


Abstract
Background: Aberrant activation of Wnt/β-catenin signaling promotes the development of several types of cancer. Recently, it has been demonstrated that Wnt pathway is also activated in myeloma. Therefore, the Wnt/β-catenin signaling molecules are attractive candidates for development of targeted therapies for this disease. Materials and Methods: This study investigated the antitumor effect of ciclopirox olamine (CIC) in vitro and in vivo in a murine myeloma model. Results: CIC demonstrated major apoptotic activity in different human and murine myeloma and lymphoma cell lines, as well as in human primary cells. In addition, β-catenin expression was down-regulated when CIC was added to lymphoma cells. Interestingly, in vitro, a significant additive effect was seen with the combination of lenalidomide plus CIC as compared to single applications. In vivo, tumor growth, was significantly reduced and overall survival increased in mice treated with CIC as compared to untreated mice. Conclusion: These results reveal a significant selective induction of apoptosis by CIC and suggest a significant in vivo effect against myeloma.
Myeloma remains an incurable disease, although major progress has been achieved in its treatment by the introduction of novel agents such as thalidomide, lenalidomide and bortezomib. In addition, in patients with refractory diseases further novel agents are urgently needed. Recently, several groups have shown that the Wnt/β-catenin pathway plays an important role in the regulation of cell proliferation, differentiation and apoptosis (1-3). Aberrant activation of the Wnt signaling pathway has major oncogenic effects (4-7). In the canonical Wnt pathway, secreted Wnt proteins bind to a receptor complex, consisting of a member of the Frizzled (Fzd) family, and the low-density lipoproteinreceptor-related proteins (LRP) 5 or LRP6. Subsequently, the cytoplasmic adaptor protein disheveled (Dvl) is phosphorylated and inhibits glycogen synthase kinase (GSK)-3β activity through its association with axin. Unphosphorylated β-catenin accumulates in the cytoplasm and translocates into the nucleus, where it interacts with T-cell (TCF) and lymphoid-enhancing (LEF) factors to activate transcription of Wnt target genes (4, 5, 8). In addition, it has been demonstrated that the Wnt pathway is activated in lymphoma and myeloma. Therefore, the Wnt/β-catenin signaling molecules are attractive candidates for development of targeted approaches in lymphoma and multiple myeloma treatment.
In addition, we recently confirmed that the diuretic agent ethacryanic acid (EA) and the antifungal agent ciclopirox olamine (CIC) inhibit Wnt/β-catenin signaling (9). CIC is a synthetic antifungal agent used topically for the treatment of yeast infections in humans and is degraded by glucoronidation (10) acting by inhibiting the membrane transfer system by interrupting Na+ K+ ATPase (11).
For all of these reasons, we tested the effect of CIC on myeloma cells as a single agent, as well as in addition to the commonly used therapeutic drugs.
Materials and Methods
Cell lines and culture conditions. The lymphoma cell lines LAM-53, SU-DHL-4, Daudi and Raji, as well as the myeloma cell lines OPM-2, RPMI-8226 and U-266 (all obtained from DSMZ, Collection of Microorganisms and Cell Culture, Braunschweig, Germany) were cultured in RPMI-1640 medium consisting of 10% heat-inactivated fetal calf serum (FCS; Invitrogen, Karlsruhe, Germany), 2.5% 1 M HEPES, and 1× penicillin/streptomycin (all from PAA Laboratories GmbH, Cölbe, Germany). Cells were cultured at a density of 3.3×105 cells ml−1 and incubated at 37°C with 5% CO2 and 95% humidity.
MPC11 (DMSZ) is a murine plasmocytoma cell line derived from the Balb/c strain expressing the immune globuline G2b. Cells were cultured in RPMI-1640 medium (PAA Laboratories GmbH, Austria) supplemented with 5% FCS, 2 mM glutamine (both from PAA, Cölbe, Germany) and 100 U/ml penicillin/100 U/ml streptomycin (both from Seromed, Jülich, Germany) at 37°C in a humidified 5% CO2 atmosphere.
Human samples. Peripheral blood mononuclear cells (PBMCs) and peripheral blood lymphocytes (PBLs) were isolated from blood samples of healthy volunteers by Ficoll density gradient centrifugation (Lymphoprep; Nycomed, Oslo, Norway). In addition, bone marrow samples from patients with myeloma were obtained. Ethics approval had been obtained according to the guidelines of the participating institutions.
Drugs and chemical reagents. In our experiments the following drugs were used in vitro: CIC (Sigma-Aldrich GmbH, Seelze, Germany), doxorubicin (Cellpharm GmbH, Hanover, Germany), rituximab (Roche Pharma AG, Grenzach Wyhlen, Germany), bortezomib (Janssen Cilag, Neuss, Germany), thalidomide (Grünenthal Pharma GmbH, Aachen, Germany) and lenalidomide (Celgene; Munich, Germany). All drugs were tested at different concentrations for 24-72 hours.
