


Abstract
Background/Aim: To evaluate the impact of an antibiotic, minocycline, on several immune parameters in response to radiation in a mouse model. Materials and Methods: C57BL/6 mice were treated with minocycline (i.p.) for 5 days, beginning immediately before radiation with 1-3 Gy 60Co γ-rays. Spleen and blood were collected on day 4 post-irradiation. Cell populations were determined in the blood and spleen. Splenocytes were activated with anti-CD3 antibody for 48 h and cytokines were quantified. Results: Minocycline increased the counts and/or percentages of splenic macrophages, granulocytes, natural killer, T- and CD8+ T-cells (p<0.05 versus radiation alone). Minocycline significantly increased the expression of interleukin-1α and β, which are radioprotective, as well as the ones of granulocyte-macrophage colony-stimulating factor and granulocyte colony-stimulating factor, which accelerate neutrophil recovery (p<0.05 versus radiation alone), while suppressing cytokines that could prevent hematopoiesis, e.g. macrophage inflammatory protein-1α, tumor necrosis factor-α and interferon-γ. Conclusion: These data indicate that minocycline should be further tested for use in restoration of the hematopoietic system after radiation exposure.
Radiation exposure is a highly significant concern in the event of nuclear disasters. Although cytokines and growth factors have been extensively researched as potential radioprotectors and radiomitigators (1), very few effective countermeasures are available. Consequently, protection against radiation damage to normal tissues is very important to the Departments of Defense, Energy and Homeland Security, as well as the National Aeronautics and Space Administration of the United States of America. Radiation in the space environment is above levels deemed to be safe on Earth and crew members could be exposed to doses as high as 1-3 Gy during solar particle events (2). In addition, radiation is widely employed as a treatment option for a wide range of malignancies. Bone marrow transplant recipients also receive doses of radiation that can result in hematopoietic syndrome. Despite advancements in treatment strategies, toxicity against normal tissues still remains a cause for concern.
Exposure to ionizing radiation can cause a variety of physiological changes collectively known as acute radiation syndrome (ARS). One of the major factors in mortality in patients with ARS is damage to the hematopoietic system. Exposure to doses ~0.5-1 Gy or above (gamma or X-rays) can compromise the hematopoietic system to some extent (3). Whole-body irradiation at doses ranging from approximately 1 to 8 Gy can lead to hematopoietic syndrome, characterized by a dramatic decrease in peripheral white blood cell (WBC), platelet and red blood cell (RBC) counts and, if accompanied by the lack of prompt medical intervention that includes bone marrow transplantation, it can rapidly lead to death (3). Hematopoietic progenitor stem cells have a restricted capacity for cell division after total-body irradiation of more than 2 to 3 Gy (4). Numerous complications are possible, including infections and internal hemorrhage. Medical management of ARS often includes administration of broad-spectrum antibiotics (5), as well as blood products and colony-stimulating factors to regenerate granulocytes. Although there are currently no drugs approved by the Food and Drug Administration (FDA) for treatment of ARS, the use of growth factors such as granulocyte colony-stimulating factor (G-CSF) and granulocyte-macrophage colony-stimulating factor (GM-CSF), is included as part of the standard therapy for ARS in the United States, resulting in improved function and numbers of macrophages and granulocytes (6). Daily injections may be needed and side-effects can occur. In more severe cases, bone marrow transplantation is a treatment option. However, since this is not feasible in emergencies involving a large number of people, other radiation countermeasures are required.
Antibiotics have been employed as an important component in treating radiation injuries to minimize risk for systemic infections and facilitate recovery of tissues, such as bone marrow and the gastrointestinal tract (7). Minocycline, a semisynthetic, second-generation tetracycline derivative, is often used to treat infections due to Neisseria meningitidis and multidrug-resistant Staphylococcus aureus. Recent studies have demonstrated that minocycline possesses other potentially therapeutic effects that are distinct from its antimicrobial action. It is reported to be neuroprotective and antiapoptotic (8). Tikka et al. demonstrated radioprotection of neuronal cells through minocycline by the reduction in the level of lactate dehydrogenase, DNA fragmentation and microglial cell numbers (9). Krauss et al. found that minocycline, unlike tetracycline, possesses antioxidant and free radical scavenging properties, similar to vitamin E (10). Tetracyclines have been reported to have radioprotective properties for hematopoietic cells by Kim et al. (5). In this context, we hypothesized that minocycline has the potential to facilitate regeneration of immune cell populations following whole-body irradiation. We further hypothesized that minocycline would enhance the capacity to secrete cytokines that facilitate hematopoiesis, thus minimizing the potential for serious complications associated with ARS. Day 4 after irradiation was selected for the analyses because ARS can result in death within a few days. Thus, an agent that protects close to the time of irradiation and (or) rapidly enhances hematopoiesis would be of considerable value. In addition, immune depression at the day 4 time point is close to the nadir after 1-3 Gy whole-body irradiation in the C57BL/6 mouse model used in the present study.
Materials and Methods
Animals and experimental design. Female C57BL/6 mice (n=80; 8-9 weeks of age; Charles River Breeding Laboratories, Inc. Hollister, CA, USA) were housed in large plastic cages (n=10/cage) and acclimatized for 5-7 days under standard vivarium conditions. Animals were divided into eight treatment groups (10 mice/group): (a) deionized water (dH2O), no irradiation (b) dH2O plus 1 Gy; (c) dH2O plus 2 Gy; (d) dH2O plus 3 Gy; (e) minocycline plus 0 Gy; (f) minocycline plus 1 Gy; (g) minocycline plus 2 Gy; and (h) minocycline plus 3 Gy. To characterize the early effects of the drug, animals were rapidly euthanized on day 4 post-irradiation using 100% CO2 in compliance with the recommendations of the National Institutes of Health and the Panel of Euthanasia of the American Veterinary Medical Association. The protocol used was approved by the Loma Linda University Institutional Animal Care and Use Committee.
