


Abstract
Estrogen regulates morphological changes in reproductive organs, such as the vagina and uterus, during the estrous cycles in mice. Estrogen depletion by ovariectomy in adults results in atrophy accompanied by apoptosis in vaginal and uterine cells, while estrogen treatment following ovariectomy elicits cell proliferation in both organs. Sequential changes in mRNA expression of wingless-related MMTV integration site (Wnt) and Notch signaling genes were analyzed in the vagina and uterus of ovariectomized adult mice after a single injection of 17β-estradiol to provide understanding over the molecular basis of differences in response to estrogen in these organs. We found estrogen-dependent up-regulation of Wnt4, Wnt5a and p21 and down-regulation of Wnt11, hairy/enhancer-of-split related with YRPW motif-1 (Hey1) and delta-like 4 (Dll4) in the vagina, and up-regulation of Wnt4, Wnt5a, Hey1, Heyl, Dll1, p21 and p53 and down-regulation of Wnt11, Hey2 and Dll4 in the uterus. The expression of Wnt4, Hey1, Hey2, Heyl, Dll1 and p53 showed different patterns after the estrogen injection. Expression patterns for Wnt5a, Wnt11, Dll4 and p21 in the vagina and uterus were similar, suggesting that these genes are involved in the proliferation of cells in both those organs in mice.
The rodent vaginal epithelium exhibits cyclical changes in response to cyclical ovarian secretions of sex hormones, displaying an alternating pattern of keratinization and mucification (1). The mouse vagina is a good model to study epithelial cell proliferation and keratinization. In ovariectomiced mice, estrogens induce proliferation of the vaginal epithelial cells together with superficial keratinization. In response to estrogens, basal epithelial cells proliferate rapidly, leading to the formation of a highly stratified epithelium (2-4). Estrogens alter cellular physiology by modulating the transcriptional activity of specific nuclear estrogen receptors (ERs) (5), which are believed to stimulate primary response genes, initiating a cascade of transcriptional events, the products of which participate in physiological responses known to be estrogen-dependent events in the target organs in vivo (6). However, the regulation of vaginal gene expression by estrogens and the molecular mechanisms underlying estrogen-mediated cell proliferation remain unclear.
Previously, we reported on expression patterns of mRNAs in the persistently-altered vagina of neonatal mice exposed to diethylstilbestrol (DES) using DNA microarrays and real-time quantitative reverse transcription-polymerase chain reaction (RT-PCR) (7-12). In the vagina exposed neonatally to DES, we found that wingless-related MMTV integration site-4 (Wnt4) was up-regulated without estrogen stimulation and was correlated to the keratinization of the vaginal epithelium (11). Moreover, we identified up-regulation of p21 (also called Waf1, Cap20, Cip1 and Sdi1) and down-regulation of hairy and enhancer of split-1 (Hes1), hairy/enhancer-of-split related with YRPW motif-1 (Hey1) and Heyl, which are target genes of Notch signaling pathway (12) in the neonatally DES-exposed mouse vagina. These genes likely lead to persistent cell proliferation and differentiation in the murine vagina due to neonatal DES exposure.
The estrogen-responsive genes mentioned above in the vagina, identified by gene profiling, provide an important foundation for understanding the functional mechanisms of estrogen regulating morphogenesis and maintenance of the reproductive organs. However, no comprehensive studies have been conducted on mRNA expression of these genes in the vagina and uterus in mice after estrogen exposure. In the present study, we analyzed sequential changes of these gene expressions in the vagina, in comparison to the uterus, of ovariectomized mice given a single injection of 17β-estradiol (E2).
Materials and Methods
Reagents. E2 and sesame oil were obtained from Sigma Chemical Co. (St. Louis, MO, USA) and Kanto Chemical (Tokyo, Japan), respectively.
Animals and treatments. C57BL/6J mice purchased from CLEA Japan (Tokyo, Japan) were maintained under 12 h light/12 h dark at 23-25°C and fed laboratory chow (CE-2; CLEAJapan) and tap water ad libitum. All procedures and protocols were approved by the Institutional Animal Care and Use Committee at the National Institute for Basic Biology, National Institutes of Natural Sciences. C57BL female mice, ovariectomized at 8 weeks of age, were given a single abdominal injection of 50 ng E2/g body weight (bw) dissolved in sesame oil at 10 weeks of age, and sacrificed at 3, 6, 9, 12, 18, 24, 36 or 48 h after the injection, or just before injection (0). Vaginae and uteri of these mice were used for real-time quantitative RT-PCR. Three mice were used for each time point.