DiOC6 and propidium iodide (PI) staining. Experiments were performed as reported recently (12). A total of 1×105 Cells were cultured in 3 ml medium in 6-well plates. CIC was dissolved in dimethyl sulfoxide (DMSO), and added at an optimized concentration of 10 μM alone, or in combination with the therapeutic agents at different concentrations for three days. The apoptosis assay was performed with 3’3-dihexyloxacarbocyanine iodide (DiOC6) staining for mitochondrial membrane potential in viable cells, and propidium iodide (PI) which binds to DNA in necrotic cells, measured by a fluorescence-activated cell sorter (FACS).
For FACS analysis, 500 μl staining solution containing 80 nM DiOC6 in FACS buffer, consisting of deficient RPMI medium with 0.5% bovine serum albumin (BSA), was mixed with equal volumes of the cell sample in a glass tube and incubated at 37°C for 15 min. After a washing step with 1% PBS/BSA the cells were re-suspended in 500 μl of 1% PBS/BSA. After addition of 5 μl PI solution (100 μg/ml), the cells were analyzed by FACS. Using this assay, viable cells reveal high fluorescence intensity for DiOC6 and a low intensity for PI. On the contrary, apoptotic cells have a low intensity for DiOC6 and also a low intensity for PI. Finally, necrotic cells have a low intensity for DiOC6 and a high intensity for PI.
Isolation of PBMCs. PBMCs were isolated from blood of healthy donors by Ficoll-Hypaque density gradient centrifugation. Blood from buffy coats was mixed 1:2 with PBS/1% BSA (both PAA) and used for a ficoll gradient (LymphoPrep; PAA). After centrifugation at 800g for 30 minutes, the leukocyte layer was removed and transferred to new tubes. Subsequently, these cells were washed three times with PBS/1%BSA and re-suspended in fresh medium, consisting of RPMI medium (PAA) with 10% fetal calf serum (Invitrogen), 2.5% hepes buffer solution (PAA) and 1% penicillin/streptomycin (PAA).
Western blot. The effect of CIC on the Wnt/β-catenin pathway was analyzed by Western blot as described recently (13).
Animal studies. In our in vivo experiments we focused on the effect of CIC and lenalidomide as single agents in comparison to their combination in a murine myeloma model. All animal experiments were performed at least in duplicate with groups of six BALB/c mice (female, 6 weeks old, 25 g; Charles River, Sulzfeld, Germany). MCP11 murine myeloma cells (5×105) were injected subcutaneously into each Balb/c mouse on day 1. Mice were divided in three groups. The first group was treated with 800 μg CIC per os per day, the second group with 750 μg lenalidomide per os per day and the third group was treated with the simultaneous application of 800 μg CIC and 750 μg lenalidomide, respectively for a time frame of 60 days. This data was compared to the Overall survival and tumor growth were measured. Tumor volume was calculated as follows: volume=length×width2×0.52. Animals were sacrificed when the tumor volume reached 2000 mm3.
Statistical analysis. For statistical analysis, relative viability was expressed as the mean±standard error of the mean (SEM). Student's t-test was used for statistical analysis. A p-value <0.05 was considered significant. Statistical survival analyses were performed with the software GraphPad InStat, Version 3.0.0 (GraphPad Software, San Diego, CA, USA). The Mann-Whitney test (non-paired, non-parametric) was also applied.
Results
In our recent research, screening of the Gen-plus drug library (Microsource) of contained 960 compounds identified CIC as a Wnt/β-catenin inhibitor (13). Since the canonical Wnt signaling pathway is activated in lymphoma and myeloma cells, we further investigated if CIC is able to induce apoptosis and decrease viability of lymphoma and myeloma cell lines, respectively.
Titration of CIC. Determination of the optimal in vitro concentration of CIC revealed that 30 μM CIC was the most effective concentration to initialize cell death in both lymphoma and myeloma cell lines, without significantly deteriorating viability of normal PBMCs (data not shown). The effect of DMSO as a toxic solvent was only observed for the OPM-2 myeloma cell line in contrast to PBMC and lymphoma cell lines. OPM-2 showed a decrease of relative viability to 66.8±2.3%.
Effect of CIC on the viability of cell lines and PBMC after 72 hours of culture. Viability decreased slowly over time. After 72 h, IC50 values of CIC for the lymphoma cell lines SU-DHL-4, LAM-53 and Raji were 7 μM, 2 μM and 6 μM, respectively. In contrast, the IC50 in PBMCs was greater than 400 μM (Table I).
