


Abstract
Background: The MeOH extracts from aerial part and roots of Rhinacanthus nasutus were investigated for new biological activities. Materials and Methods: The MeOH extract of the root was stepwise separated by organic solvents into n-hexane, EtOAc, n-BuOH and H2O layer fractions. Cytotoxic activity against human tumor and normal cells was determined by the MTT method. Nitric oxide (NO) was determined by the Griess method. Osteoclastogenesis was monitored by tartrate-resistant acid phosphatase (TRAP) activity. Results: The MeOH extract of the root showed much higher tumor-specific cytotoxicity than that of the aerial part. The EtOAc fraction of the root showed the highest tumor-specific cytotoxicity, followed by the n-BuOH, n-hexane and H2O fractions. None of the four fractions protected the cells from the cytotoxicity of UV irradiation. The n-BuOH fraction not only stimulated NO production by mouse macrophage-like RAW264.7 cells, but also inhibited the lipopolysaccharide (LPS)-stimulated NO production. The EtOAc fraction inhibited the receptor activator for nuclear factor-κB ligand (RANKL)-stimulated osteoclastogenesis of the RAW264.7 cells most potently, followed by the n-hexane, n-BuOH and H2O fractions. The n-BuOH fraction slightly, but significantly stimulated osteoclastogenesis. Conclusion: Antitumor and macrophage/osteoclast-modulating substances are enriched in EtOAc and n-BuOH fractions of MeOH extract of R. nasutus roots.
The shrub Rhinacanthus nasutus (L.) Kurz (Acanthaceae) (Figure 1) is found in Southeast Asian countries. The root and aerial part of R. nasutus contain naphthoquinone esters such as rhinacanthins C, D, N and Q that display apoptosis-inducing activity (1), in vitro (2-4) and in vivo (2) antitumor and antiviral activity (5), antiallergy activity (6) and anti-inflammatory activity (7, 8) and are used for the treatment of pneumonia, diabetes, hypertension and skin diseases (9). In order to search for new biological activities of the components of R. nasutus root, the MeOH extract of this plant was roughly separated into four fractions by partition with organic solvents and their possible biological activity including tumor-specific cytotoxicity, UV protection effect, inhibition of lipopolysaccharide (LPS)-induced macrophage activation and inhibition of receptor activator for nuclear factor-κB ligand (RANKL)-induced osteoclastogenesis was tested.
Materials and Methods
Materials. The following chemicals and reagents were obtained from the indicated companies: Dulbecco's modified Eagle's medium (DMEM) (Invitrogen, Carlsbad, CA, USA); fetal bovine serum (FBS) (SAFC Biosciences, St. Louis, MO, USA); RPMI-1640 medium, α-minimal essential medium (α-MEM), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and LPS from Escherichia coli (serotype 0111:B4) (Sigma-Aldrich, St. Louis, MO, USA); dimethyl sulfoxide (DMSO) (Wako Pure Chemical, Osaka, Japan) and RANKL (R&D Systems, Minneapolis, MN, USA).
Extracts and sequential fractionation with organic solvents. The aerial part (stem and leaf) and root of R. nasutus were supplied from the Chiayi Grass-Produce Cooperation Farm (Chiayi County, Taiwan, China) during the autumn of 2009. This specimen was identified by one of the authors (Professor Dr. Y. Shirataki) and also a voucher specimen (#201006060) is deposited in the Medicinal Plant Garden of Josai University.
The air-dried aerial parts (1.74 kg) and roots (1.18 kg) of R. nasutus were extracted three times with MeOH under reflux for 3 hours respectively. The three MeOH solutions were combined and evaporated to dryness to give 171 g (aerial parts) and 83.1g (roots) of extract, respectively. The root extract was suspended in MeOH (0.75 l) and H2O (75 ml), then partitioned with n-hexane (0.75, 0.38, 0.38 l). The aqueous MeOH layer solution was concentrated and resuspended in H2O (1.4 l), then extracted with EtOAc (1.4, 0.7, 0.7 l) and n-BuOH (1.2, 0.5, 0.5 l), successively (Figure 2). The n-hexane and EtOAc layer solutions were dried over anhydrous Na2SO4 and evaporated in vacuo to give n-hexane (6.55 g) and EtOAc (17.0 g) fractions, respectively. The n-BuOH and part of the aqueous layer solutions were also evaporated in vacuo to yield n-BuOH (11.7 g) and H2O fractions, respectively.

