


Abstract
Background/Aim: The aim of the present study was to develop the basis for the use of surfactants in the treatment of pulmonary tuberculosis (TB). Bacteria are surrounded by a thick lipid coat primarily consisting of trehalose dimycolate (TDM) and, consequently, are well shielded from the immune system's response and antibiotics. This protective barrier was removed by exposing the bacteria to certain surfactants. Materials and Methods: Dodecyl maltoside (DDM) and octyl glucoside (OG) were utilized as non-toxic surfactants. Results: Electron microscopy (EM) studies revealed that aggregated bacteria were also covered with excessive TDM which exacerbate the treatment efforts. Light and EM studies demonstrated that DDM and OG disperse the aggregated bacteria and are bactericidal. Conclusion: The studies presented here establish that certain surfactants are proficient in removing MTB's shield and, because they are well known as cell permeabilizing agents, they may also enhance the effectiveness of antibiotics and the immune system's response in the treatment of pulmonary TB.
- Mycobacterium tuberculosis
- trehalose dimycolate
- octyl β-D-glucopyranoside
- n-dodecyl β-D-maltoside
- bacterial coat
Abbreviations: MTB, M. Tuberculosis; MDR-TB, multidrug resistance TB; XDR-TB, extreme drug resistance TB; TDM, trehalase dimycolate; OG, octyl β-D-glucopyranoside; DDM, n-dodecyl β-D maltoside; BAL, bronchial alveolar lavage; EM, electron microscopy; CMC, critical micelle concentration; DPPC, dipalmitoyl phosphatidylcholine; PG, phosphatidyl glycerol.
Mycobacterium tuberculosis (MTB) has been and remains as one of the most difficult bacterial pathogens to treat. Its pervasiveness is expressed in 8 million new cases reported worldwide which result in an estimated two million deaths per year. Even though a quadripartite antibiotic regimen may offer successful treatment, its six to eight months' duration is expensive and the accumulated toxicity of the drugs often result in poor patient compliance (1). The compliance issue has been further exacerbated by the emergence of multidrug-resistant TB (MDR-TB) and extreme drug-resistant TB (XDR-TB) which are major threats to world health. The World Health Organization estimates that there are 450,000 new cases of the MDR-TB each year and this number is expected to increase because of the lack of new therapeutic options (1).
A significant contributor to its tenacity is a relatively thick, waxy, hard lipid coat primarily consisting of trehalose dimycolate (TDM) which very effectively protects the bacteria from the immune system and antibiotics. In this regard, H. Bloch characterized this lipid coat (2) and found it was produced in excessive quantities and contributed to the inflammatory response associated with pulmonary MTB (3). The significance of this barrier to the survival of mycobacteria has been demonstrated by the study of mutants that have reduced levels of TDM and, consequently, exhibit reduced antibiotic resistance and are not pathogenic (4).
But its armor is MTB's Achilles' heel. Accordingly, we propose a direct, rapid and localized approach to treating pulmonary MTB in which the bacteria are stripped of their shield by direct exposure to surfactants introduced into the lung. In this regard, surfactants have been established since the early 1960s as an efficient method of disrupting lipid bilayers and have a long history of use as the principal means used to extract and isolate proteins from cellular membranes. Thus, in this study, the proficiency of certain surfactants to disrupt MTB membrane and their toxicity in the lungs of mice has been evaluated. The surfactants exhibit no significant toxicity under conditions that are bactericidal to MTB. The electron and light microscopy studies of MTB presented here are in further support of our contention that certain surfactants offer a novel, fast acting, targeted treatment modality that is well suited for breaching the bacteria's armor and exposing them to antibiotics and the immune response for an uncharacteristically rapid clearance of the disease.
Materials and Methods
Surfactants. Octyl β-D-glucopyranoside (OG) and n-dodecyl β-D maltoside (DDM) (Sigma-Aldrich, Dallas, TX, USA) were weighed and added to 0.15 M NaCl the day of use to obtain 1%, 0.2%, 0.1% (w/v) stock solutions.