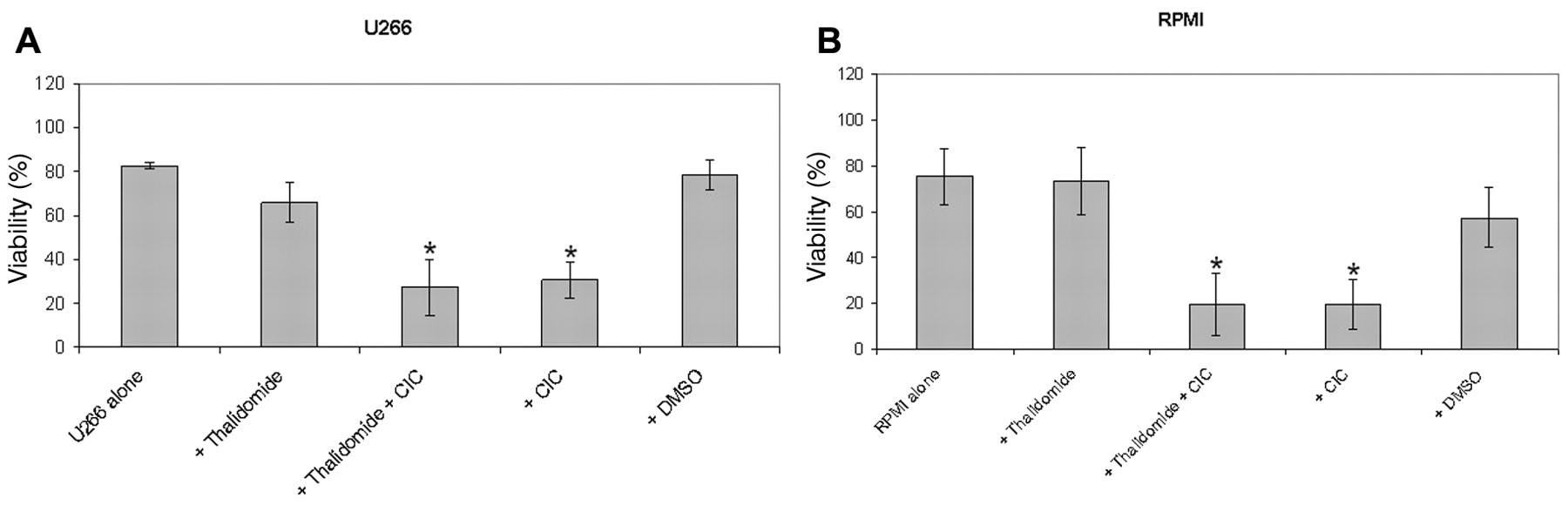
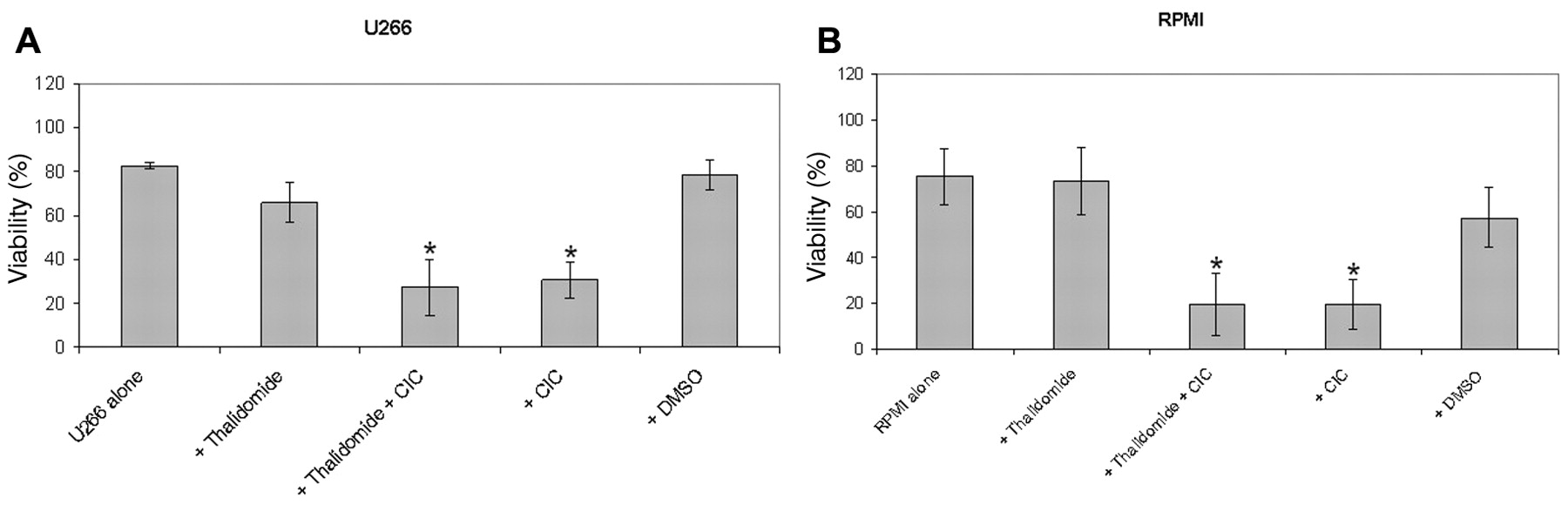
Effect of thalidomide in combination with CIC on viability of the U266 (A), RPMI (B) and OPM-2 (C) myeloma cells. 1×105 cells were cultured with different compounds for three days. The concentration of thalidomide was U266 160μM for U266, RPMI 20μM for RPMI, and OPM 40 μM for OPM cells. CIC was used at a concentration of 5 μM. Then cell viability was measured by DIOC6 staining in flow cytometry. Results represent data from three, two and five separate experiments, respectively. Separate U266 n=3, RPMI n=2, OPM n=5 experiments. Data are shown as mean±SD.