In addition, C57BL mice ovariectomized at 8 weeks were given a single abdominal injection of 50 ng E2/g bw (n=3) or the oil vehicle-alone (n=3) at 10 weeks of age. These mice were given a single injection of 50 μg of bromodeoxyuridine (BrdU)/g bw 22 or 46 h after the E2 injection, and were then sacrificed 2 h after the BrdU injection. These mice were used for histology and immunohistochemistry.
BrdU immunostaining. Tissues embedded in paraffin were sectioned at 8 μm and stained with hematoxylin and eosin. In addition, de-paraffinized sections were incubated in 0.3% H2O2 in methanol for 30 min, then immersed in 2 N HCl for 20 min in order to denature the genomic DNA. After washing with phosphate-buffered saline (PBS) in 0.5% Tween, the sections were incubated with anti-BrdU antibody (Boehringer Mannheim, Mannheim, Germany) diluted at 1:20 in PBS containing 1% bovine serum albumin (BSA) (SigmaChemical, Co.) overnight at 4°C. The sections were subsequently incubated with 3,3-diaminobenzidine tetrahydrochloride containing hydrogen peroxide. BrdU-labeling index (percentage) was estimated by counting the number of BrdU-incorporated cells per h in the basal layer of vaginal epithelium as described previously (7, 8).
Real-time quantitative RT-PCR. Total RNA, isolated with RNeasy kit (Qiagen, Chatsworth, CA, USA) from each group of vaginae, was used for real-time quantitative RT-PCR reactions carried-out with SuperScript III reverse transcriptase (Invitrogen, Carlsbad, CA, USA).
Changes in gene expression were confirmed and quantified using the ABI Prism 5700 Sequence Detection System (Applied Biosystems, Foster City, CA, USA) and SYBR Green Master Mix (Applied Biosystems), according to the manufacturer's instructions. PCR conditions were as follows: 50°C for 2 min, 95°C for 10 min, and 36 cycles of 95°C for 15 s and 60°C for 1 min in 15 μl volumes. Relative RNA equivalents for each sample were obtained by standardization of L8 ribosomal protein levels. Sequences of gene primer sets are given in Table I. Three pools of samples per group were run in triplicate to determine sample reproducibility, and the average relative RNA equivalents per sample were used for further analysis. Error bars represent the standard error, with all values represented as fold-change compared to the control treatment group normalized to an average of 1.0.
Immunohistochemistry. Tissues were fixed in 10% neutral-buffered formalin, embedded in paraffin and sectioned at 4 μm. The sections were de-paraffinized and incubated with 0.3% H2O2 in methanol for 15 min to eliminate endogenous peroxidase. After washing with PBS, the sections were stained with LSAB™ 2 kit, Universal (Dako, Carpinteria, CA, USA), according to the manufacturer-supplied protocol. Polyclonal antibody against Wnt4 was obtained from R&D Systems, Inc. (Minneapolis, MN, USA). The tissue sections were incubated at 1:200 dilution in PBS, containing 1% BSA overnight at 4°C. For negative controls, a normal goat immunoglobulin (IgG) fraction (Dako) was used at the same dilution. Monoclonal antibody against p21 was obtained from Santa Cruz, Inc. (Minneapolis, MN, USA). The sections were incubated at 1:50 dilution in PBS containing 1% BSA, overnight at 4°C. For negative controls, murine IgG2b (Dako) was used at the same dilution.
Statistical analysis. Statistical analyses were performed using one-way analysis of variance (ANOVA). Differences of p<0.05 were considered significant.