Evaluation of surfactant toxicity. Female (C57BL/6) mice (6-9wk of age and ~20g from Jackson laboratory, USA) were utilized for pulmonary aerosolization. Mice were anesthetized with 150-250 μl of 2.5% Avertin prior to aerosolization and 500 μl was administered before exsanguination. Animal experiments were performed as per the guidelines developed and approved by the Animal Welfare committee, University of Texas Health Science Center at Houston.
Pulmonary aerosolization. A non-invasive aerosol delivery to mice lungs of 25 μl of surfactant in 0.15M NaCl was administered utilizing a Penn-Century syringe and shaft (5). Control mice were administered 25 μl of saline without surfactant. The aerosolization of surfactant solutions required approximately one second. Four mice in each group were treated two days in succession weekly for three weeks and once in the fourth week. The fifth week, mice were sacrificed and their lungs, spleen and livers were removed for histological evaluation. The tissues were fixed in buffered 10% formalin and tissue sections were prepared on a microscope slide by the Department of Pathology Histology Laboratory and stained with hematoxylin and eosin according to standard protocols.
Bronchial alveolar lavage (BAL). After exsanguination, a blunt 21-gauge needle was secured with a thread in the trachea and the lungs were lavaged with 1-2 ml of phosphate buffered saline. The total cell count was determined in the lavage using a hemocytometer and the lavage was cytospum onto microscope slides and stained by the Diff-Quick procedure for differential cell counts. Total protein was also estimated in the lavage.
Electron microscopy. The Erdman strain of MTB was grown in Middlebrook 7H9 media containing 0.05% Tween 80 for approximately two weeks before sampling. The medium was supplemented with 0.2% glycerol, 0.5% bovine serum albumin and 0.2% glucose. The preparation was vortexed briefly to disperse clumps then a volume of ~100 μl was centrifuged in Eppendorf tubes for 60 seconds. The precipitate was suspended in PBS and 6 μl added to carbon-coated formvar grids. After wicking with bibulous paper, the specimen was stained 4× with 6 μl of 0.25% methylamine tungstate containing 10 μg/ml Bacitracin and air dried. Prior to their removal from the biosafety laboratory, the grids were heated for 20 min at ~ 60°C on a heat block. The bacteria were also grown in media without surfactant and the resulting clumps were sonicated 30 s or vortexed with glass beads for 5 min to disperse the preparation. A mutant H37RvΔ KasB mTB strain grown in the media described above was also examined. The mycolic acid chains of the mutant are 2-4 C atoms shorter than in the Erdman strain (6). Images were recorded with a JEOL JEM 1200 electron microscope operated at 100 KV using conventional irradiation procedures with an under focus of ~2 μm.
Light microscopy. The Erdman strain was grown as described above in Middlebrook media without Tween in order to obtain the bacteria in the corded state. A sample was vortexed for 15 s with and without 1% OG or DDM in PBS then sampled at the indicated time for imaging in a light microscope after staining using the Kinyon procedure. The volume was chosen so that the image recorded at ×2 was representative of the entire specimen under view.
Results
Electron microscopy. Early electron microscopy (EM) studies of MTB often displayed thin sections derived from the embedded bacteria. Depending on the orientation of the plane of section, their shapes varied between round to oval (7). Images of individual bacteria reveal a hotdog-like shape ~0.5 μm in diameter and 2-5 μm in length (Figure 1A). The bacteria were grown in the presence of Tween 80, which partially limits aggregation and minimizes the formation of clumps. For practical considerations, the bacteria are usually grown in the presence of a surfactant. The hydrophobic effect (8) related to their TDM outer coat drives their aggregation and an extensive study of the role of TDM in the formation of the chorded state of MTB and its role in pathogenesis has been reviewed (9). Figure 1A illustrates the varied length and nearly constant diameter of MTB and also shows that the bacteria divide by pinching off at their ends. The characteristic dark, circular spots probably represent a region of concentrated nucleic acid (nucleoids).