Effect of doxorubicin in combination with CIC on viability of lymphoma cell lines. In general, lymphoma patients are treated with doxorubicin and rituximab besides other drugs. Therefore, we tested these drugs in vitro in lymphoma cells in combination with CIC.
For Daudi and Raji cells, the use of doxorubicin alone led to a significant decrease in viability of Daudi and Raji cells. However, the combination of CIC plus doxorubicin did not further reduce the viability of Raji cells. Therefore, we would exclude a synergistic effect (data not shown). The use of rituximab alone led to significantly reduced viability of these cells. However, the combination of CIC plus rituximab did not further reduce the viability of Daudi and Raji cells. As with doxorubicin, we excluded a synergistic effect of this combination (data not shown).
Treatment of myeloma has changed in recent times. More and more patients are treated with novel drugs, such as bortezomib, lenalidomide and thalidomide – the latter already well known. Therefore, we tested these drugs in OPM-2 myeloma cells in combination with CIC. The use of bortezomib led to a significant decrease in the viability of OPM-2 myeloma cells. However, the combination of CIC plus bortezomib did not further reduce the viability of OPM cells. We also excluded a synergistic effect of this combination (data not shown).
Effect of thalidomide, lenalidomide and CIC on viability of myeloma cell lines. Thalidomide and CIC used alone significantly (p<0.05) reduced the viability of myeloma cell lines in vitro (Table I). A mean IC50 value for myeloma cells was determined: CIC (6 μM) was significantly more effective than lenalidomide (>200 μM) and thalidomide (>251 μM; Table I). In contrast, the effect on normal PBMC and PBL was marginal for CIC and lenalidomide, no effect was found for thalidomide (Table I).
The effect of thalidomide, lenalidomide and CIC on myeloma (OPM-2, U266, RPMI 8226, KMS 18 and MPC11) and lymphoma (LAM53, SU-DHL-4 and Raji) cell lines was assayed. PBL derived from healthy individuals were used as normal controls. 1×105 cells were cultured with each compound using various concentrations for three days. Then cell viability was measured by DIOC6 staining by flow cytometry. Results represent data from 2 to 4 separate experiments each. ND; not done.
Effect of thalidomide, lenalidomide and CIC on viability of lymphoma cell lines. Only CIC significantly (p<0.05) reduced the viability of lymphoma cell lines in vitro (Table I).
Effect of thalidomide in combination with CIC on viability of myeloma cell lines. In OPM-2, U-266 and RPMI-8226 cells, the use of thalidomide alone led to a significant decrease in viability of OPM-2, U-266 and RPMI-8226 myeloma cells. Interestingly, the combination of CIC plus thalidomide led to a further decrease of viability of OPM-2, U-266 and RPMI-8226 cells; suggesting a synergistic effect for this combination (Figure 1).
Effect of lenalidomide in combination with CIC on viability of myeloma cell lines. In OPM-2 and RPMI-8226 cells, the use of lenalidomide led to a significant decrease in viability of OPM-2 and RPMI-8226 myeloma cells. Like with thalidomide, the addition of lenalidomide to CIC led to further decreased viability of both OPM-2 and RPMI-8226 cells. These results indicate a synergistic effect for this combination (Figure 2).