Rhinacanthus nasutus (L). Kurz.
Cell culture. Human promyelocytic leukemia HL-60 cells were provided by Professor K. Nakaya, Showa University, Japan. Human oral squamous cell carcinoma cell lines (HSC-2, HSC-3, HSC-4) were provided by Professor M. Nagumo, Showa University. Normal human oral cells, gingival fibroblast (HGF), pulp cells (HPC) and periodontal ligament fibroblast (HPLF), were prepared from periodontal tissues, according to the guideline of the Intramural Ethic Committee (No. A0808), after obtaining the informed consent from a 12-year-old patient at the Meikai University Hospital. Since normal oral cells have a limited lifespan of 43-47 population doubling levels (PDL) (10), they were used at 8-15 PDL. Mouse macrophage-like RAW264.7 cells (11) were purchased from Dainippon Sumitomo Pharma (Osaka, Japan). The HL-60 cells were cultured in RPMI-1640 medium supplemented with 10% heat-inactivated FBS, 100 U/ml penicillin G and 100 μg/ml streptomycin sulfate under a humidified 5% CO2 atmosphere. The other cells were cultured in DMEM supplemented with 10% heat-inactivated FBS, 100 U/ml penicillin G and 100 μg/ml streptomycin sulfate. The normal cells were detached by 0.25% trypsin-0.025% EDTA-2Na in phosphate-buffered saline without Mg2+ and Ca2+ (PBS(−)) and subcultured at a 1:4 split ratio once a week, with a medium change in between the subcultures. The five tumor cell lines were similarly trypsinized and subcultured.
Assay for cytotoxic activity. The cells (3×103/well, 0.1 ml/well) were seeded in 96-microwell plates (Becton Dickenson, Franklin Lakes, NJ, USA) and incubated for 48 hours to allow cell attachment. Near-confluent cells were treated for 48 hours with different concentrations of the test compounds in fresh medium. The relative viable cell number of adherent cells (except for HL-60 cells) was then determined by the MTT method. In brief, control and sample-treated cells were incubated for 4 hours with 0.2 mg/ml of MTT in the culture medium. After removing the medium, the reaction product, formazan, was extracted with DMSO and the absorbance (the relative viable cell number) was measured at 540 nm by a microplate reader (Multiskan Bichromatic Labsystems, Helsinki, Finland). The viability of the suspended cells, i.e. HL-60, was determined by cell counting with a hemocytometer after staining with 0.15% trypan blue. The 50% cytotoxic concentration (CC50) was determined from the dose–response curve. The tumor-specificity index (TS) was calculated by the following equation: TS=mean CC50 (normal cells)/mean CC50 (all tumor cell lines).

Extraction and fractionation of R. nasutus root with organic solvents.
Assay for UV protection. HSC-2 cells, that showed the highest sensitivity against UV irradiation among 6 adherent cell lines tested (10), were inoculated into 96-microwell plates (3×103/well, 0.1 ml/well) and incubated for 48 hours to allow cell attachment. The culture supernatant was replaced with PBS(−), placed at 21 cm distance from a UV lamp (wavelength: 253.7 nm) and exposed to UV irradiation (6 J/m2/min) for 1 min. The PBS was replaced with fresh DMEM containing 10% FBS, and the cells were incubated for a further 48 hours to determine the relative viable cell number by the MTT method, as described above.
Effect on NO production by macrophage-like cells. The RAW264.7 cells (6×103/well, 0.1 ml/well) were inoculated into 96-microwell plates, and incubated for 24 hours with or without LPS (100 ng/ml). LPS stimulates NO production via the induction of inducible NO synthase (iNOS) expression (13). The medium was then replaced with phenol red-free DMEM medium containing 10% FBS and the indicated concentration of the test samples. After incubation for 24 hours, the NO released into the culture supernatant was measured by the Griess method. From the dose–response curve, the 50% effective concentration (EC50) was calculated. The attached cells were stained with MTT reagent to determine the CC50, as described above. The selectivity index (SI) for the inhibition of NO production was determined by the following equation: SI=CC50/EC50 (12).