Drug treatment and irradiation. Minocycline was purchased from Triax Pharmaceuticals, LLC, Cranford, NJ, USA. Animals in the respective groups were given an intraperitoneal (i.p.) injection of minocycline hydrochloride (45 mg/kg in a volume of 0.1 ml) immediately before irradiation or i.p. injections of dH2O. The dose of drug and timing of the injections was based on previous reports (11-13). An Eldorado unit containing a Co-60 source was used to administer whole-body radiation at a dose rate of 1.58 Gy/min for a total dose of 1, 2 or 3 Gy to the mice in the respective groups. Non-anesthetized mice were placed individually into rectangular plastic aerated boxes (30×30×60 mm3). A second injection of minocycline (45 mg/kg) or dH2O was administered to the appropriate groups immediately after irradiation. This was followed by three consecutive injections of minocycline (22.5 mg/kg) or dH2O on the following three days post-irradiation. Similar treatment was given to sham-irradiated groups.
Blood and spleen collection. Blood was collected by cardiac puncture immediately after euthanasia in 1 ml tuberculin syringes containing [K2]EDTA. Spleens were harvested and single-celled suspensions were prepared by processing the spleens in complete RPMI-1640 medium (Irvine Scientific, Santa Ana, CA, USA) using sterile applicator sticks. Spleens were washed and centrifuged to remove debris followed by lysis of RBC using 2 ml lysis buffer for 4 min at 4°C. Splenic leukocytes were suspended in 2 ml of RPMI-1640 medium for further analysis.
Relative spleen mass (RSM). Body weight was recorded for each mouse at the time of euthanasia and the spleens were weighed immediately after excision. Relative spleen mass was calculated as follows: RSM=spleen mass (mg)/body mass (g).
Analysis of cell populations in spleen and blood. Spleen and blood cells were analyzed on an automated analyzer (HESKA™ Vet ABC-Diff Hematology Analyzer: HESKA Corp., Waukesha, WI, USA). WBC counts, as well as numbers and percentages of lymphocytes, granulocytes, and monocytes/macrophages were obtained. For the blood, additional values were obtained as follows: RBC count, hemoglobin (HGB), hematocrit (HCT), mean corpuscular hemoglobin (MCH), MCHC: mean corpuscular hemoglobin concentration; MCV: mean corpuscular volume; RDW: RBC distribution width; PLT: platelet count; MPV: mean platelet volume; and EOS: eosinophils.
Flow cytometric analysis of lymphocyte populations in spleen. Specific lymphocyte populations in the spleen were determined using monoclonal antibody (mAb) mixtures purchased from Pharmingen (San Diego, CA, USA). A direct staining method was used to evaluate a minimum of 5,000 gated events for each sample with a FACSCalibur™ flow cytometer and the CellQuest™ software version 3.1 (Becton Dickinson, Inc., San Jose, CA, USA). The mAbs were conjugated with fluorescein isothiocyanate (FITC), R-phycoerythrin (PE), allophycocyanin (APC), or peridinin chlorophyll protein (PerCP). Firstly, the CD45 marker was used to identify the leukocytes; mononuclear cells (MNC) were then gated on CD45 and side scatter. mAbs specific for CD3 (T-cells), CD4 (T-helper or Th subset), CD8 (T-cytotoxic or Tc subset), CD19 (B-cells), and NK1.1 (NK cells) were used for further analysis. T-Regulatory (T-reg) cell analysis was carried out with a staining kit which included FJK-16s*PE [anti-forkhead box P3 (FoxP3)], CD4*FITC, and CD25*APC (eBioscience Inc., San Diego, CA, USA). The CD4+CD25+ T-cells, both with and without FoxP3, were quantified by gating on side scatter and the CD4+ cells, followed by analysis of the CD25+ versus FJK-16s+ (FoxP3+) subset.
Activation of splenocytes using anti-CD3-coated plates. The splenocytes were quantified after lysis of RBC using an automated hematology analyzer (HESKA Corp.). The density of splenocytes was adjusted to 2×106 cells/ml with complete RPMI-1640 medium and 100 μl/well were dispensed into 96-well plates coated with immobilized anti-CD3 antibody (anti-Mouse CD3 BioCoat™ T-Cell Activation, 96-well Assay Plates; BD Pharmingen, San Diego, CA, USA). One-hundred microliters of RPMI-1640 was added to each well to make the total volume equal to 200 μl/well. This procedure results in the activation of the T-cell receptor (TCR)/CD3 signaling machinery. The plates were then incubated at 37°C for 48 h and supernatants were harvested after 48 h and stored at −80°C until further analysis.
Cytokine analysis. Spleen supernatants at −80°C were thawed and analyzed for 22 different cytokines and chemokines using the Mouse Cytokine/Chemokine Milliplex MAP Kit (Millipore, Billerica, MA, USA). The cytokines/chemokines evaluated were as follows: interleukin-1α (IL-1α), IL-1β, IL-4, IL-5, IL-6, IL-7, IL-9, IL-10, IL-12(p70), IL-13, IL-15, IL-17, granulocyte colony-stimulating factor (G-CSF), granulocyte-macrophage (GM)-CSF, interferon-γ (IFN-γ), IFN-γ-induced protein 10 (IP-10), keratinocyte chemoattractant (KC, also known as CXCL1), monocyte chemotactic protein-1 (MCP-1), macrophage inflammatory protein-1α (MIP-1α), regulated upon activation, normal T-cell expressed and secreted (RANTES) and tumor necrosis factor-α (TNF-α). Vascular endothelial growth factor (VEGF) was analyzed in the anti-CD3-activated supernatants using a VEGF enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis, MN, USA).
Statistical analysis. The data were analyzed using one-way or two-way analysis of variance (ANOVA). Tukey's test was performed for pair-wise multiple comparisons, when indicated. Means and standard errors of means (SEM) are presented here; results from each mouse were included in the analyses. A p-value of <0.05 indicated significance. SigmaStat™ software, version 2.03 (SPSS Inc., Chicago, IL, USA) was used to analyze the data.
Results
Relative spleen mass. As indicated in Figure 1, radiation generally reduced the RSM (p<0.05). Although there was no minocycline by radiation interaction, treatment with the drug increased RSM regardless of radiation. In post-hoc analysis, minocycline significantly (p<0.05) increased RSM in all radiation groups except for the one treated with 3 Gy, where only a trend for higher RSM was present (p<0.1).