A seldom seen image (Figure 1B) from the same growth medium is a single bacterium that represents an extraordinary display of the TDM coat (white) that has partially broken loose from the bacterium. The hardness of the coat is apparent because it maintains the contour of the bacterium and is consistent with the TDM phase transition temperature of 65°C of M. chelonae and MTB measured by differential scanning colorimetry (10).
A typical representation of bacteria in this media show that they have a “fisherman's” net morphology that often extends 40 μm or more (Figure 2). The side by side arrangement of the bacteria maximizes their contacts and favors their convoluted rope-like appearance. The edge of the aggregate has a foam-like appearance which may be representative of excessive TDM that has formed liposomes and micelles.
The aggregated bacteria have a more condensed arrangement when grown in media without surfactant (Figures 3A and B). Instead of the net-like appearance, the bacteria are more condensed and form amorphous structures which are also very large, greater than 10 μm. Sonication of this preparation results in rupture of the TDM outer shell and exposure of the encapsulated bacteria (Figure 3B). The shell breaks off like shattered glass, and the resulting fragments are so firm that they maintain the imprint of the bacteria, similarly to Figure 1B. When stirred with a glass rod, they have the consistency of fine sand and must be dispersed by vortexing with glass beads or sonication to reduce them to a size amenable for imaging by EM. It appears that TDM covers the individual bacteria in the aggregate, which is further enclosed within an external blanket of TDM. These EM images offer a new perspective of Bloch and Noll's finding that the bacteria are associated with an excessive quantity of TDM (3). This condensed, aggregated state of the bacteria is probably representative of their morphology seen with a light microscope in their corded state (11) and in the cavities of patients with cavitary TB (see below).

A: The morphology of M. tuberculosis (MTB). The bacteria have variable length with constant width (~0.5 μm) and display several characteristic round spots along their length which are probably nucleiods. Tween 80 in the growth media significantly limits the display of TDM (white) on their exterior surface. B: A bacterium which maintained a more extensive TDM coat (white) which has been partially dislodged but remarkably maintains the bacterium's shape. This image illustrates the rigidity of the TDM outer layer.
Light microscopy. We examined the effect of DDM and OG on the aggregated state of the bacteria. These surfactants were selected because they are among the most commonly used for the purpose of disrupting cellular membranes in order to isolate their associated proteins. In part, their proficiency is related to their high critical micelle concentration (CMC): value of 10−2 – 10−3 M. The bacteria were grown in media without Tween 80 and primarily reside in the corded state (Figures 4A and B; depicted in greater detail in Figures 3A and B. After exposure of the preparation for 30 min to 1% OG (Figures 4C and D) or 1% DDM for 2 h (Figures 4G and H) most of the cords have disappeared and there is a significant diminution of the bacteria which is more pronounced after a 24-h exposure to OG. The clearance of the bacteria indicates that OG and DDM disrupt the MTB structure and are bactericidal. Conversely, MTB treated 24 h with 1% dipalmitoylphosphatidylcholine (DPPC) showed no change. DPPC is the major component of human lung surfactants. However, there was a significant reduction in the aggregated state after a one-week exposure to DPPC (data not shown).

Representative field of the MTB grown in media containing Tween (surfactant). Most of the bacteria are associated side by side in an enormous net-like aggregate. The bubbles and rod-like shapes associated with their edge probably represent liposomes and micelles comprised of TDM. Note the bacteria display the spots seen in Figure 1, indicating that they lack their TDM outer coat seen in Figure 1B.