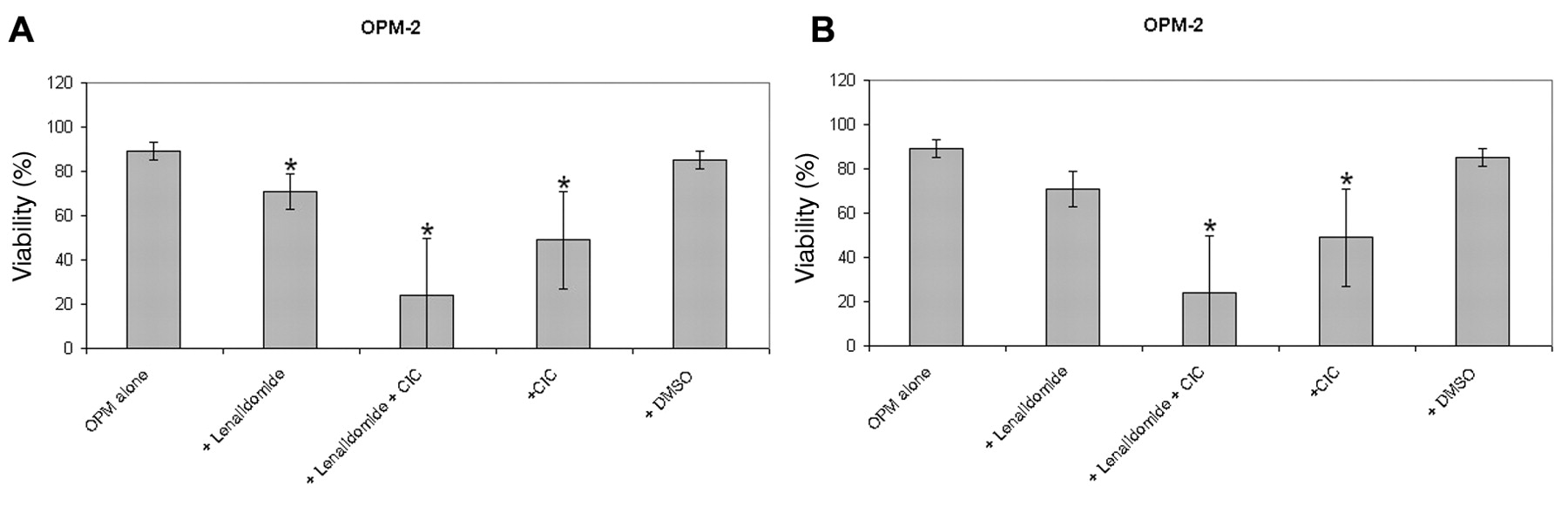
CIC synergizes with conventional therapy (lenalidomide) in OPM-2 (A) and RPMI (B) myeloma cells. 1×105 OPM cells were cultured with the different compounds for three days. The concentration of lenalidomide was 12 μM for OPM and 80 μM for RPMI cells. CIC was used at a concentration of 5 μM. Then cell viability was measured by DIOC6 staining in flow cytometry. Results represent data from eight and five OPM n=8, RPMI n=5 separate experiments, respectively. Data are shown as mean±SD.
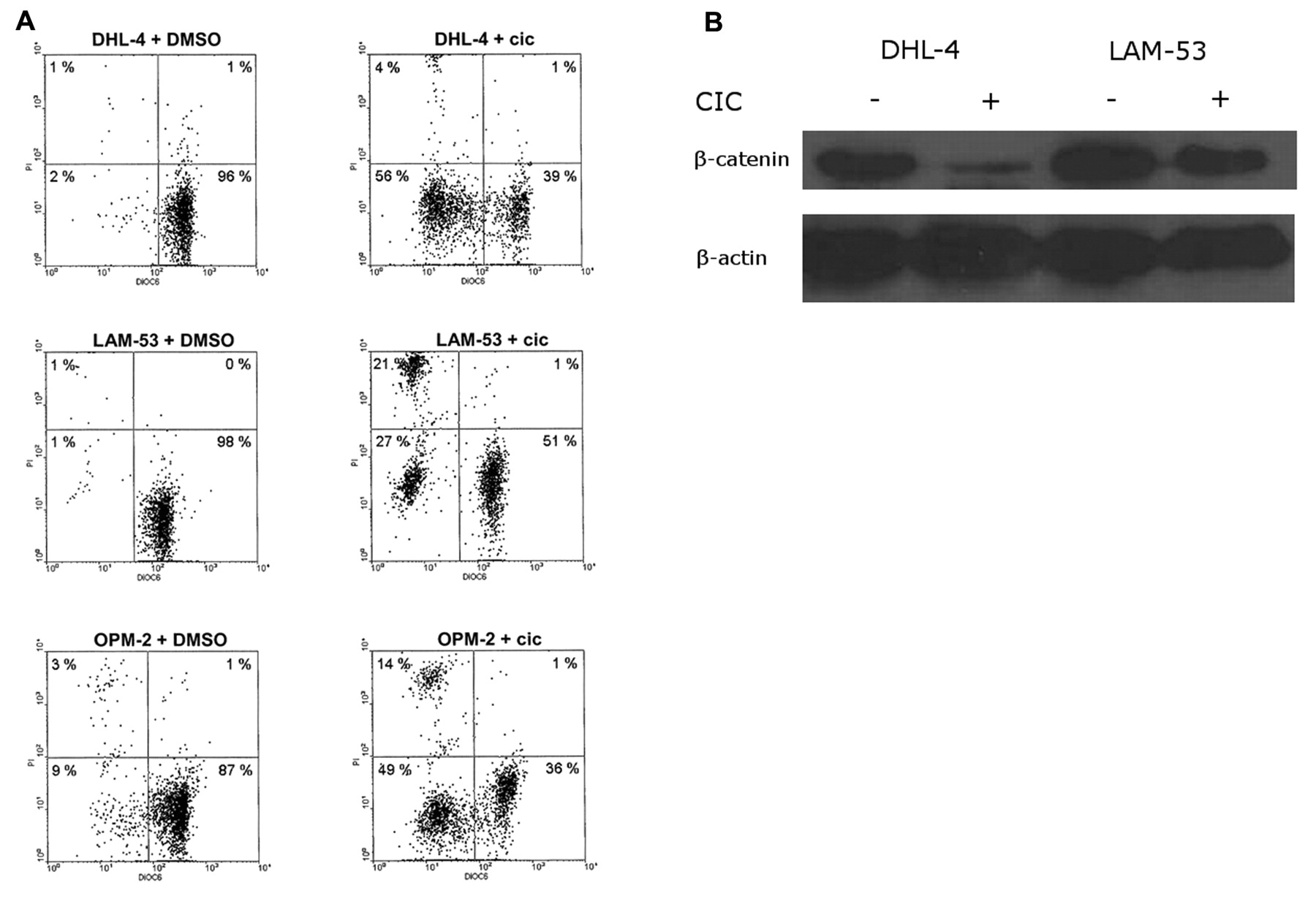
Inhibition of beta catenin by CIC in lymphoma cells. A: Flow cytometric results. CIC was used at a concentration of 10 μM. B: We analyzed the effect of CIC on the Wnt/β-catenin pathway by Western blot.
Effect of CIC on the Wnt/β-catenin pathway in lymphoma and myeloma cells. As a next step, we analyzed the effect of CIC on the Wnt/β-catenin pathway. The myeloma cell line OPM-2 did not express enough beta catenin compared to lymphoma cell lines. However, we could show that beta catenin is down regulated when CIC is added to lymphoma cells (Figure 3).
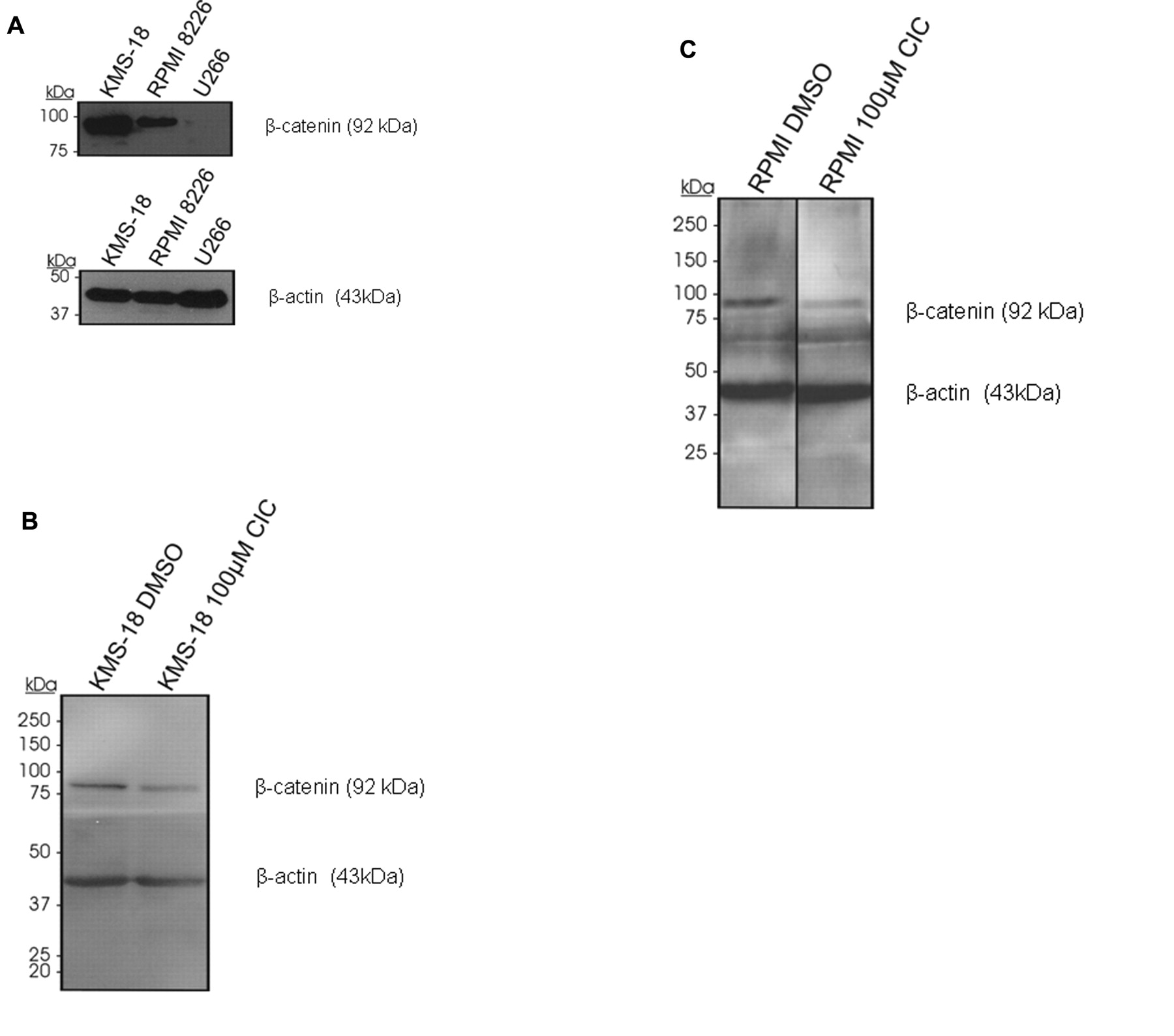
The effect of CIC on β-catenin expression in myeloma cells was tested. KMS18 and RPMI cells were found to be β-catenin positive in contrast to U266 cells being negative (Figure 4A). When exposed to CIC, β-catenin expression decreased in KMS-18 cells (Figure 4B) and slightly decreased in RPMI 8226 cells (Figure 4C).
Effect of CIC on viability of lymphoma primary cells. Preliminary data suggest that CIC inhibits the growth of primary lymphoma cells derived from patients with chronic lymphocytic lymphoma (Figure 5).