Results
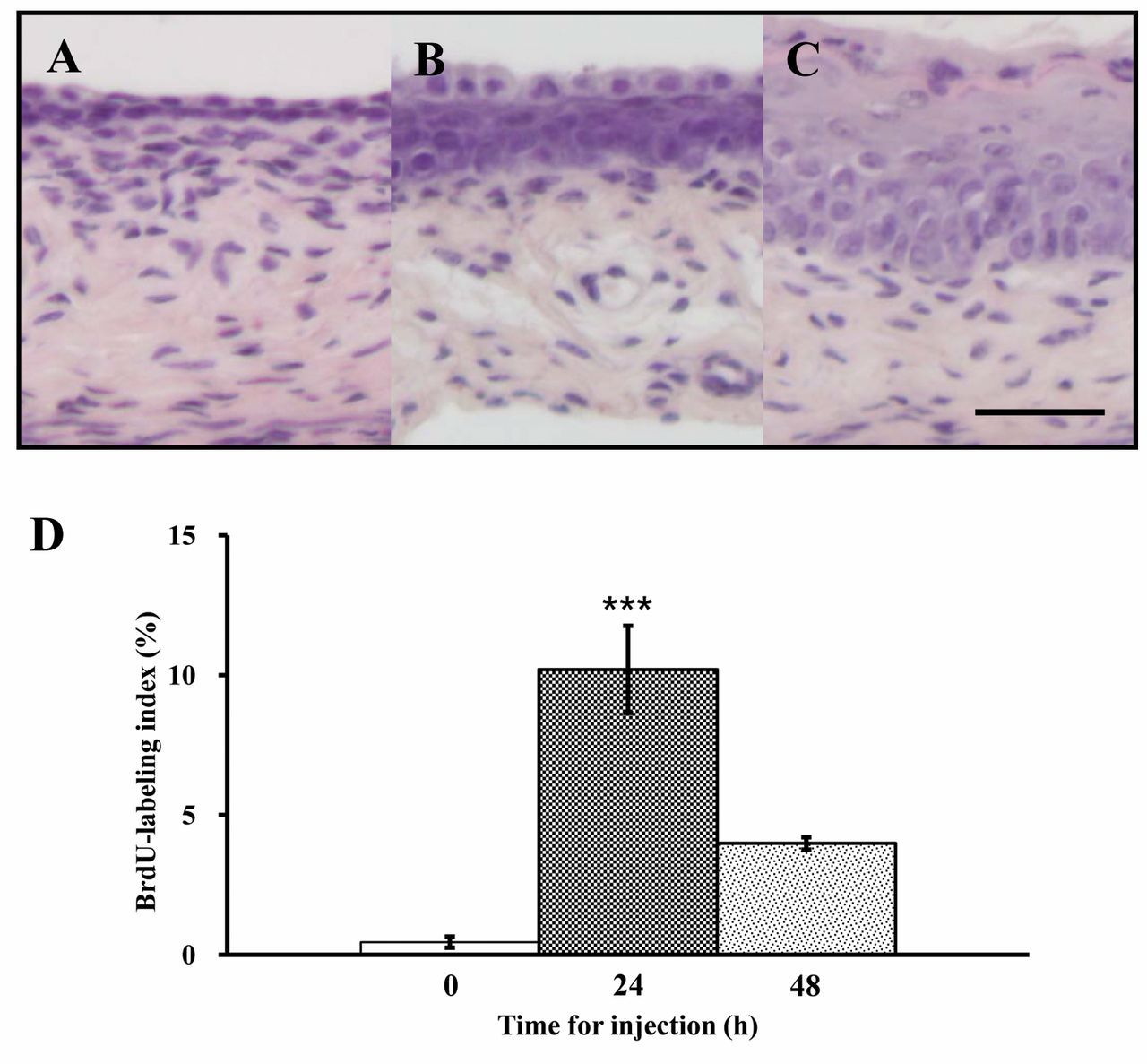
Morphological changes and differential expression of cell cycle-related genes in vaginae of ovariectomized mice given a single injection of E2. Histological changes in the vagina following E2 injection are shown in Figures 1A-C. The epithelium exhibited proliferation at 24 h (Figure 1B), and underwent stratification with superficial keratinization at 48 h after the injection (Figure 1C). The basal cells of vaginal epithelium at 24 h after the injection exhibited a high proliferative activity (10.2%), confirmed by BrdU immunostaining (Figure 1D). At 48 h after the injection, proliferative activity declined to 4.0% (Figure 1D).
Since cyclin A2 is expressed from S phase to M phase during cell division, sequential changes in gene expression levels of cyclin A2 in addition to keratin 1 were examined after E2 treatment. The gene expression of cyclin A2 was increased 18-24 h after E2 treatment (Figure 2A), and that of keratin 1 had a similar pattern to that of cyclin A2 (Figure 2B). Up-regulation of cyclin A2 and keratin 1 was correlated with epithelial stratification and keratinization in the vagina.