Erythrocyte and platelet parameters in blood. There were no consistent main effects or interactions between radiation and minocycline treatment on any RBC or platelet parameter (Table I). In post-hoc analysis of RBC counts, there were radiation-induced reductions in the dH2O-treated groups that were irradiated with 1 and 3 Gy compared to their 0 Gy counterpart (p<0.05). This was also reflected in radiation-induced increases in RDW in the same groups (p<0.05). There were minocycline-induced reductions in MCV and RDW in the 3 Gy groups (p<0.05), as well as an increase in MPV in the 2 Gy group.
WBC and leukocyte populations in blood. Figure 2 shows the counts and percentages of lymphocytes, monocytes and granulocytes in the blood. Radiation generally reduced the counts of all three of these cell types (p<0.05), resulting in a significant radiation dose-dependent decrease in total WBC number. Addition of the drug did not significantly enhance any of the cell counts. The drug-induced decrease in lymphocytes was also apparent in the total WBC counts at 0 Gy (p<0.05 versus dH2O) (Figure 2). In terms of percentages, the granulocytes increased in the presence of the drug at all radiation doses, while the lymphocyte percentages decreased at 2 and 3 Gy (p<0.05). The changes in eosinophil counts and percentages were similar to the overall pattern of changes noted for granulocytes (Table II).

Relative spleen mass. Each bar represents the mean±SEM for n=10 mice/group. Two-way ANOVA: §p<0.05 for main effect of radiation; †p<0.05 for main effect of drug. Tukey test: *p<0.05 dH2O vs. minocycline at each radiation dose; ap<0.05 vs. 0 Gy within dH2O-treated groups; bp<0.05 vs. 0 Gy within minocycline-treated groups;Tp<0.1 vs. dH20 plus 3 Gy.
WBC and leukocyte populations in spleen. The distribution of lymphocytes, macrophages and granulocytes in the spleen is shown in Figure 3. Radiation generally caused a decline in the counts of these three leukocyte types (p<0.05). Treatment with minocycline caused a significant increase in granulocyte and macrophage counts in both the 0 Gy- and 2 Gy-irradiated groups (p<0.05), resulting in a significant main effect of this drug. The pattern of changes in these leukocyte types was reflected in the total WBC counts (Figure 3). A significant increase in granulocyte percentages was observed in the presence of minocycline in the un-irradiated group (p<0.05). The lymphocyte percentages decreased significantly in all groups on treatment with minocycline (p<0.05). Splenic eosinophil counts and percentages had similar patterns to those of granulocytes (Table II).
Flow cytometric analysis of lymphocyte populations in spleen. Radiation caused a decrease in the T-, B- and NK cell counts (p<0.05, Figure 4). There were main effects of minocycline on the T-cell and NK cell populations (p<0.05). In the post-hoc analysis, addition of the drug significantly increased the T-cell counts at 0 Gy, 2 Gy and 3 Gy (p<0.05 versus the corresponding groups without drug). Minocycline alone increased the B-cell counts in the 0 Gy group (p<0.05). Although drug-treated groups had consistently higher NK cell counts, significance was found only at 2 Gy (p<0.05).
Summary of erythrocyte and platelet characteristics in blood of mice treated or not with radiation and/or minocycline.
Counts and percentages of eosinophils in blood and spleen from mice treated or not with radiation and/or minocycline.
Consistent with the total T-cell count, radiation caused a decline in the CD4+ Th and CD8+ Tc cells (p<0.05, Figure 4). While there was no significant impact of minocycline on these two subsets, the high counts in the 0 Gy and 2 Gy groups (p<0.05) resulted in there being a main effect of the drug (p<0.05). Both radiation and minocycline generally increased CD4:CD8 cell ratios (p<0.05). However, for radiation, this was primarily due to changes in the 2 and 3 Gy-treated groups. Treatment with the drug generally mitigated or abolished the radiation-induced increases at the higher doses (p<0.05), resulting in a significant drug-by-radiation interaction (p<0.05).
When looking at the percentages (Figure 5), radiation caused a decrease in B-cells, with corresponding increases in the T- and NK cell populations (p<0.05). Since minocycline caused a significant proportional increase in B-cells at 0 Gy (p<0.05), there was a significant main effect of the drug (p<0.05) and a drug-by-radiation interaction (p<0.05). Radiation-induced increases in NK cell percentages were generally augmented when mice were treated with minocycline. However, in post-hoc analysis, the drug-induced increase reached significance only in the 2 Gy-treated group (p<0.05), resulting in a significant drug-by-radiation interaction (p<0.05).
Overall, radiation increased Th cell percentages, while reducing that of Tc cells (p<0.05, Figure 5). The drug did not significantly impact the proportion of Th cells. Although the drug caused an increase in the Tc cell percentages (p<0.05), significance was found only at 2 Gy in post-hoc analysis (p<0.05); this led to a significant drug-by-radiation interaction (p<0.05).

Leukocyte populations in blood. The data were obtained using an automated hematology analyzer. Each bar represents the mean±SEM for n=9-10 mice/group. Two-way ANOVA: §p<0.05 for the main effect of radiation; †p<0.05 for the main effect of drug; ‡p<0.05 for interaction between radiation and minocycline, Tukey test: *p<0.05 dH2O vs. minocycline at each radiation dose; ap<0.05 vs. 0 Gy within dH2O-treated groups; bp<0.05 vs. 0 Gy within minocycline-treated groups.