Inhibition of beta catenin by CIC in myeloma cells. A: Western Blot analysis for the detection of β-catenin in myeloma cell lines under normal conditions. The myeloma cell lines RPMI 8226, KMS-18 and U266 were analyzed according to their β-catenin expression. B: Effect of CIC on β-catenin expression by Western blot analysis. KMS-18 cells were treated with CIC for 24h, cells were lysed and then Western blot analysis was performed for β-catenin. β-actin immunoblotting served as the loading control. As primary antibodies purified mouse anti-β-catenin and β-actin were used at a dilution of 1:1000 and as secondary antibody served goat anti-mouse IgG-HRP in a dilution of 1:5000. Finally, ECL reagent was utilized for detection. C: Effect of CIC on β-catenin expression by Western blot analysis. RPMI 8226 cells were treated with CIC for 24h, cells were lysed and then Western blot analysis was performed for β-catenin. β-actin immunoblotting served as the loading control. As primary antibodies Purified mouse anti-β-catenin and β-actin were used at a dilution of 1:1000 as secondary antibody served goat anti-mouse IgG-HRP at a dilution of 1:5000. Finally, ECL reagent was utilized for detection.
Effect of lenalidomide, CIC and of the combination of lenalidomide and CIC in vivo. The effect of lenalidomide, CIC and the combination of lenalidomide and CIC was tested in vivo by subcutaneous injection of 5×105 MPC11 myeloma cells in BALB/c mice. Tumor growth was significantly reduced and overall survival was significantly increased in mice treated with CIC or CIC plus lenalidomide as compared to control mice (Figure 6A and 6B).
Discussion