Sequences of gene primer sets for real-time quantitative RT-PCR.
Localization of Wnt4 and p21 proteins in the mouse vagina. The localization of Wnt4 and p21 protein in the proliferating epithelial cells of vaginae was examined by immunohistochemistry (IHC). Wnt4 staining was observed in the basal and middle layers of vaginal epithelium 24 h after the E2 injection (Figure 3A). p21 was localized in the basal layer of vaginal epithelium in E2-treated mice (Figure 3C). No response was visible in the vaginal epithelium stained with normal goat immunoglobulin (IgG) as a negative control (Figure 3B and D). Hence, th Wnt4 and p21 proteins were evident in the proliferating vaginal epithelial cells.
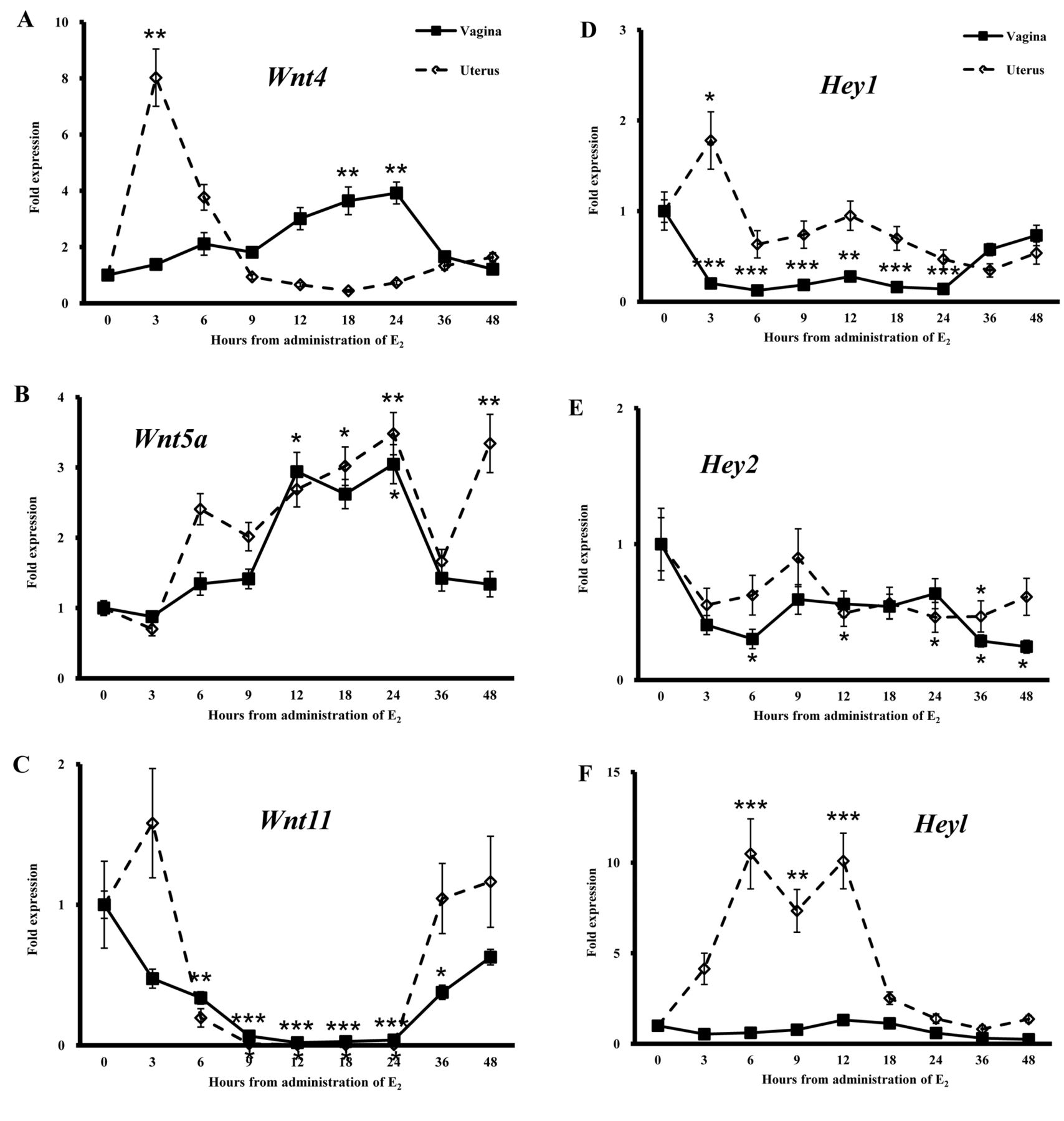
Differential expression of Wnt genes and Notch signal pathway-related genes in the vagina and uterus of ovariectomized mice given a single injection of E2. On the basis of the morphological findings in the vagina of ovariectomized mice given E2, we compared the expression patterns of Wnt genes and Notch signal pathway-related genes in the vagina and uterus after treatment with E2. In the vagina, expression of Wnt4 was increased at 18 and 24 h after the E2 injection, while in the uterus, the expression was transiently increased only at 3 h (Figure 4A). Expression of Wnt5a was increased at 12 and 24 h after E2 injection in the vagina and at 18, 24 and 36 h in the uterus, respectively (Figure 4B). Expression of Wnt11 was decreased at 6-36 h after E2 administration in the vagina and at 9-24 h in the uterus, respectively (Figure 4C). The expression of Wnt4 was different between the vagina and uterus, while the pattern for Wnt5a and Wnt11 revealed a similar trend in both organs.