Osteoclastogenesis in cell culture. The RAW264.7 cells were suspended in α-MEM supplemented with 5% FBS and seeded at 2×103 cells/well in 96-well plates (0.1 ml/well) in the presence or absence of RANKL (10 ng/ml), with or without the test samples. After 4 days in culture, tartrate-resistant acid phosphatase (TRAP) activity of the medium was determined and TRAP staining of the cells was performed.
Cytotoxic activity of aerial part and root extracts of R. nasutus against cultured human tumor and normal cells.
Measurement of TRAP activity and TRAP-staining. Culture media (30 μl) were incubated for 30 min at 37°C with 30 μl of 600 mM sodium acetate buffer (pH 5.5) containing L-ascorbic acid (17.6 mg/ml), sodium tartrate dehydrate (9.2 mg/ml), disodium 4-nitrophenylphosphate (3.6 mg/ml), Triton X-100 (0.3%), EDTA (6 mM) and NaCl (600 mM). The reaction was terminated by the addition of 30 μl of NaOH (300 mM) and the absorbance at 405 nm was measured by a microplate reader. TRAP histochemical staining of the cells was performed using a leukocyte acid phosphatase kit (Sigma-Aldrich). The cultured cells were fixed with 100% MeOH for 1 min at room temperature, air-dried and then stained for TRAP activity.
Statistical analysis. The difference between two groups was evaluated by Student's t-test.
Results
Cytotoxicity of extracts and fractions. The MeOH extract of the aerial part of R. nasutus showed very weak cytotoxicity against both human tumor HSC-2, HSC-3, HSC-4 and HL-60 and normal cells (HGF, HPC and HPLF), yielding a very low level of tumor specificity (TS=1.1, 1.0; Experiments I and II in Table I).
On the other hand, the MeOH extract of the root of R. nasutus showed much higher cytotoxicity against human tumor cell lines, yielding much higher tumor specificity (TS≥4.6, >2.2; Table I). Among the four fractions prepared from the MeOH extract of the root (Figure 2), the EtOAc fraction showed the highest tumor-specific cytotoxicity (TS=7.0 and 19, Experiments I and II, respectively), followed by the n-BuOH (TS=5.5, 5.0), n-hexane (TS≥4.8, 5.0) and H2O fractions (TS=><1.0, ><1.0) (Table I).
Effect on NO production in LPS-activated RAW264.7 cells of aerial part and root extracts of R. nasutus.
Effect on UV cytotoxicity. One-minute exposure of HSC-2 cells to UV irradiation induced irreversible cell death after 48 hours' incubation (Figure 3). The MeOH extract (Figure 3A), and all four fractions (Figure 3B-E) did not protect the cells from UV induced cytotoxicity.