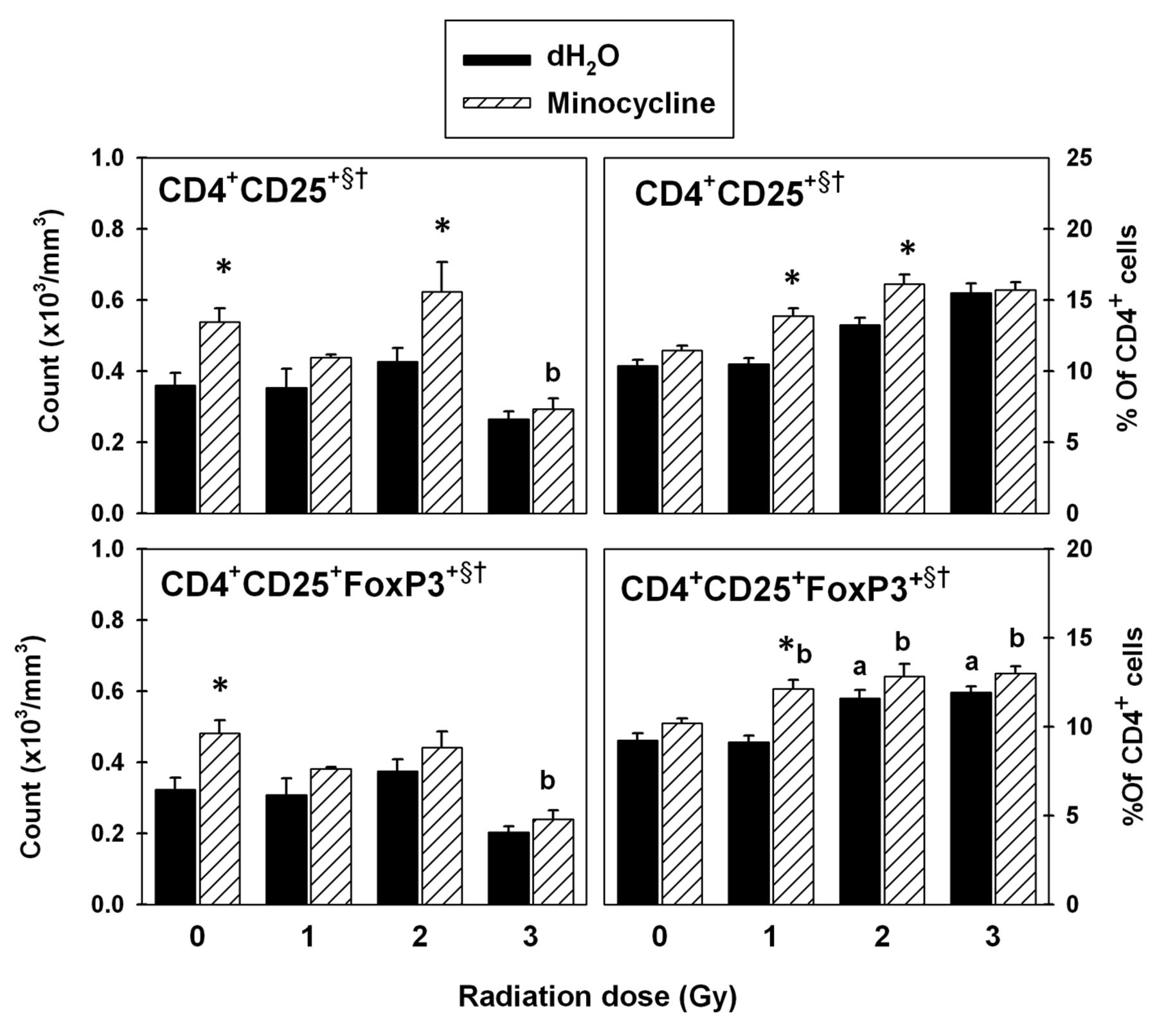
Flow cytometric analysis of CD4+CD25+FoxP3+ T-cells in spleen. Radiation generally increased the CD4+CD25+ T-cell counts (Figure 6). Addition of minocycline caused a significant increase at 0 Gy and 2 Gy (p<0.05). This resulted in significant main effects of radiation and drug on these cells (p<0.05). Radiation also caused a significant decrease in the CD4+CD25+FoxP3+ T-reg cell counts (p<0.05). Although minocycline-treated groups had consistently higher counts, significance was found only at 0 Gy (p<0.05). This resulted in significant main effects of radiation and drug on the CD4+CD25+FoxP3+T-reg population (p<0.05).

Leukocyte populations in spleen. The data were obtained using an automated hematology analyzer. Each bar represents the mean±SEM for n=9-10 mice/group. Two-way ANOVA: §p<0.05 for main the effect of radiation; †p<0.05 for the main effect of drug; ‡p<0.05 for interaction between radiation and minocycline. Tukey test: *p<0.05 dH2O vs. minocycline at each radiation dose; ap<0.05 vs. 0 Gy within dH2O-treated groups; bp<0.05 vs. 0 Gy within minocycline-treated groups.
While radiation generally increased the percentages of CD4+CD25+ T-cells with increasing dose, addition of minocycline significantly enhanced the effect of radiation at 1 Gy and 2 Gy (p<0.05; Figure 6). This resulted in main effects of radiation and drug on CD4+CD25+ T-cell percentages (p<0.05). Radiation at 2 Gy and 3 Gy increased the CD4+CD25+FoxP3+ T-reg cell percentages significantly (p<0.05). At 1 Gy, the percentage of these cells was higher compared to the group without drug (p<0.05).
Cytokines in spleen supernatants. Numerous changes occurred in cytokine levels that were dependent on minocycline, radiation, or both. Hence, for the sake of clarity in the discussion, as well as simplicity, each cytokine was assigned to one of two groups. Group I included cytokines that increased in the presence of the drug, while group II included cytokines that were reduced in the presence of the drug.
Group I cytokines. This group included G-CSF, GM-CSF, IL-1α, IL-1β, IL-4, IL-5, IL-6, IL-10, IL-13, IL-17 and RANTES (Figure 7). Two-way ANOVA showed that radiation had a significant main effect (p<0.05) on the levels of all of these cytokines except IL-10, IL-17 and RANTES, while a main drug effect was noted for all except IL-5. A drug-by-radiation interaction (p<0.05) was noted for all of the cytokines in this group except for IL-10 and RANTES.

Lymphocyte population counts in spleen. The data were obtained using fluorescence-labeled monoclonal antibodies and flow cytometry. Each bar represents the mean±SEM for n=5-10 mice/group. Two-way ANOVA: §p<0.05 for the main effect of radiation; †p<0.05 for the main effect of drug; ‡p<0.05 for interaction between radiation and minocycline. Tukey test: *p<0.05 dH2O vs. minocycline at each radiation dose; ap<0.05 vs. 0 Gy within dH2O-treated groups; bp<0.05 vs. 0 Gy within minocycline-treated groups.