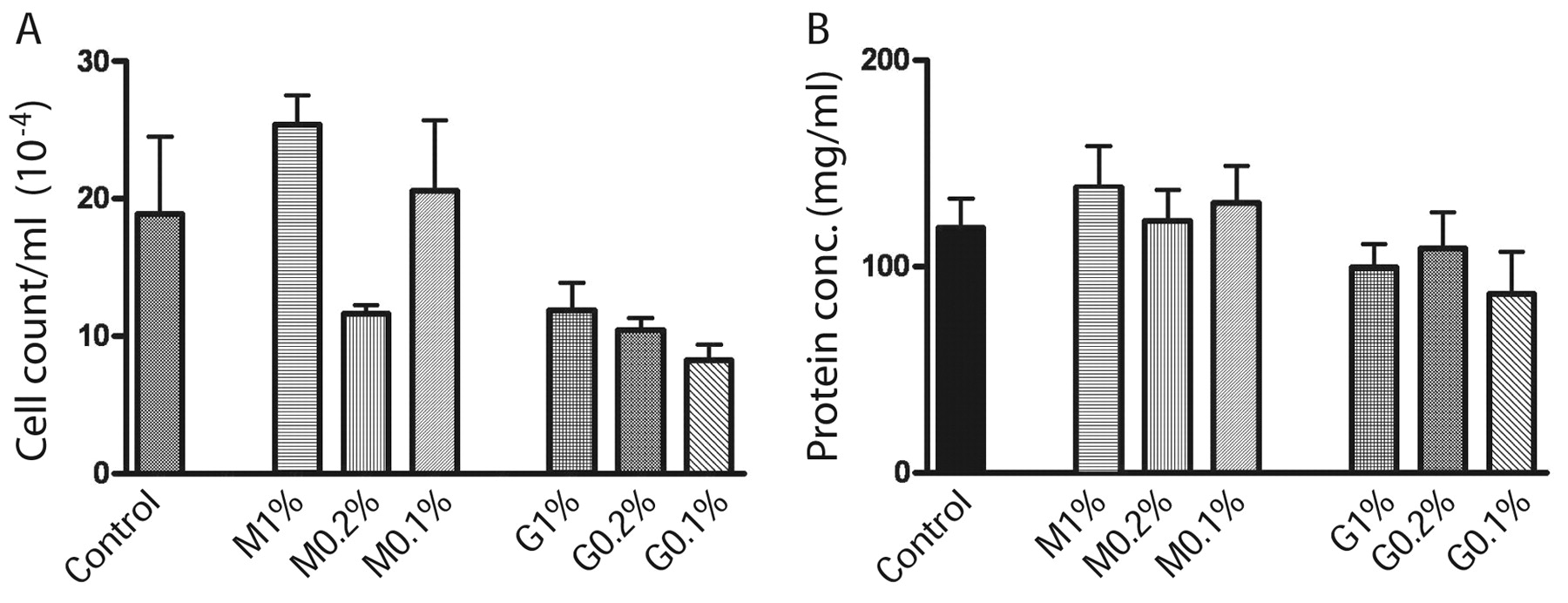
Toxicity evaluation of OG and DDM. In comparative studies of surfactant toxicity utilizing cell cultures, OG and DDM were determined to be the least toxic, which is partially related to their lack of charge (12). We evaluated their toxicity when aerosolized into lungs of mice over a concentration range of 0.1-1% (w/v). Recall, 1% surfactants mostly eliminated the aggregated state of MTBs and is bactericidal (Figure 4).
Our use of the non-invasive endotrachial aerosolization of mice lungs has a significant advantage over exposing the mice in an aerosolization chamber because the drug is precisely administered only to the lung and avoids the mice receiving unknown quantities of the drug by licking their fur or exposing their air passages. Mice were weighed and carefully examined after anesthesia. Their fur remained glossy, physical activity remained unchanged and there was no loss of weight during the five weeks of treatment, indicating that their overall health and well being were not adversely affected by the procedures. The histological examination of the lung, liver and spleen tissues (Figure 5) demonstrated that introduction of the surfactants did not result in any inflammation or damage to the bronchial and alveolar structures. Moreover, there was no change in leukocyte (Figure 6A) and protein (Figure 6B) content in BAL and the overall cytological composition of lavage fluids were the same. The liver and spleen also did not exhibit any toxic response (Figure 5). We conclude that the proposed use of OG and DDM to treat MTB localized to the lung does not cause any destructive or inflammatory changes in the bronchial tree and the lung parenchyma and did not cause any registered toxic effects. The absence of any indication of damage to the parenchyma of the lung is probably related to the fact that human lung is bathed in surfactant (DPPC) from birth.