The expression of Hey1 was decreased at 3-24 h after the E2 injection in the vagina. On the other hand, the expression increased transiently 3 h in the uterus (Figure 4D). Expression of Hey2 was decreased at 12, 24 and 36 h in the uterus, while in the vagina, the expression was decreased at 6, 36 and 48 h (Figure 4E). Expression of Heyl was increased 6-12 h after the administration only in the uterus (Figure 4E and F). However, the expression showed no change in the vagina after E2 injection (Figure 4F). The expression of Dll1 was increased in the uterus 6 h after administration, while the expression remained unchanged in the vagina (Figure 4G). The expression of Dll4 was decreased in the vagina at 3, 9, 18-48 h after E2 administration and in the uterus at 3-24 h (Figure 4H).
The expression of p21 and p53 was increased at 6-12 h after E2 administration in the uterus (Figure 4I and J). In the vagina, however, E2 injection only increased p21 expression at 12 h, with no effect on the expression of p53 (Figure 4I and J).
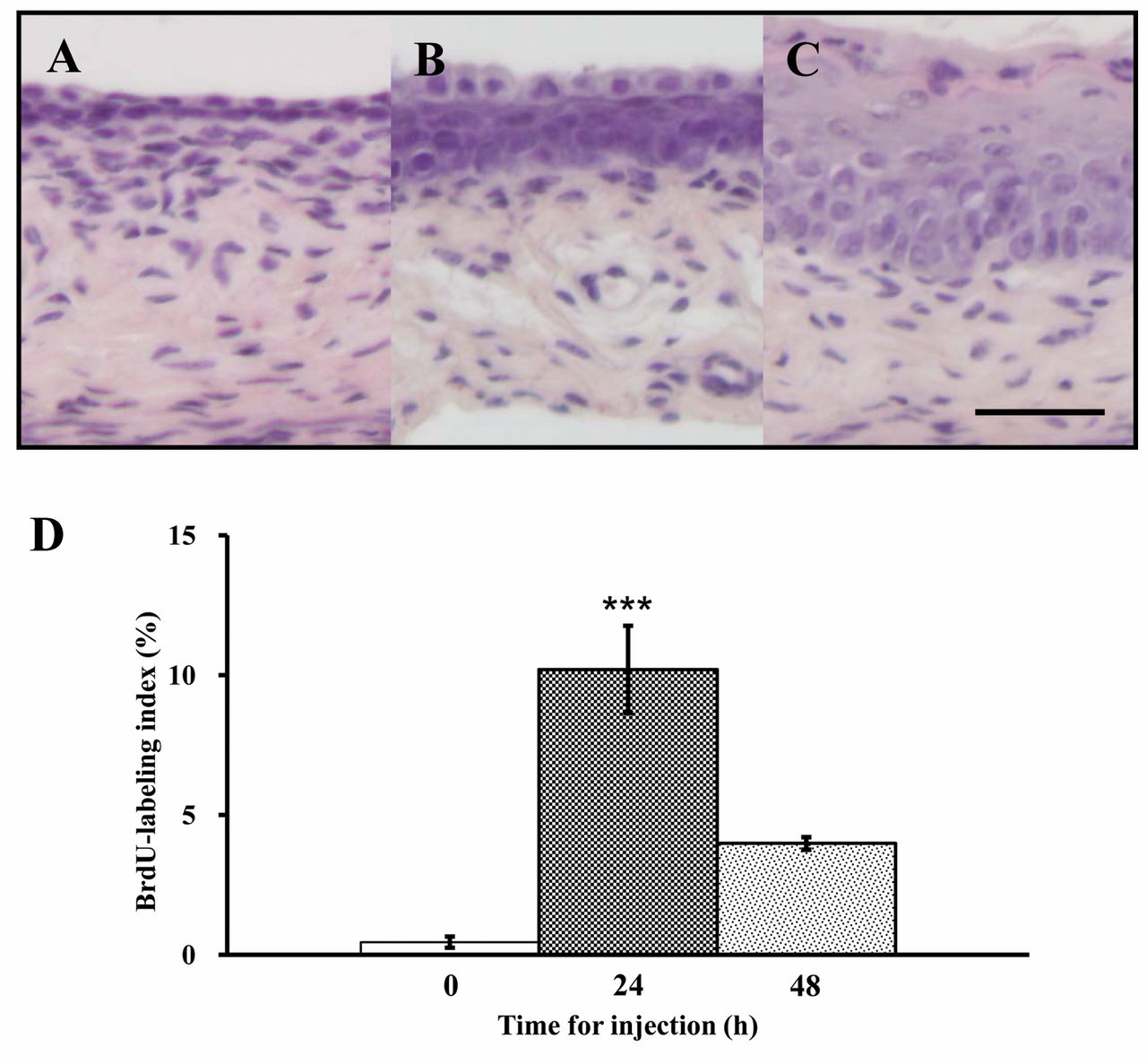
Histology of vagina of a 10-week-old mouse, ovariectomized at 8 weeks of age, and killed just before (A) or 24 h (B) and 48 h (C) after a single abdominal injection of 50 ng 17β-estradiol (E2)/g bw dissolved in sesame oil at 10 weeks of the injection. Bar=50 μm. Incidence (%) of BrdU-incorporated basal cells in the vaginal epithelium of mice sacrificed before injection, and 24 and 48 h after a single E2 injection (D). Data are the means±standard error (n=3). ***p<0.001 vs. controls (one-way ANOVA).