The Wnt pathway is strictly regulated showing its importance in humans. It has been shown to play a critical role in the early phases of B lymphocyte development. Multiple myeloma (MM) cells, but not cells from healthy donors and patients with monoclonal gammopathy of undetermined significance or other plasma cell dyscrasias involving bone marrow express the Wnt-signaling antagonist DKK1 (14). In MM, thalidomide has been used to treat patients. Many hypotheses have been proposed to explain the molecular mechanism of thalidomide's teratogenicity, in particular regarding limb defects. Most experimental evidence in vivo has been provided for a model suggesting the generation of oxidative stress by thalidomide with subsequent down regulation of both Wnt and Akt survival pathways. In addition, transcription factors Tbx5 and Sall4 are involved in thalidomide induced molecular pathology (15). Since thalidomide down regulates the Wnt pathway and is used for treatment of myeloma patients, testing the combination of thalidomide and Wnt inhibitors became of particular interest. We used CIC, a drug that has recently been shown to induce apoptosis and down regulate the beta catenin expression in lymphoma cells. Most interestingly, we could demonstrate a synergistic effect of the combination of thalidomide and CIC in myeloma cells.

Effect of CIC on viability of lymphoma primary cells. Primary lymphoma cells derived from blood derived from patients with chronic lymphocytic lymphoma were incubated for three days with CIC at a concentration of 200 μM. Then cell viability was measured by DIOC6 staining in flow cytometry. Results represent data from two separate experiments. Data are shown as mean.