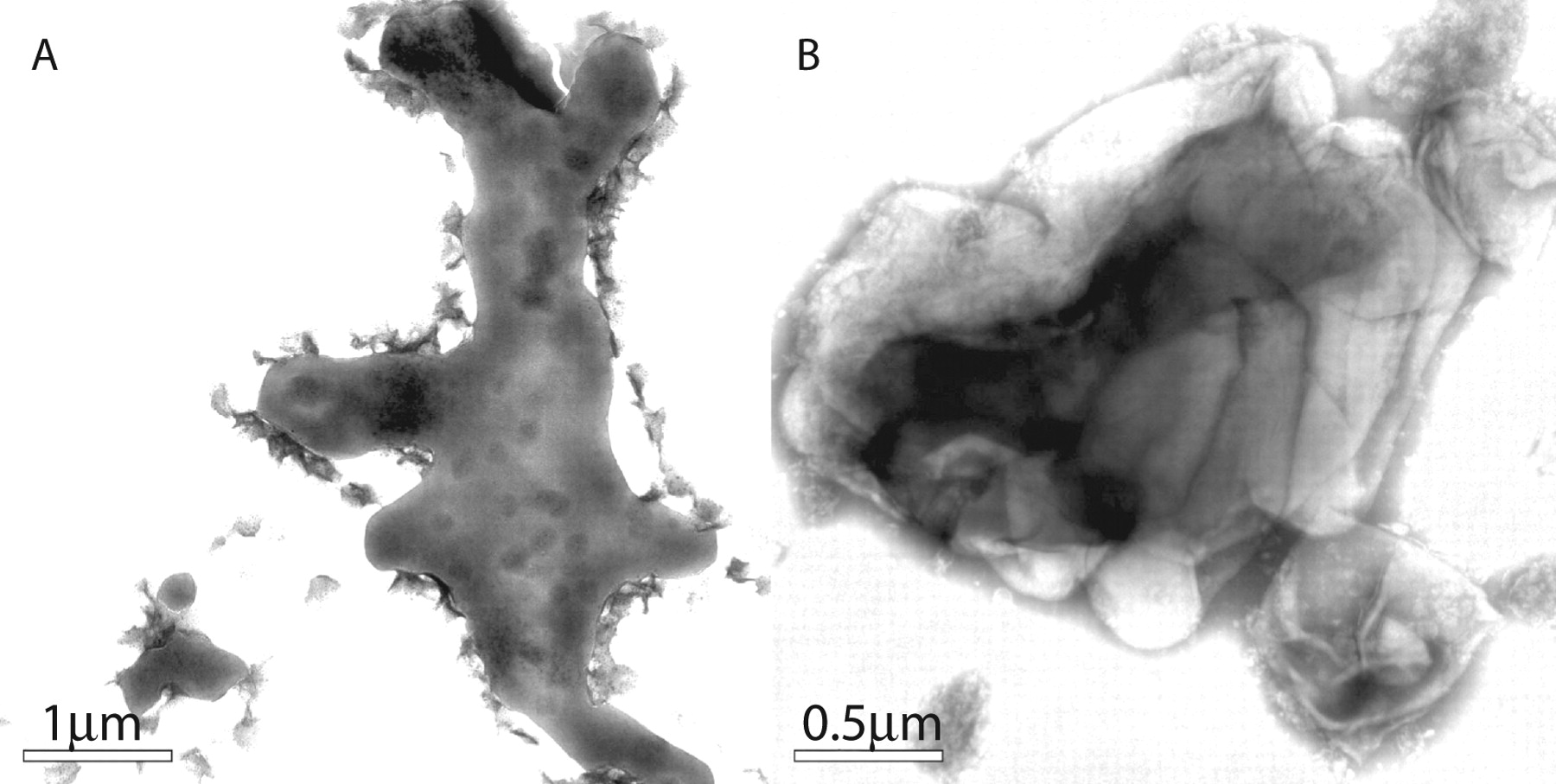
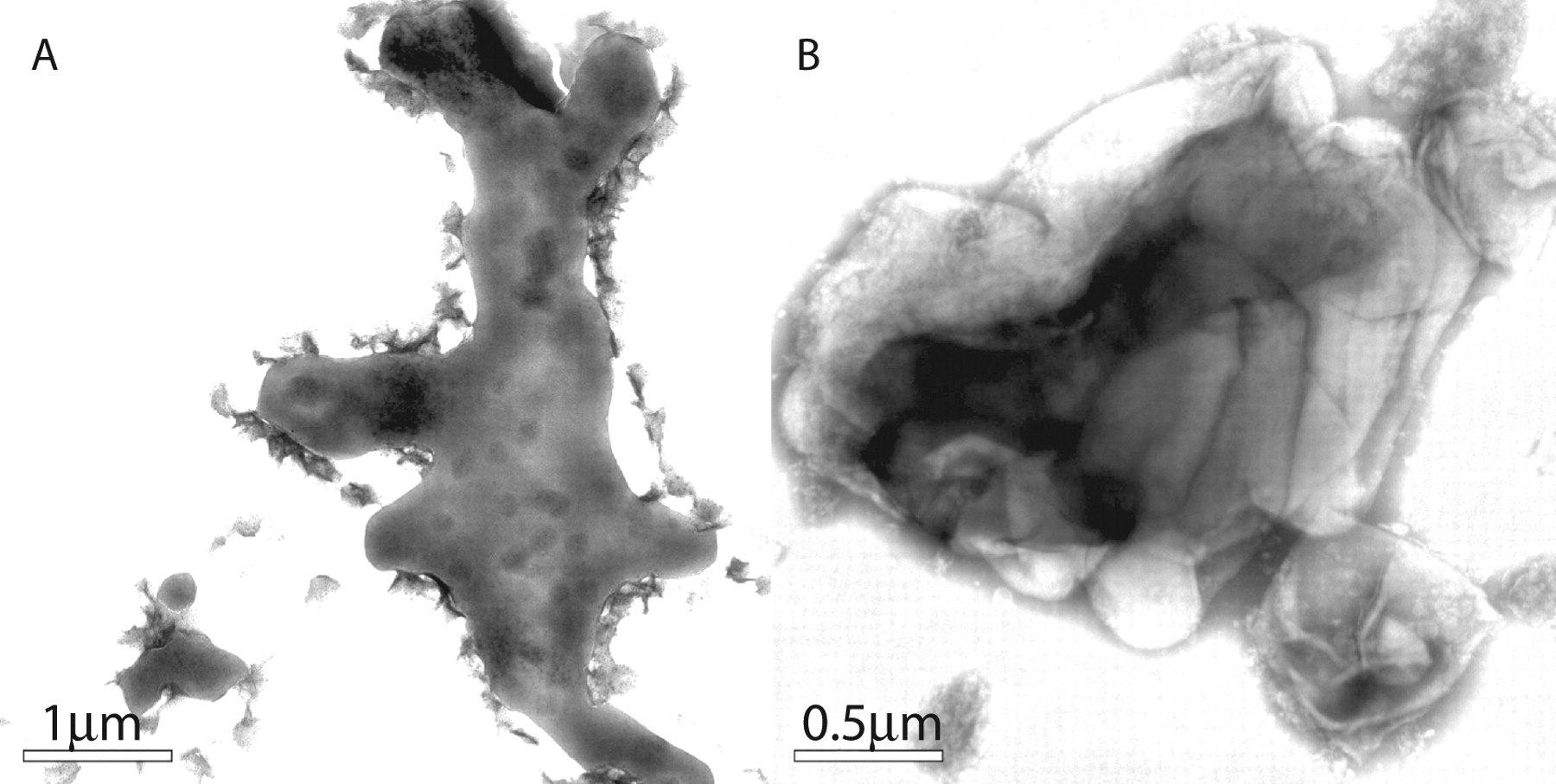
A: Appearance of bacteria grown in media in the absence of Tween. Instead of the net-like appearance, they are more condensed in amorphous structures of enormous size which probably represent the EM view of cords. B: The KasB strain also forms similar structures and sonication ruptures them to reveal their contents of MTB that reside inside. The image of this strain is shown because fortuitously a portion of the shattered sack broke off like glass and remained next to the aggregated bacteria. Its rigidity maintains the contour of the bacteria similarly to Figure 1B. It is reasonable to conclude that these sacks consist primarily of TDM and the underlying bacteria are also covered by TDM.