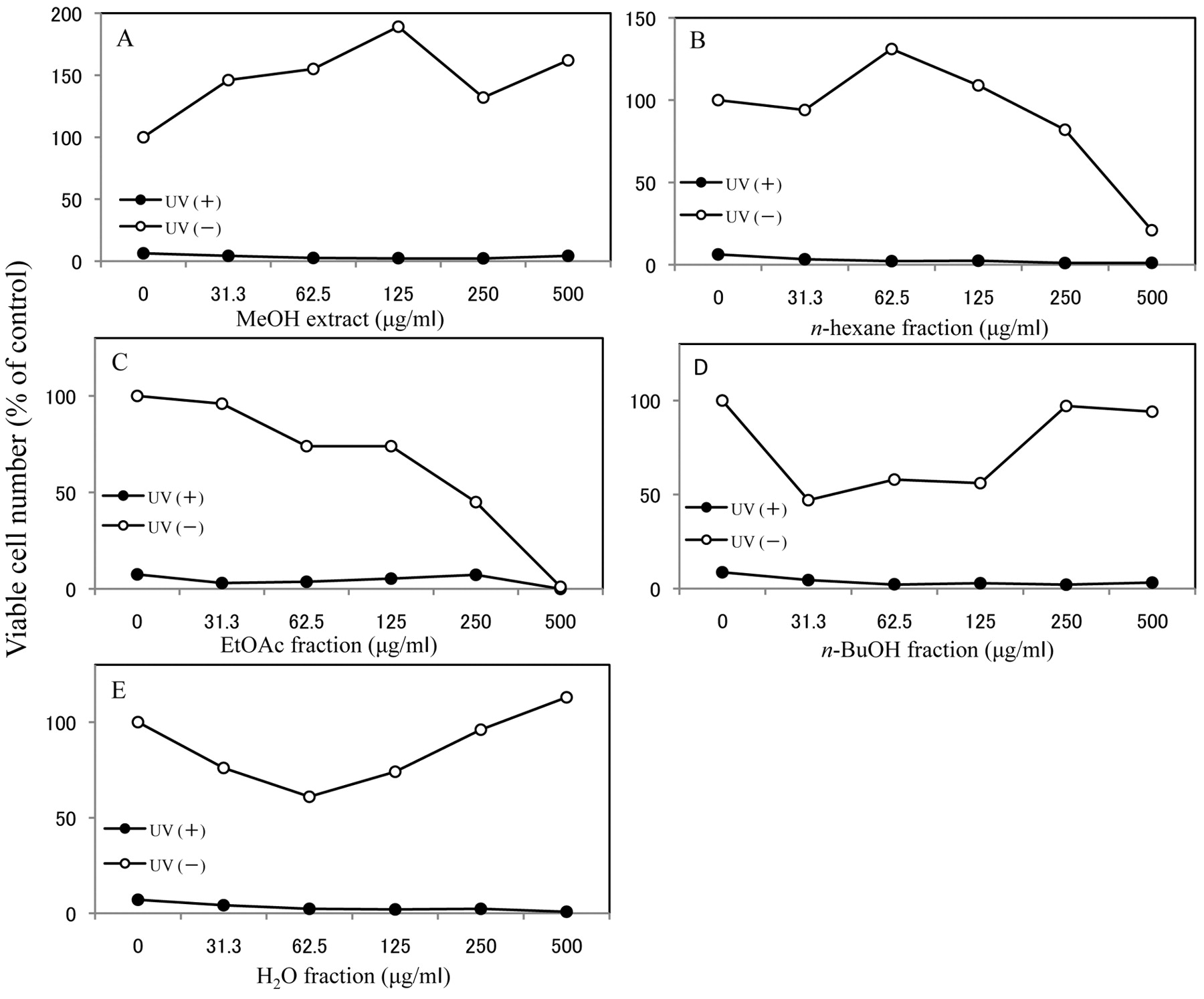
Effect of R. nasutus root on UV-induced cytotoxicity extracts. HSC-2 cells were exposed to UV irradiation (6 J/m2/min, 1 min) in the absence or presence of MeOH extract of R. nasutus roots (A), or its n-hexane (B), EtOAc (C), n-BuOH (D) or H2O (E) fraction. After incubation for 48 hours in fresh medium, the viable cell number was determined by MTT method and expressed as % of control that had not been exposed to UV irradiation. Mean±S.D. of triplicate assays.
NO production by macrophage-like cells. Treatment of the RAW264.7 cells with LPS elevated the extracellular concentration of NO from the background level (2 μM) to up to 24 μM (Figure 4).
The MeOH extract of the aerial part dose-dependently inhibited the LPS-stimulated NO production (EC50=34.6 μg/ml, CC50=310 μg/ml, SI=9.0; Figure 4A, Table II). The MeOH extract of the root also showed comparable inhibitory activity (EC50=5.16 μg/ml, CC50≥31.3 μg/ml, SI≥6.1; Figure 4B, Table II)
Among the four fractions of the MeOH extract of the root, the n-BuOH fraction (Figure 4E) showed the highest inhibitory activity of NO production (SI≥20), followed by the EtOAc (SI=3.1; Figure 4D), n-hexane (SI=2.2; Figure 4C) and H2O fractions (SI=1.3; Figure 4F) (Table II). It should be noted that the n-BuOH fraction alone significantly stimulated NO production by the RAW264.7 cells without LPS, suggesting the presence of both stimulators (Figure 4E) and inhibitors in the n-BuOH fraction.
