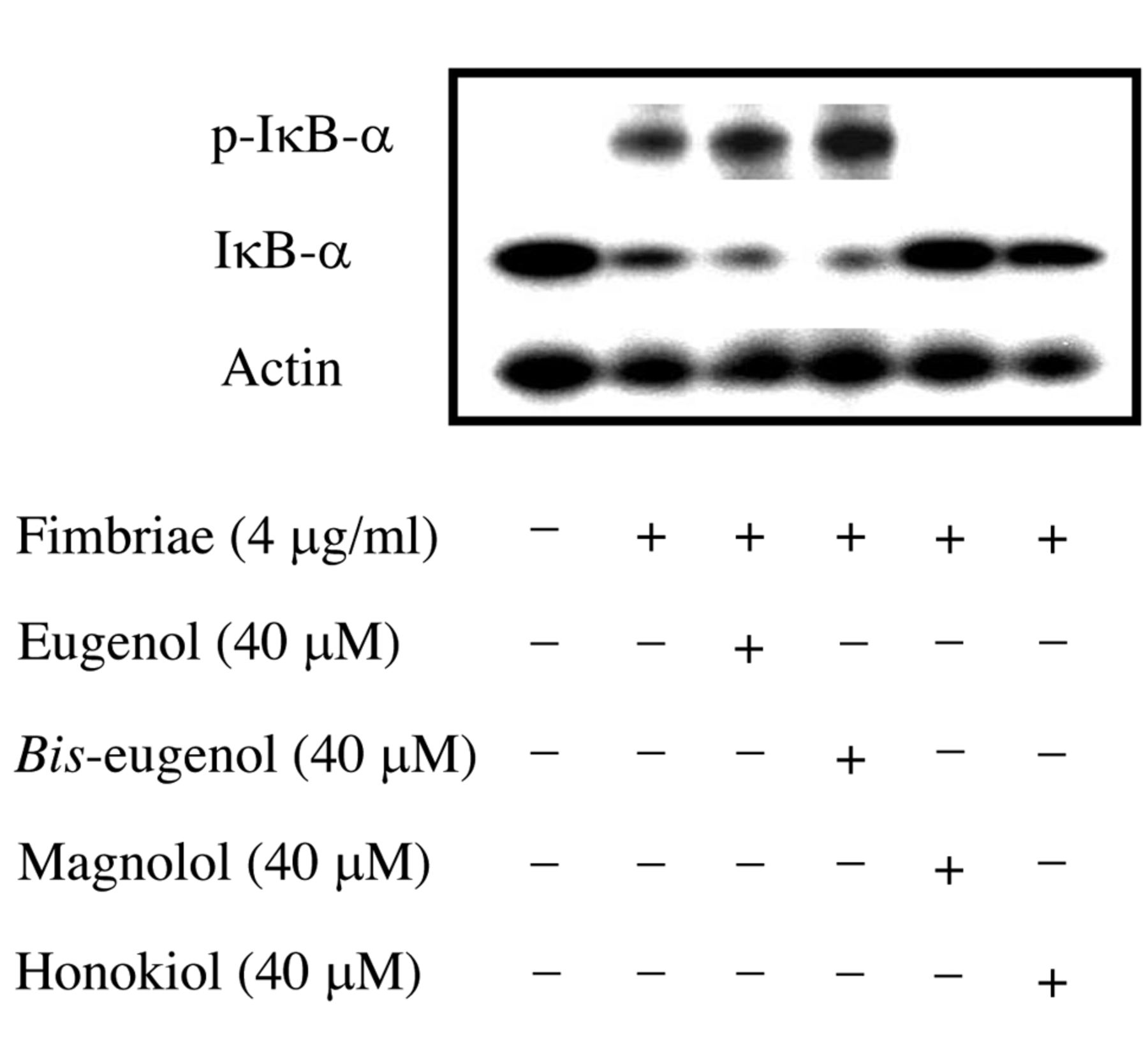
Inhibitory effects of magnolol and honokiol on phosphorylation-dependent proteolysis of Porphyromonas gingivalis fimbria-stimulated inhibitory kappa B alpha (IκB-α). The cells were pre-treated for 30 min with or without eugenol, bis-eugenol, magnolol or honokiol at 40 μM, and then incubated with or without the fimbriae at 4 μg/ml. Equal amounts of cell lysates were analyzed by western blotting after sodium dodecyl sulfate-polyacrylamide gel electrophoresis with phospho-specific anti-IκB-α antibody, anti-IκB-α antibody or anti-β-actin antibody. Three independent experiments were performed, and similar results were obtained.