Post-hoc Tukey analysis revealed that minocycline significantly enhanced the concentrations of IL-13 and RANTES in non-irradiated and in all irradiated groups compared to the corresponding groups that did not receive the drug (p<0.05). Significant drug-induced increases in G-CSF, GM-CSF and IL-1β occurred in all irradiated groups. With respect to the remaining cytokines, spleen supernatants from irradiated mice treated with minocycline had significantly higher levels of IL-1α, IL-4, IL-5, IL-6, IL-10 and IL-17 compared to their counterparts exposed to 1, 2 or 3 Gy (p<0.05).
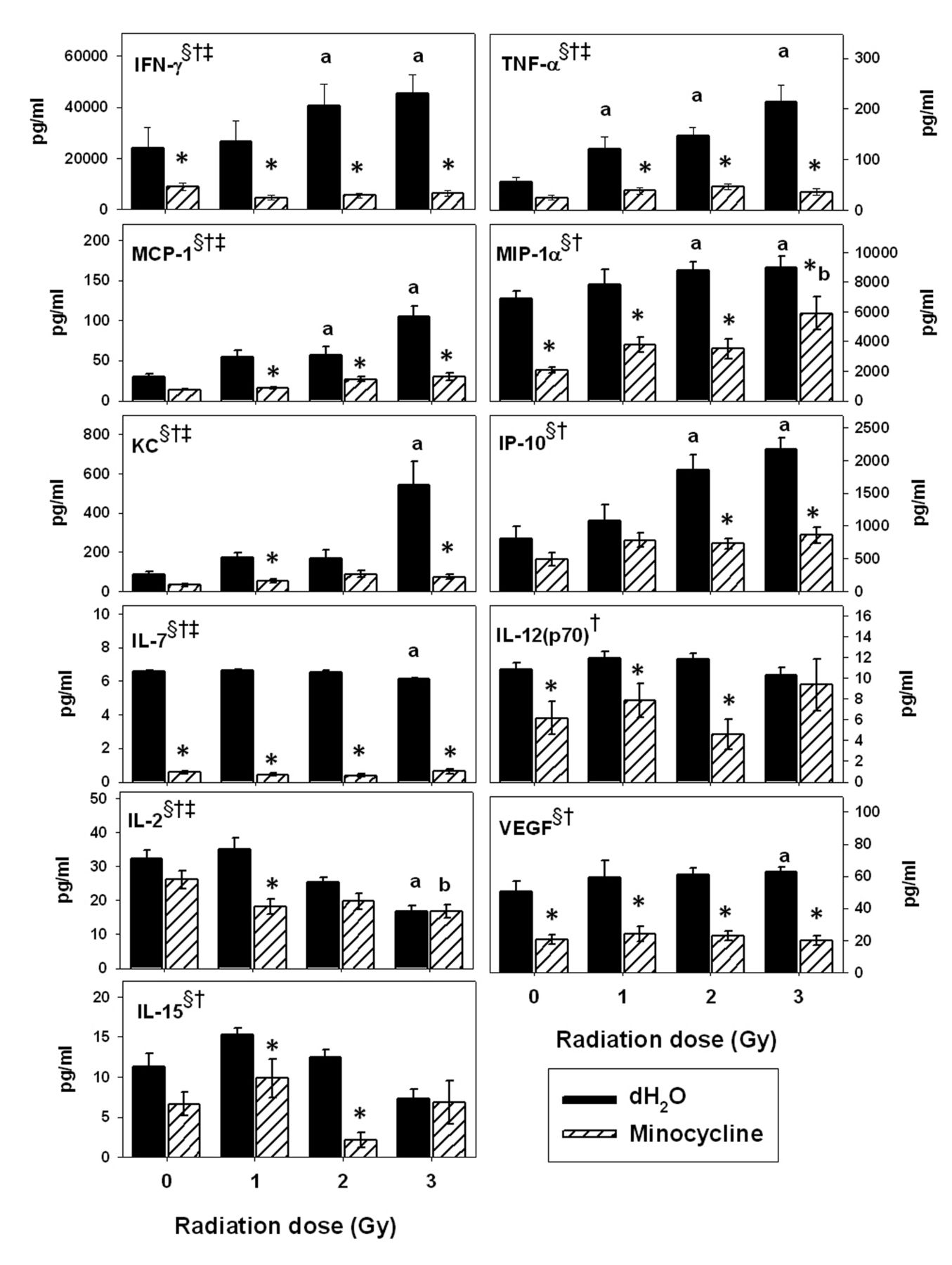
Group II cytokines. This group consisted of IFN-γ, TNF-α, MCP-1, MIP-1α, KC, IP-10, IL-7, IL-12(p70), IL-2, VEGF and IL-15 (Figure 8). Two-way ANOVA showed that radiation had a significant main effect on the levels of all of these cytokines except IL-12(p70) and that minocycline had a main effect on all of them (p<0.05). A significant drug-by-radiation interaction was noted for IFN-γ, TNF-α, MCP-1, KC, IL-7, IL-12(p70) and IL-2 (p<0.05).

Lymphocyte population percentages in spleen. The data were obtained using fluorescence-labeled monoclonal antibodies and flow cytometry. Each bar represents the mean±SEM for n=5-10 mice/group. Two-way ANOVA: §p<0.05 for the main effect of radiation; †p<0.05 for the main effect of drug; ‡p<0.05 for interaction between radiation and minocycline. Tukey test: *p<0.05 dH2O vs. minocycline at each radiation dose; ap<0.05 vs. 0 Gy within dH2O-treated groups; bp<0.05 vs. 0 Gy within minocycline-treated groups.
The post-hoc Tukey test revealed that radiation, especially at the higher doses, significantly enhanced the levels of IFN-γ, TNF-α, MCP-1, MIP-1α, KC, IP-10 and VEGF and reduced the levels of IL-2 and IL-7 compared to 0 Gy (p<0.05). However, addition of minocycline frequently minimized these radiation-induced changes. Mice treated with the drug had significantly lower IFN-γ, MIP-1α, IL-7 and VEGF concentrations in spleen supernatants (p<0.05) compared to their irradiated counterparts, regardless of dose; low levels were also noted compared to 0 Gy (p<0.05). In addition, the drug reduced the enhancing effects of radiation on MCP-1, KC, IP-10 and IL-15, although statistical significance was not obtained for all radiation doses.