Osteoclastogenesis. Treatment of the RAW264.7 cells with RANKL stimulated the production of osteoclasts (Figure 5A). Among the four fractions of the MeOH extract of the root, the n-BuOH fraction alone slightly, but significantly (p<0.01) stimulated osteoclastogenesis (Figure 5B, upper panel).
The MeOH extract from the aerial part of R. nasutus (6.4-160 μg/ml) did not inhibit the RANKL-stimulated osteoclastogenesis, whereas the MeOH extract from the root significantly inhibited the RANKL-stimulated osteoclastogenesis at as little as 10 μg/ml (Figure 5B, lower panel). Among the four fractions of the MeOH extract of the root, the EtOAc fraction showed the highest inhibitory activity, followed by the n-hexane, n-BuOH and H2O fractions. These data suggested the presence of both stimulators and inhibitors of osteoclastogenesis in the n-BuOH fraction.

Effect of extracts of R. nasutus on NO production in RAW264.7 cells. Cells were incubated for 24 hours with MeOH extract of aerial part (A) or root (B) of R. nasutus, or the n-hexane (C), EtOAc (D), n-BuOH (E) and H2O (F) fraction of the root in the presence or absence of LPS (100 ng/ml). The viable cell number (●: +LPS; ○: −LPS) and extracellular NO concentration (▪: +LPS; □: − LPS) was then determined by MTT and Griess methods, respectively. Mean ± S.D. of triplicate assays.
Discussion
The present study demonstrated, for the first time, various new biological activities of MeOH extracts of R. nasutus root although none of the extracts of fractions protected the cells from UV cytotoxicity in contrast to the highly efficient UV protective effect of sodium ascorbate reported previously (10). Among the four different fractions of the MeOH extract, the EtOAc fraction showed higher cytotoxicity against several human tumor cell lines (three oral squamous cell carcinomas and a promyelocytic leukemia) than against three human normal oral cells, with a TS value of 7.0 to 19 (Table I). The type of cell death induced by the EtOAc fraction in the carcinoma cells remains to be investigated, since it depends on both the type of inducer and target cell (14). The EtOAc fraction also significantly (p<0.01) inhibited osteoclastogenesis at extremely low concentrations (2 μg/ml).
The present study also demonstrated that the n-BuOH fraction of the MeOH extract of the roots stimulated NO production and osteoclastogenesis in the RAW264.7 macrophage-like cells, but conversely inhibited the LPS-stimulated NO production and RANKL-induced osteoclastogenesis in the same cells. This suggested the presence of both stimulators and inhibitors for macrophages and osteoclasts in the n-BuOH fraction. Further purification is crucial to separate the active principle from the cytotoxic substances. We have recently separated the n-hexane fraction by silica gel column chromatography into a flow-through fraction and adsorbed fractions which were then eluted stepwise with n-hexane-EtOAc (9:1), n-hexane-EtOAc (4:1), n-hexane-EtOAc (7:3) and n-hexane-EtOAc (1:1) in order to separate the active principle(s) from the cytotoxic substance(s). Rhinacanthin C was isolated from the n-hexane-EtOAc (9:1) fraction (15). The identification and biological activities of active principle(s) in other fractions will be reported elsewhere.

Effects of aerial part and root extracts of R. nasutus on osteoclast differentiation. RAW264.7 cells were cultured for 4 days in the presence or absence of RANKL with MeOH extract of aerial part or root, or n-hexane, EtOAc, n-BuOH or H2O fraction of root extract. (A) TRAP staining, representative range of morphological appearance. (B) TRAP activity of the medium. All samples contained 0.2% DMSO as a vehicle control. Mean±SD of quadruplicate assays. **p<0.01, compared to control.
Acknowledgements
We are grateful to Ms. Lu Grace, Taiwan, for her kind supply of R. nasutus.
- Received November 27, 2010.
- Revision received January 27, 2011.
- Accepted January 28, 2011.
- Copyright © 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}