The energy values of both the highest occupied molecular orbital (HOMO) and the lowest unoccupied molecular orbital (LUMO) energy of the parent phenol derivatives were also obtained from the full optimized geometry described above. The absolute value of HOMO energy was adopted as an approximate ionization potential value according to Koopmans' theory (24).
All of the molecular modeling and calculations were performed with the Spartan 06 for Windows software package (Wavefunction Inc., Irvine, CA, USA).
Chemical hardness (η) and electronegativity (χ) were calculated using Equation 2 and 3, respectively:
 (Eq.2)
(Eq.2)
 (Eq.3)
where ELUMO and EHOMO are the energy levels for the frontier orbitals.
(Eq.3)
where ELUMO and EHOMO are the energy levels for the frontier orbitals.
Results
Radical-scavenging activity. The results are shown in Figure 2. The radical-scavenging activity, i.e. IP (min), declined in the order honokiol (31.51) >bis-eugenol (30.94) > magnolol (21.62) >eugenol (14.72). The stoichiometric factor (n), determined using Equation 1, declined in the order honokiol (2.14), bis-eugenol (2.1) > magnolol (1.47) > eugenol (0.99). Honokiol and bis-eugenol scavenged approximately two radicals. The radical-scavenging activity of magnolol was less than that of honokiol and bis-eugenol. Eugenol scavenged approximately one radical.
Cytotoxicity. The cytotoxicity of eugenol, bis-eugenol, magnolol and honokiol towards RAW264.7 cells was investigated using CCK-8. The results are shown in Figure 3. The (50%) cytotoxic concentration (CC50, μM) for eugenol, bis-eugenol and honokiol and magnolol was 560, 780, 400 and 90, respectively. The cytotoxicity of magnolol was the highest, followed by honokiol. bis-Eugenol exhibited the lowest cytotoxicity.
COX-2 inhibition. The inhibitory effects of magnolol and honokiol on P. gingivalis fimbria-stimulated COX-2 production in RAW264.7 cells were investigated within a concentration range of 0.4-40 μM. The results are shown in Figure 4. Magnolol and honokiol completely inhibited the fimbria-induced production of COX-2 at a non-cytotoxic concentration of 40 μM, whereas eugenol and bis-eugenol did not have an inhibitory effect even at concentrations higher than 40 μM. The latter compounds did not induce COX-2 protein expression in the cells. Furthermore, using real-time PCR, we tested whether magnolol and honokiol inhibited fimbria-induced expression of COX-2 mRNA. The results are shown in Figure 5. Magnolol and honokiol also completely inhibited the expression of COX-2 mRNA, whereas eugenol and bis-eugenol had no effect. These results indicate that magnolol and honokiol exerted a markedly stronger inhibitory effect on COX-2 expression than did eugenol and bis-eugenol.
Inhibition of NF-κB activation. The results are shown in Figure 6. Magnolol and honokiol clearly inhibited both the phosphorylation and degradation of IκB-α in RAW264.7, cells when stimulated by the fimbriae. However, eugenol and bis-eugenol had no effect. The inhibitory effects of magnolol and honokiol on the binding of NF-κB to its consensus sequence in fimbria-stimulated RAW cells were also investigated using an ELISA-like microwell colorimetric transcription factor assay kit. The results are shown in Figure 7. The fimbria-stimulated binding of NF-κB subunits such as p50, p52, p65 and RelB to its consensus sequences were markedly inhibited by magnolol and honokiol, but not by eugenol and bis-eugenol. The fimbria-stimulated binding of NF-κB was also completely inhibited by an excess amount of wild-type consensus oligonucleotide provided in the kit (data not shown). These findings indicate that magnolol and honokiol are potent inhibitors of fimbria-triggered cellular signaling in RAW264.7 cells.