Discussion
Estrogens induce epithelial cell proliferation and differentiation, whereas estrogen depletion results in atrophy accompanied by apoptosis in adult female mouse reproductive organs, such as the vagina and uterus (2-4). In order to understand the underlying mechanisms of estrogen functions in reproductive organs in mice, we have analyzed estrogen-responsive genes in reproductive organs, and found that expression patterns of genes are different between the vagina and uterus (9, 13, 14). Previously, we reported on the expression of Wnt- and Notch-related genes in the vagina and uterus exposed-neonatally to DES (9-12), and the localization of Wnt4 or p21 proteins and quantitation of BrdU-positive cells after E2 administration in the ovariectomized mouse uterus (15, 16). Regarding the vagina, however, detailed information has not been reported. Therefore, we investigated gene expression of Wnt- and Notch-related genes in the vagina and uterus after a single injection of E2 in mice.
Sassoon's group (13, 17, 18) reported that Wnt4, Wnt7a, and Wnt5a are required for Müllerian duct formation, subsequent differentiation, and posterior growth, respectively, and the expression of these genes in the uterus changes during the estrous cycle (17). These findings suggest important roles of Wnt family genes in various reproductive physiologies. In the present study, Wnt4, but not Wnt5a and Wnt11, showed different expression pattern in the vagina and uterus.