Discussion
Our study is unique in that it characterized the impact of minocycline on important immune parameters relevant to ARS in an irradiated mammalian model. In essence, the results indicate that the drug has the potential to promote hematopoietic recovery after radiation exposure. The significantly increased RSM is a broad indicator that supports minocycline as an enhancer of hematopoiesis. Indeed, spleen weight has long been used as an index for determining hematopoietic recovery after irradiation (14).
A serious consequence of radiation injury is bone marrow failure, resulting in leukopenia that can influence morbidity and mortality (15). Agents that act as radiation protectants/mitigators when used near the time of radiation exposure are urgently needed. Many of the compounds under investigation stimulate the differentiation of stem cells in bone marrow (16). In the present study, there were very few and relatively minor differences among groups in erythrocyte and platelet parameters. However, there were numerous differences in cytokine concentrations and distribution patterns of specific leukocyte populations that are well-known to secrete important immunomodulatory cytokines. Since the major goal was to quantify the efficacy of minocycline as a radioprotectant against ARS, for the sake of clarity, most of the remainder of the discussion is divided into two sections based on the immunomodulatory effects of the drug. Group I cytokines were generally increased by minocycline, whereas the group II cytokines were reduced.
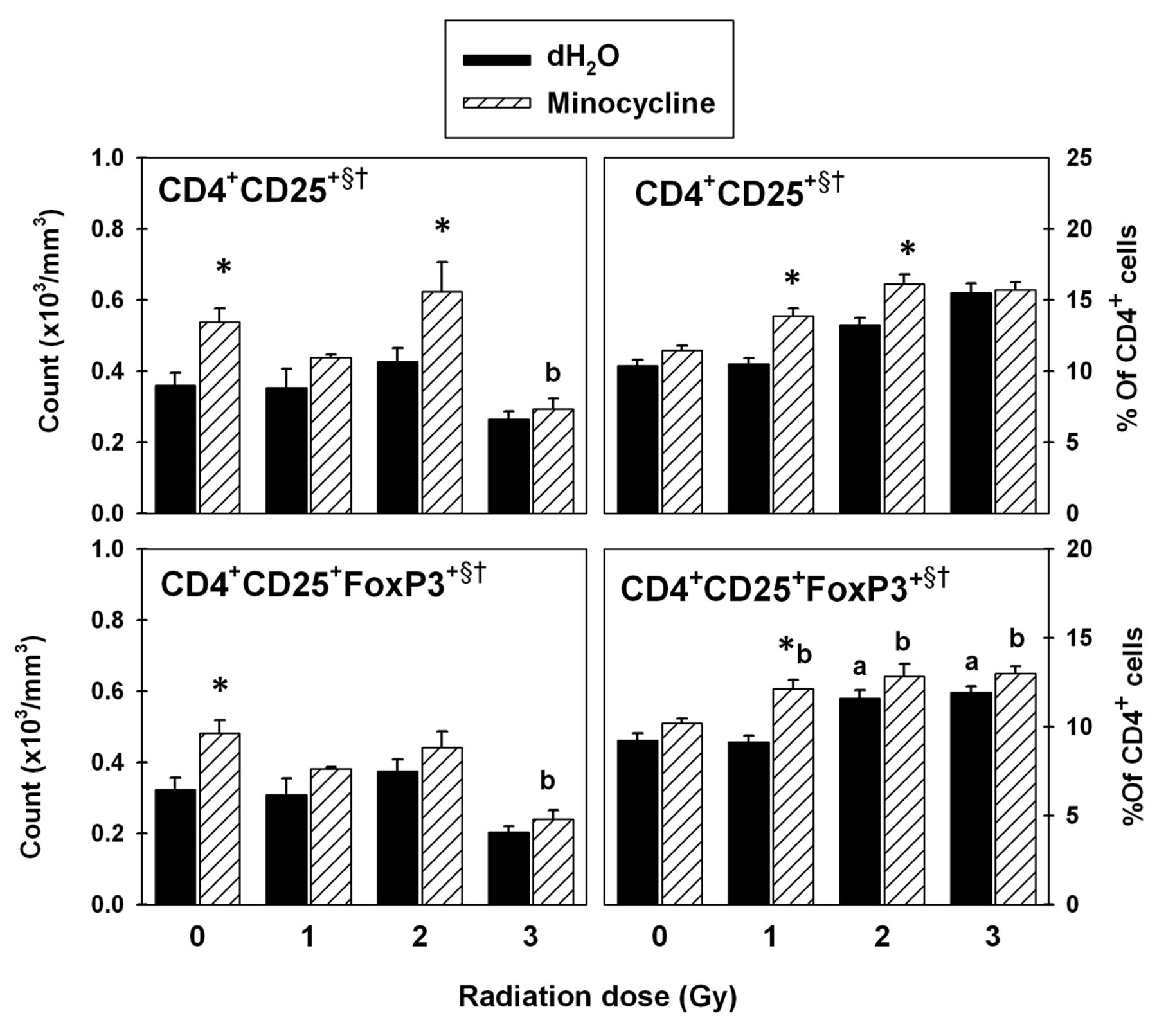
T-regulatory cells in spleen. The data were obtained using fluorescence-labeled monoclonal antibodies and flow cytometry. Each bar represents the mean±SEM for n=5-10 mice/group. Two-way ANOVA: §p<0.05 for main effect of radiation; †p<0.05 for main effect of drug. Tukey test: *p<0.05 dH2O vs. minocycline at each radiation dose; ap<0.05 vs. 0 Gy within dH2O-treated groups; bp<0.05 vs. 0 Gy within minocycline-treated groups.
Group I: Cytokines up-regulated by minocycline and related cell populations. Treatment with minocycline increased G-CSF, GM-CSF and IL-1β in spleen supernatants from irradiated mice. Furthermore, these increases were incrementally enhanced with increasing radiation dose. Similar, although less pronounced, radiation-dependent drug effects were noted in IL-1α, IL-4, IL-5 and IL-6.