Inhibition of Porphyromonas gingivalis fimbria-stimulated nuclear factor kappa B (NF-κB) activation by magnolol and honokiol in RAW264.7 cells. The cells were pre-treated for 30 min with or without eugenol, bis-eugenol, magnolol or honokiol, and then incubated with or without the fimbriae at 4 μg/ml for 1 h. Nuclear extracts were prepared and used in a TransAM (Active Motif) ELISA-like assay to quantify the NF-κB p50, P65, P52 and RelB DNA-binding activity. The results are presented as means±standard error (SE) of three independent experiments. SE<15%. Significant differences between samples for eugenol, bis-eugenol, magnolol and honokiol were observed for inhibition of NF-κB subunit binding. *p<0.001.
Calculation. The BDE and orbital energy are shown in Table I. BDE (kcal/mol) values declined in the order honokiol (396) >magnolol (392) >eugenol (347) >bis-eugenol (337). The radical decomposition of honokiol appeared to occur preferably at the ortho-position, and not at the para-one, in view of its BDE value. The χ value (eV) declined in the order magnolol (3.2) >honokiol (3.0) >bis-eugenol (2.7) >eugenol (2.0). The χ values for magnolol and honokiol were greater than those for bis-eugenol and eugenol.
Discussion
Magnolol is an isomer of honokiol, differing in the relative arrangement of the hydroxy group to allyl group on the phenolic ring (Figure 1). Eugenol and its dimer, bis-eugenol, are compounds related to magnolol, which possess methoxy groups. The inhibitory effects of magnolol on the induction of nitric oxide synthase (NOS) and COX-2 in RAW264.7 cells induced by LPS have been previously investigated, and it was found that co-treatment with magnolol significantly inhibited LPS-stimulated production of inducible NOS (iNOS) and COX-2 proteins and expression of their genes (25). Magnolol and honokiol have also been reported to exhibit antimicrobial activity against periodontopathic microorganisms and to have a relatively low cytotoxic effect on human gingival cells (26). Magnolol and honokiol may exert preventive effects against chronic diseases caused by infection with oral bacteria. Our results indicated that magnolol and honokiol, but not eugenol and bis-eugenol, significantly inhibited expression of the COX-2 gene induced by exposure to P. gingivalis fimbriae. In addition, magnolol and honokiol inhibited the fimbria-stimulated phosphorylation-dependent proteolysis of IκB-α and the transcriptional activity of NF-κB in the cells. Tse et al. previously reported suppression of NF-κB activation and NF-κB-regulated gene expression through inhibition of I-κB kinase activation (27). The effects of exposure to P. gingivalis fimbriae observed in the present study were similar to those previously reported (25, 27).
We previously reported that bis-eugenol inhibited LPS-stimulated inflammatory cytokine expression at a relatively high concentration (500 μM) but at a low cytotoxic concentration, resulting in weak inhibitory activity of this compound (28). The cytotoxicity of eugenol and bis-eugenol towards RAW 264.7 cells was lower than that of magnolol and honokiol (Figure 2). The fimbriae were found to bind several cellular receptors, such as toll-like receptor-2 and CD11/CD18 (29), suggesting that the COX-2-inhibitory efforts of magnolol and honokiol may affect several signal transduction mechanisms.
Phenolic O-H bond dissociation enthalpy and orbital energy for magnolol and related compounds at DFT B3LYP/6-31G* level.
Redox-sensitive transcription factor NF-κB is implicated in the regulation of many genes that participate in various inflammatory responses. NF-κB is composed of homo- and heterodimeric complexes made up of various dimer combinations of the five subunits of the NF-kB family, p50, p65, c-Rel, p52 and RelB, which serve to activate specific sets of genes, such as these encoding adhesion molecules, immunoreceptors and cytokines (30). The p50/p65 heterodimers and p50/p50 homodimers are the most common dimers found in the NF-κB signaling pathways. Magnolol and honokiol inhibited the fimbria-stimulated binding of four NF-κB subunits to its consensus sequences (Figure 7). Thus, magnolol and honokiol may act as potent inhibitors of fimbria-related diseases through the NF-κB signaling cascade.