In addition to thalidomide, patients with myeloma are also being treated more frequently with the novel agents bortezomib and lenalidomide. Therefore, we tested these drugs in myeloma cells in combination with the Wnt inhibitor EA. Our research demonstrates a synergistic effect of the combination of lenalidomide/thalidomide and CIC for myeloma cells. Recently, similar results were obtained with other Wnt inhibitors (16-18). However, we failed to demonstrate such an effect for the combination of bortezomib and the Wnt inhibitors. This might be explained by the molecular relationship of thalidomide and lenalidomide, in contrast to bortezomib which belongs to a completely different class of drugs. In the lymphoma cell lines SU-DHL4 and LAM-53, the Wnt inhibitor CIC induced apoptosis through down-regulation of β-catenin, an important molecule within the Wnt pathway.
Recently, we also shown that a different drug inhibiting the wnt pathway, EA, is efficacious in primary cultures derived from patients with chronic lymphocytic leukemia (13). CIC and EA were identified as a Wnt inhibitor using a cell-based Wnt reporter assay. In vitro assays further confirmed the inhibitory effect of EA and CIC on Wnt/β-catenin signaling. Cell viability assays showed that CIC and EA selectively induced cell death in primary CLL cells. Exposure of CLL cells to EA decreased the expression of Wnt/β-catenin target genes, including LEF-1, cyclin D1 and fibronectin. Immune co-precipitation experiments demonstrated that EA could directly bind to LEF-1 protein and destabilize the LEF-1/β-catenin complex. N-acetyl-L-cysteine (NAC), which can react with the α, β-unsaturated ketone in EA, but not other anti-oxidants, prevented the drug's inhibition of Wnt/β-catenin activation and its ability to induce apoptosis in CLL cells.

In vivo efficacy of CIC, lenalidomide (R) and of the combination of lenalidomide and CIC. The effect of lenalidomide, CIC and of the combination of lenalidomide and CIC was tested in vivo. BALB/c mice were injected with 5×105 MPC11 myeloma cells subcutaneously. (A) Tumor growth and (B) overall survival were measured. Six animals were assayed per group. Control: mice without treatment.
Our results are in accordance with a recent report of Sukhdeo et al. demonstrating that the canonical Wnt signaling pathway is activated in MM through constitutively active β-catenin (19). Interestingly, we demonstrated a synergistic effect of the combination of lenalidomide and Wnt inhibitors in myeloma cells. This observation might lead to novel treatment options being developed for patients with MM.
In conclusion, our results reveal a significant induction of apoptosis by CIC in both lymphoma and myeloma cells. Combined with our previous results, our data suggest that CIC can inhibit Wnt/β-catenin signaling in both lymphoma and myeloma cell lines.
Acknowledgements
We kindly acknowledge the support of T. Kipps, La Jolla and P. Brossart, Bonn, Germany, the excellent technical expertise of Petra Alpmann and Sabine Blaum-Feder and Axel Glasmacher, Bonn for improving the language of our manuscript.
This study was kindly supported in part by the Tumorinitiative von Haller e.V., Bonn and by the Deutsche Krebshilfe, Bonn, Germany.
- Received May 13, 2011.
- Revision received July 14, 2011.
- Accepted July 15, 2011.
- Copyright © 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Targeting Histone Demethylases in MYC-Driven Neuroblastomas with Ciclopirox
- {beta}-Catenin Inhibitor BC2059 Is Efficacious as Monotherapy or in Combination with Proteasome Inhibitor Bortezomib in Multiple Myeloma
- In Vitro Apoptosis Induction by Fenofibrate in Lymphoma and Multiple Myeloma
- Griseofulvin Efficiently Induces Apoptosis in In Vitro Treatment of Lymphoma and Multiple Myeloma
- Targeting Prostate Cancer with a Combination of WNT Inhibitors and a Bi-functional Peptide
- Clofibrate Demonstrates Efficacy in In Vitro Treatment of Lymphoma and Multiple Myeloma
- In Vitro Efficacy of Naftifine Against Lymphoma and Multiple Myeloma
- Flunarizine Exhibits In Vitro Efficacy Against Lymphoma and Multiple Myeloma Cells
- In Vitro Efficacy of Cinnarizine Against Lymphoma and Multiple Myeloma
- Effect of Wnt Inhibitors in Pancreatic Cancer
- Targeting the Wnt/Beta-Catenin Pathway in Renal Cell Carcinoma
- Blocking eIF5A Modification in Cervical Cancer Cells Alters the Expression of Cancer-Related Genes and Suppresses Cell Proliferation
- Targeting the Wnt/Beta-Catenin Pathway in Multiple Myeloma
- Targeting Renal Cancer with a Combination of WNT Inhibitors and a Bi-Functional Peptide
- Antagonistic Effect of Small-molecule Inhibitors of Wnt/{beta}-catenin in Multiple Myeloma