Although T-cells in the spleen samples were activated with anti-CD3 mAbs, other cell types (e.g. fibroblasts), may have contributed to the levels of G-CSF, GM-CSF, IL-5 and IL-6, particularly in the groups treated with minocycline (17). However, the pattern for all of these cytokines is remarkably similar to that of the granulocyte percentages in the spleen (as well as in blood). This is important because the assay involved a constant number of leukocytes per well and proportional differences likely played a dominant role in the cytokine response. Neutrophils tend to persist when microenvironmental signals that involve infection are present (18) and a proportional increase in this population may be due to similar signals generated by radiation-induced cell debris. This increased survival could lead to an increase in the presence of cytokines produced by this population.
Indeed, neutrophils, the major type of granulocyte, and monocytes can secrete GM-CSF, IL-1α and IL-1β (19). In addition, GM-CSF induces production of IL-1, IL-6 and G-CSF by neutrophils (20). Although IL-1α and IL-1β are two distinct cytokines, they bind to the same receptors (21). Regardless of the source, a minocycline-induced increase in these cytokines is important because they can be radioprotective or aid in recovery post-exposure (22). For instance, lethality due to total-body irradiation is reduced substantially by G-CSF treatment (23). G-CSF and GM-CSF hasten myeloid progenitor cell proliferation (24); GM-CSF also inhibits radiation-induced apoptosis (25) and promotes eosinophil growth and function (26). Administration of IL-1 improves survival in animals (27). Radioprotective mechanisms of IL-1 include: a) induction of bone marrow cells into the relatively radioresistant S phase of the cell cycle (28); b) induction of manganese superoxide dismutase (MnSOD) (29) which protects against oxidative stress (30); and c) facilitation of granulopoiesis (31). Interestingly, some researchers also mention that IL-1 can be cytotoxic to tumors (32).

Quantitative analysis of granulocyte colony-stimulating factor (G-CSF), granulocyte-monocyte CSF (GM-CSF), interleukin-1α (IL-1α), IL-1β, IL-4, IL-5, IL-6, IL-10, IL-13 and IL-17 and regulated upon activation, normal T-cell expressed and secreted (RANTES). Each bar represents the mean±SEM for n=8-10 mice/group. Two-way ANOVA: §p<0.05 for main the effect of radiation; †p<0.05 for the main effect of drug; ‡p<0.05 for interaction between radiation and minocycline. Tukey test: *p<0.05 dH2O vs. minocycline at each radiation dose; bp<0.05 vs. 0 Gy within minocycline-treated groups.
Although minocycline generally increased IL-4, IL-5 and IL-6 expression, its impact was most prominent in the mice that also underwent 3 Gy irradiation. While a common factor among these cytokines is activation of T- or B-cells (33), they also have other activities. IL-4 and IL-5 are Th2 cell-derived cytokines (34), IL-4 stimulates hematopoietic progenitor cell proliferation (35), and IL-5 promotes proliferation and differentiation of eosinophils (36). The up-regulation of IL-5 in the presence of minocycline may explain the correlative increase in eosinophils. Similar patterns were observed for IL-6, a cytokine produced by dendritic cells, macrophages and B-cells (37) that promotes differentiation of the Th2 lineage while inhibiting Th1 cell differentiation (37). IL-6 enhances hematopoietic recovery by accelerating mutilineage hematopoiesis and increasing myeloid progenitors (22, 24); its administration after lethal radiation promotes survival in animals (27).

Quantitative analysis of interferon-γ (IFN-γ), tumor necrosis-α (TNF-α), monocyte chemotactic protein-1 (MCP-1), macrophage inflammatory protein-1α (MIP-1α), keratinocyte chemoattractant (KC), IFN-γ-induced protein 10 (IP-10), interleukin-7 (IL-7), IL-12(p70), IL-2, vascular endothelial growth factor (VEGF) and IL-15. Each bar represents the mean±SEM for n=8-10 mice/group. Two-way ANOVA: §p<0.05 for main effect of radiation; †p<0.05 for the main effect of drug; ‡p<0.05 for interaction between radiation and minocycline. Tukey test: *p<0.05 dH2O vs. minocycline at each radiation dose; ap<0.05 vs. 0 Gy within dH2O-treated groups; bp<0.05 vs. 0 Gy within minocycline-treated groups.
For IL-10 and IL-17, the drug response was biphasic, where significant drug-induced increases occurred only in the 1 Gy- and 3 Gy-treated mice. Values for the 0 Gy and 2 Gy groups, however, were consistently higher when minocycline was included. IL-10 is produced by macrophages, Th2 cells and especially T-reg cells (38), while IL-17 is produced by the CD4+ (39) and CD8+ T-cell subsets (40). However, the pattern of secretion was not reflected in the proportions of the mentioned cells. IL-10 inhibits the suppressive activity of IFN on hematopoiesis (41). The minocycline-induced increase in IL-10 is in accordance with previous observations (42). An interesting additional point is that IL-10 may reduce risk for fibrosis (43, 44), a serious late consequence of radiation treatment. IL-17, similarly to IL-10, induces the production of hematopoietic cytokines, e.g. IL-6 and G-CSF (39).
IL-13 and RANTES were increased by minocycline, but radiation had no detectable influence on their levels. In contrast to other cytokines in group I, IL-13 is produced by Th2 (45), NK (46) and dendritic cells (47), as well as eosinophils (48, 49). The minocycline-induced increases found here are consistent with reports that the drug preferentially stimulates Th2 cells over Th1 (50), thus enhancing IL-13 production. Furthermore, the high eosinophil percentages observed in the presence of the drug could also contribute to this response. Since IL-13 stimulates the growth of hematopoietic progenitor cells (51), minocycline could help speed-up recovery. RANTES is secreted by CD8+ T-cells (52, 53), including memory T-cells that tend to be relatively radioresistant (54). This could explain the lack of a radiation response in the level of this chemokine despite an overall decrease in total lymphocyte proportions. NK cells also produce RANTES (55) and this correlated with increased NK cell percentages. RANTES attracts monocytes, eosinophils and T-cells to sites of tissue damage (56, 57), is required for normal T-cell function (58), and increases CD8+ T-cell counts (59).