The adhesion of P. gingivalis to host cells is likely a prerequisite step in the pathogenesis of P. gingivalis-induced periodontal disease. P. gingivalis binds and invades epithelial cells, and the fimbriae are known to be intrinsically involved in the first step of this process (31). The dynamic systemic pro-inflammatory cellular response to localized periodontal bacteria can occasionally lead to widespread organ damage or death. We have previously demonstrated that fimbriae bind to specific cellular receptors (32), induce the production of pro-inflammatory cytokines (20, 32, 33), and stimulate bone resorption in vitro through signal transduction mechanisms (21, 34). Interestingly, P. gingivalis fimbriae, as well as LPS, can stimulate COX-2 expression in RAW264.7 cells. Magnolol and honokiol scavenge a variety of reactive oxygen and nitrogen species, including the hydroxyl radical, hydrogen peroxide, singlet oxygen, nitric oxide and peroxynitrite anion, and protect against several neurodegenerative diseases and dermatological disorders (7-9). The potent anti-inflammatory activity of magnolol and honokiol may lead to a high degree of endogenous radical-scavenging and anti-inflammatory activity.
Ogata et al. previously investigated the antioxidant activity of honokiol, magnolol and related compounds in vitro and found that their activity declined in the order eugenol>honokiol>magnolol (35). In electron spin resonance spectroscopy (ESR) experiments, they found that eugenol-alone trapped hydroxyl radicals. We also previously investigated the O2−-scavenging activity of eugenol and bis-eugenol using ESR spectroscopy, and found that the scavenging activity of eugenol was greater than that of bis-eugenol (36). These finding suggest that eugenol may be an effective antioxidant acting against O2− and hydroxyl radicals. By contrast, in the present study, the radical-scavenging activity against alkyl radicals declined in the order honokiol, bis-eugenol>magnolol>eugenol. This discrepancy may be related to the oxygen pressure in the experimental systems employed. Our model using the IP method was able to explain the mechanism of radical-scavenging activity and to predict the chain-breaking activity of phenolic compounds, because the measurement by DSC is highly sensitive. Our model was used under nearly aerobic conditions. An oxygen tension of less than 15 Torr is well-known to be similar to that in many tissues. The stoichiometric factor n value of 1 for eugenol suggests that eugenol undergoes dimerization due to its radical-coupling reaction. From this, it has been assumed that eugenol may act as a pro-oxidant in cellular systems via quinone methide intermediates. In cellular systems, eugenol may have a tendency for intermolecular interaction between its methoxy group and its phenolic OH group, which reduces its antioxidative activity. In addition, although bis-eugenol exhibited potent radical-scavenging activity similar to that of honokiol, this compound may also have a tendency for intermolecular interactions between the methoxy group and the phenolic OH group in cellular systems, and consequently this compound may have only weak antioxidant activity due to the decrease in phenolic function at the cellular level. Therefore, the COX-2-inhibitory activity of this compound might have been considerably reduced in comparison with honokiol and magnolol. By contrast, honokiol and magnolol exhibited potent antioxidative activity in cellular systems, resulting in potent COX-2 inhibitory activity. Dikalov et al. investigated the reaction of honokiol in cell-free and cellular systems using ESR and high-performance liquid chromatography (HPLC) techniques and found that it is an effective scavenger of free radicals (37). Honokiol, magnolol, bis-eugenol and eugenol are phenolic compounds with allyl groups in the molecule (Figure 1), but in the present study their biological effects appeared to differ greatly, probably due to the difference in phenolic function induced by their molecular substituents. On the other hand, Zhao and Liu have shown that the radical-scavenging activity of magnolol is lower than that of honokiol (7), similarly to our present findings. Moreover, the lower antioxidant ability of magnolol in comparison with honokiol may be related to the intramolecular hydrogen bond formed between the di-ortho-hydroxyl groups, which interfere with abstraction of the hydrogen atom in the hydroxyl group (7). This finding was supported by our present findings; the n value for honokiol and magnolol was 2.1 and 1.5, respectively. Formation of the intramolecular hydrogen bond may enhance the cytotoxicity of magnolol, which may have a tendency to form quinone methide metabolites in cellular systems. The high cytotoxicity of magnolol may be partly responsible for its reactive quinone methide formation, because of increasing glutathione conjugation of the reactive quinone methide. However, the potent anti-inflammatory activities of honokiol and magnolol were similar in the present study. We previously investigated LPS-stimulated COX-2-inhibitory activity in the presence of 4,4’-biphenol, 2,2’-biphenol and phenol using RAW264.7 cells, and found that 2-2’ biphenol exhibited weak but significant COX-2 inhibition, whereas 4,4’-biphenol and phenol had no activity (38). This indicates that the potent COX-2 inhibition by honokiol and magnolol is probably related to the allyl substituents in their molecules (Figure 1).