Cyclin A2 is expressed from S phase to M phase, so the cell cycle phase after E2 treatment can be determined by analyzing changes in cyclin expression (19). In the present study, expression of cyclin A2 and keratin 1 was increased 24 h after a single injection of E2 in the vagina, similarly to the BrdU labeling index and the expression of Wnt4. These results suggest that Wnt4 might act on epithelial stratification and keratinization in the vagina. In the uterus, Wnt4 transiently increased 3 h after E2 treatment; Wnt4 is expressed in the uterine epithelium at pro-estrus at the time of highest estrogen levels (17, 20), suggesting that estrogen regulates Wnt4 expression in the uterus. On the other hand, Wnt5a and Wnt11 had similar expression patterns in the vagina and uterus. Wnt5a expression has been reported to change in both organs during the estrous cycle (17), and it is expressed at higher levels at pro-estrus in the vaginal epithelium and stroma (17), suggesting involvement in cell proliferation and keratinization in the vagina. We confirmed the similarity of the expression patterns among Wnt5a, cyclin A2 and keratin 1 in the vagina in the present study. In the uterus, Wnt5a has been detected in both the epithelium and the stroma at estrus, but only in the uterine stroma at pro-estrous (17). Zhu et al. (21) reported that the expression of Wnt11 is inhibited by androgens. The present data indicate that Wnt11 is down-regulated in both organs by E2 treatment, showing negative correlation with cell proliferation in both organs, as in the vagina of neonatally DES-exposed mice (11).

Fold expression of cyclin A2 (A) and keratin 1 (B) after a single injection of 50 ng 17β-estradiol (E2)/g bw. Expression of these genes in control mice was regarded as the basal level (1.0). Data are the means±standard error (n=3). *p<0.05 and ***p<0.001 vs. controls (one-way ANOVA).

Immunohistochemistry by wingless-related MMTV integration site-4 (Wnt4) (A) and p21 (C). Vaginae 10-week-old, ovariectomized mice were killed 24 h after a single injection of 17β-estradiol (E2). For negative controls, sections were incubated with pre-immune serum instead of primary antibody (B, D). Bar=50 μm. The boundary between the epithelium and the stroma is indicated by a dotted line.
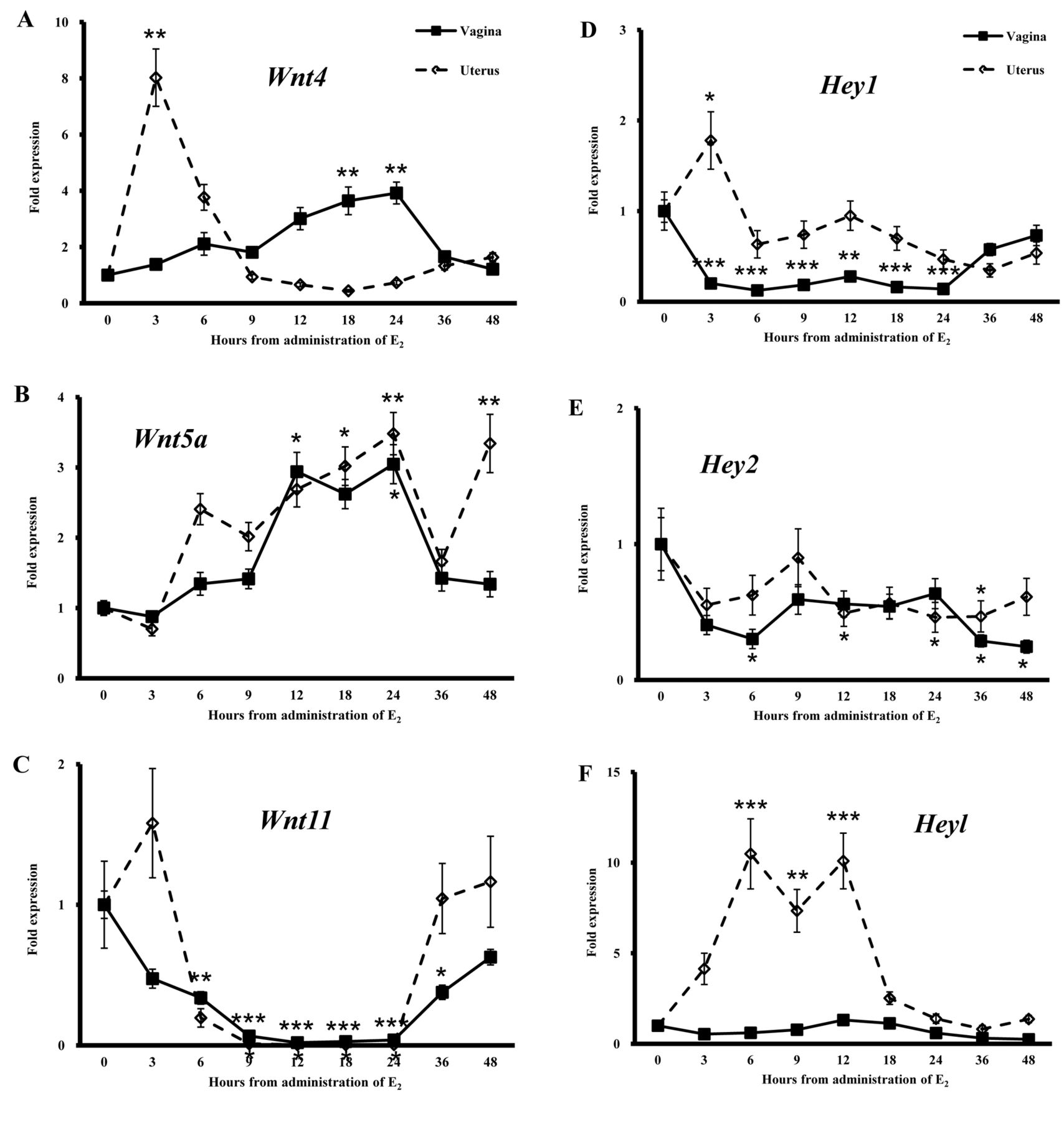

Fold expression of wingless-related MMTV integration site-4 (Wnt4) (A), Wnt5a (B), Wnt 11 (C), hairy/enhancer-of-split related with YRPW motif-1 (Hey1) (D), Hey2 (E), Heyl (F), delta-like-1 (Dll1) (G), Dll4 (H), p21 (I) and p53 (J) in the vagina and uterus after a single injection of 50 ng 17β-estradiol (E2)/g bw. Expression of these genes in control mice before E2 injection (0 h) was regarded as the basal level (1.0). Data are the means±standard error (n=3). *p<0.05, **p<0.01 and ***p<0.001 vs. controls (one-way ANOVA).