Group II: Cytokines/chemokines down-regulated by the drug and related cell populations. While radiation increased production capacity for IFN-γ, TNF-α, MCP-1, MIP-1α, KC and IP-10, minocycline dramatically reduced their expression. Since many of these cytokines inhibit hematopoietic activity, the minocycline-induced decreases further support a role for this drug in hematopoietic recovery. The low levels of IFN-γ and TNF-α noted here are consistent with the literature (60). Both of these cytokines can inhibit hematopoiesis, but also have potent pro-inflammatory effects (61-63). Activation of the p38 mitogen-activated protein kinase (p38 MAPK) pathway can lead to suppression of hematopoiesis. This pathway also activates TNF-α and IFN-γ which are myelosuppressive cytokines (64). Minocycline has been shown to inhibit the p38 MAPK pathway (65). The drug may be helpful in restoration of hematopoiesis because it inhibits this pathway.
MCP-1 is produced by keratinocytes and attracts monocytes, basophils and T-cells (66). Like IFN-γ and TNF-α, MIP-1α down-regulates the proliferation of hematopoietic progenitor cells (67). Karpus and Kennedy found that anti-MIP-1α treatment prevented the occurrence of acute experimental autoimmune encephalitis (68). Since minocycline reduced both MIP-1α and MCP-1 production, it may aid in the repression of inflammation that occurs after radiation insult.
KC, a chemokine that attracts neutrophils and monocytes, has been reported to increase in response to radiation (69). Recent studies have shown that it can be produced by pulmonary fibroblasts in response to radiation and thus may be utilized as a marker of radiation-induced lung injury (69). Monocytes, lymphocytes, keratinocytes and endothelial cells produce IP-10 in response to IFN-γ (70). As with many of the other cytokines in this group, IP-10 inhibits early hematopoietic progenitors (71). It can also inhibit the actions of GM-CSF (72). Finally, IP-10 has been implicated in radiation-induced lung fibrosis (73).
In contrast to the first six cytokines in group II, the impact of radiation on IL-7, IL-12(p70) and VEGF was minimal (albeit significant for both IL-7 and VEGF), while minocycline caused significant decreases in most groups of mice. IL-7 is produced by stromal cells in the bone marrow, spleen, thymus and gut (74) and is important for the development of B- and T-cells (75). Administration of IL-7 to patients with metastatic melanoma or sarcoma reduced the number of CD4+CD25+FoxP3+ T-reg cells (76). The T-reg cells restrict inflammation and prevent carcinogenesis by reducing cellular damage and cell proliferation (77). It has also been reported that naturally-occurring T-reg cells secrete high amounts of thioredoxin, which confers increased tolerance to oxidative stress (78). However, IL-7 has also been reported to promote development of T-cell lymphoma and acute lymphoblastic leukemia (79). Thus, the very low level of IL-7 in the minocycline-treated groups suggests that the risk for these types of malignancies may be reduced by the drug.
IL-12 is produced by dendritic cells, macrophages and B-cells (80). Golab et al. found that this cytokine has myelosuppressive properties and slows down hematopoiesis in the bone marrow (81). Although IL-12 promotes hematopoiesis in the spleen, this does not fully compensate for its depressive effect on cells in the bone marrow (71). Moreover, since IL-12 induces the production of IFN-γ by Th1 cells (82), the low IL-12 levels correlate well with the reduced IFN-γ levels seen in our study.
VEGF is typically produced by endothelial cells, platelets and macrophages (83, 84). Previous researchers have shown that levels of this cytokine tend to increase after radiation exposure (85). VEGF is a potent promoter of angiogenesis and helps to facilitate tumor growth. Thus, the minocycline-induced reduction in the high VEGF levels seen in the irradiated groups adds to the potential of this drug for utilization during cancer radiotherapy.
Like other cytokines in group II, levels of IL-2 and IL-15 induced by the anti-CD3 mAb were generally reduced by minocycline treatment. These findings are consistent with reports that this drug inhibits T-cell activation (86). IL-2 is produced by T-lymphocytes, while activated splenic dendritic cells can produce IL-15 in response to IFN-γ (87), the latter being a product of activated Th1 cells. Venkataraman and Westerman have reported that radiation causes increased production and decreased utilization of IL-2 by mononuclear cells (88). Thus, suppression of T-cell activation by minocycline is a likely explanation for reduced IL-2 and IL-15 levels. However, unlike all of the other cytokines in group II, IL-2 and IL-15 levels generally decreased with increasing radiation dose. This is consistent with the low T-cell proportions found in the mice of irradiated groups. These two cytokines share many biological properties, such as promotion of T-cell proliferation and NK cell development. However, while IL-15 is primarily implicated in inflammatory diseases, IL-2 can both promote and control inflammation (89).
Conclusion
In summary, our data demonstrate that treatment with minocycline modulates the capacity to produce cytokines, related to hematopoiesis, in irradiated and non-irradiated mice. The drug also counteracted radiation-induced declines in certain cell populations, especially of monocytes/macrophages, granulocytes, NK cells, T-cells and CD8+ T-cells in the spleen. Protection of these cell populations can be attributed to the cytokine pattern obtained, with special emphasis on increased G-CSF, GM-CSF, IL-1α, IL-1β levels and reduced IFN-γ, TNF-α, MIP-1α, IL-12(p70) and IL-15 levels. Minocycline up-regulated both pro- and anti-inflammatory cytokines/chemokines and cell populations that could promote, as well as restrict, inflammation. This implies that the drug may balance the radiation-induced inflammatory response, preventing it from getting out of control, while simultaneously enhancing the body's capacity to maintain innate and adaptive immune cell types. Overall, the results do indicate that minocycline up-regulates production capacity for several cytokines that have been strongly implicated in myelorestoration. The findings also suggest that the drug may be utilized to speed up hematopoietic recovery post-irradiation. Further testing of minocycline should be carried out to confirm its potential as a countermeasure against ARS and as an adjuvant during cancer radiotherapy.
Acknowledgements
The assistance of Drs. Xiao Wen Mao, Xian Luo-Owen and Jian Tian, as well as Steven Rightnar, Celso Perez, Gordon Harding, Leticia Ortloff, Linda Ritter and Brandon Bianski, in various aspects of this study is greatly appreciated. The study was supported by National Aeronautics and Space Administration grant NNX10AJ31G and the LLUMC Department of Radiation Medicine.
- Received March 29, 2012.
- Revision received May 26, 2012.
- Accepted May 28, 2012.
- Copyright © 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}