We have recently investigated the quantitative structure–activity relationships for various phenolic compounds using chemical hardness (η) and electronegativity (χ) principles, in addition to the BDE value, and found that the mechanisms responsible for the cytotoxicity and COX-2 inhibition of these compounds may be interpreted using computational chemistry (6, 38). Magnolol and honokiol, with potent COX-2 inhibition, had large χ values. The COX-2-inhibitory effects of these compounds may be controlled by their χ values.
In conclusion, magnolol and honokiol, but not eugenol and bis-eugenol, were shown in the present study to significantly inhibit expression of the COX-2 gene, induced by exposure to the fimbriae of P. gingivalis through suppression of NF-κB activation in RAW264.7 cells. These findings suggest that magnolol and honokiol may be capable of preventing periodontopathic bacteria-induced oral diseases and chronic infections.
- Received June 6, 2012.
- Revision received September 13, 2012.
- Accepted September 20, 2012.
- Copyright © 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Radical-scavenging and Pro-/anti-inflammatory Activity of Tetracycline and Related Phenolic Compounds With or Without Visible Light Irradiation
- Cytotoxicity and Pro-inflammatory Properties of Aliphatic Alpha, Beta-unsaturated Acid and Ester Monomers in RAW264.7 Cells and Their Chemical Reactivity
- Cytotoxicity and Pro-/Anti-inflammatory Properties of Cinnamates, Acrylates and Methacrylates Against RAW264.7 Cells
- Anti-inflammatory Activity of {beta}-Carotene, Lycopene and Tri-n-butylborane, a Scavenger of Reactive Oxygen Species
- Expression of Cyclooxygenase-2, Nitric Oxide Synthase 2 and Heme Oxygenase-1 mRNA Induced by Bis-Eugenol in RAW264.7 Cells and their Antioxidant Activity Determined Using the Induction Period Method
- Radical-scavenging and Anti-inflammatory Activity of Quercetin and Related Compounds and Their Combinations Against RAW264.7 Cells Stimulated with Porphyromonas gingivalis Fimbriae. Relationships between Anti-inflammatory Activity and Quantum Chemical Parameters
- The Radical Scavenging Activity and Cytotoxicity of Resveratrol, Orcinol and 4-Allylphenol and their Inhibitory Effects on Cox-2 Gene Expression and Nf-{kappa}b Activation in RAW264.7 Cells Stimulated with Porphyromonas gingivalis-fimbriae
- Anti-inflammatory Activity of the Artificial Antioxidants 2-Tert-butyl-4-methoxyphenol (BHA), 2,6-Di-tert-butyl-4-methylphenol (BHT) and 2,4,6-Tri-tert-butylphenol (TBP), and their Various Combinations
- Inhibitory Effects of p-Cresol and p-Hydroxy Anisole Dimers on Expression of the Cyclooxygenase-2 Gene and Lipopolysaccharide-stimulated Activation of Nuclear Factor-{kappa}B in RAW264.7 Cells
- Inhibitory Effects of Tocopherols on Expression of the Cyclooxygenase-2 Gene in RAW264.7 Cells Stimulated by Lipopolysaccharide, Tumor Necrosis Factor-{alpha} or Porphyromonas gingivalis Fimbriae