Previously, we reported that Hey1 and Heyl, Notch target genes, are persistently down-regulated in the vagina exhibiting estrogen-independent epithelial cell proliferation in the neonatally DES-exposed mice (12). In this study, Hey1, Hey2 and Heyl had differential expression patterns between vagina and uterus in response to E2 stimulation. Hey1 was down-regulated at 3-24 h after E2 treatment in the vagina, suggesting the opposite behavior in proliferation of vaginal cells to that of uterine cells. Hey1 and Heyl may have roles in maintaining undifferentiated quiescent cells, as in satellite muscle cells (22).
We reported that Notch ligand genes, Dll1 and Dll4, are unaltered in the vagina exhibiting estrogen-independent epithelial cell proliferation in the neonatally DES-exposed mice (12). Dll1 and Dll4 genes had a differential expression pattern between the vagina and uterus after E2 treatment in this study. Expression of Dll1 was transiently increased 6 h after E2 stimulation but it showed no change in the vagina. Dll4 is reported to be involved in tumor angiogenesis (23, 24), and is expressed in the arterial endothelium (25). Dll4, as well as Wnt11, exhibited down-regulation by E2 in both organs, suggesting a negative correlation with cell proliferation.
Finally, we confirmed the expression of p21 and p53. p21 is a direct target of p53 tumor suppressor, and mediates p53-dependent cell cycle arrest in response to DNA damage (26). In this study, we found the expression patterns of p21 and p53 in the uterus to be similar.
In conclusion, we demonstrated the difference in the expression patterns of Wnt4, Wnt5a, Wnt11, Hey1, Hey2, Heyl, Dll1, Dll4, p21 and p53 between the vagina and uterus after E2 stimulation. Moreover, we identified the localization of Wnt4 and p21 proteins in the ovariectmized adult mouse vagina exhibiting epithelial stratification, after a single injection of E2. Additional elucidation of the molecular mechanism of cell proliferation in the vagina and uterus is essential.
Acknowledgements
We thank Dr. Raphael Guzman, Department of Molecular and Cell Biology and the Cancer Research Laboratory, University of California, Berkeley, CA, USA, for his critical reading of the manuscript. This work was supported by Grants-in-Aid from the Ministry of Education, Culture, Sport, Science and Technology (S.M., T.S., T.I., Y.O.) and a Grant from the Ministry of Health Labor and Welfare (T.I.), Japan.
- Received July 25, 2012.
- Revision received October 10, 2012.
- Accepted October 11, 2012.
- Copyright © 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}