A recent study of the aerosolization of 100 μl of DDM at 0.24% into rat lungs showed an acute inflammatory response when administered seven days in succession (13). Toxicity was gauged by an aproximately two-fold increase in three enzyme activities and protein in the BAL over the control. Even though a full assessment of toxicity is not possible from this study, it may be advantageous to include a period between drug administrations to avoid any significant chronic toxic response as in our study.
Discussion
It is well known that 90% of patients who are infected with MTB self resolve the disease without clinical intervention (14). This notion seems improbable given the difficulty encountered in treating patients that have clinical symptoms. In any event, this self cure rate is impressive and suggests that an understanding of this mechanism may lead to a more effective treatment of the disease. We propose that lung surfactants may not only alter the TDM outer barrier resulting in bacteria that are more accessible to the immune system but also may adversely affect the function of their more extended membrane pore structures that are required for oxygen/carbon dioxide exchange and the entry of nutrients.
As a consequence of our EM and light microscopic studies on the effect of surfactants on the TDM outer coat, we propose that the lung surfactants DPPC and phosphatidyl glycerol (PG) limit the formation of the TDM protective coat similar to Tween 80 (cf. Figures 1 and 3). DPPC and PG constitute over 95% of lung surfactant (15) and we have shown that DPPC slowly reverses the corded state of the bacteria. It is apparent that the bacteria in the non-corded state (Figures 1 and 2) are more accessible to the immune system than those in the highly aggregated, corded state that are covered with TDM (Figures 3 and 4). Accordingly, it seems probable that lung surfactants may initially maintain bacteria in the non-corded state. Clearly, this scenario requires joint participation of two separate systems and if either is compromised the self cure fails such as in immune compromised individuals. The significance of the lungs surfactants in the progression of the disease has been established (16) and these studies support our hypothesis (see below).

Light microscopic characterization of the acid fast stained MTB in the corded state and the effect of 1% OG or DDM on the preparation grown without Tween. Corded bacteria (A-B). After exposure of bacteria to 1% OG for 30 min (C-D) and 24 h (E-F). Exposure of bacteria to DDM for 2 hrs, (G-H). The surfactants greatly reduce the size of the cords in a time-dependent manner and are bactericidal.
Our studies show that OG and DDM offer an effective, fast-acting method for reversing the corded state of MTB and are bactericidal (Figure 4). Most importantly, the effective concentration (1%) for this purpose is not toxic to mouse lung (Figures 5 and 6). A priori this result is unexpected because surfactants are non-specific and therefore are expected to express similar toxicity to all cells (12). We attribute this selectivity to the lungs parenchyma's adaptation to surfactants which begins before birth. Tissue suitability for harsh environments is well illustrated by the gastric mucosa in an acid environment. In contrast, the adjacent esophagus is non-tolerant and is susceptible to erosion. The insensitivity of lung tissue to surfactants raises the possibility of new therapeutic approaches utilizing these agents.
In addition to the MTB bactericidal activity of OG and DDM, these surfactants have been established as highly proficient cellular and tissue permeabilizing agents (12). For example, insulin aerosolized with tetradecyl-maltoside into the lungs of rats reaches the systemic circulation and regulates blood glucose levels (17). DDM with certain proteins have been infused i.v. into patients for treatment of multiple sclerosis (18). Consequently, the combination of antibiotics and surfactants aerosolized into the lungs offers an attractive alternative to the present systemic approach to treating pulmonary MTB with its associated toxic effects.
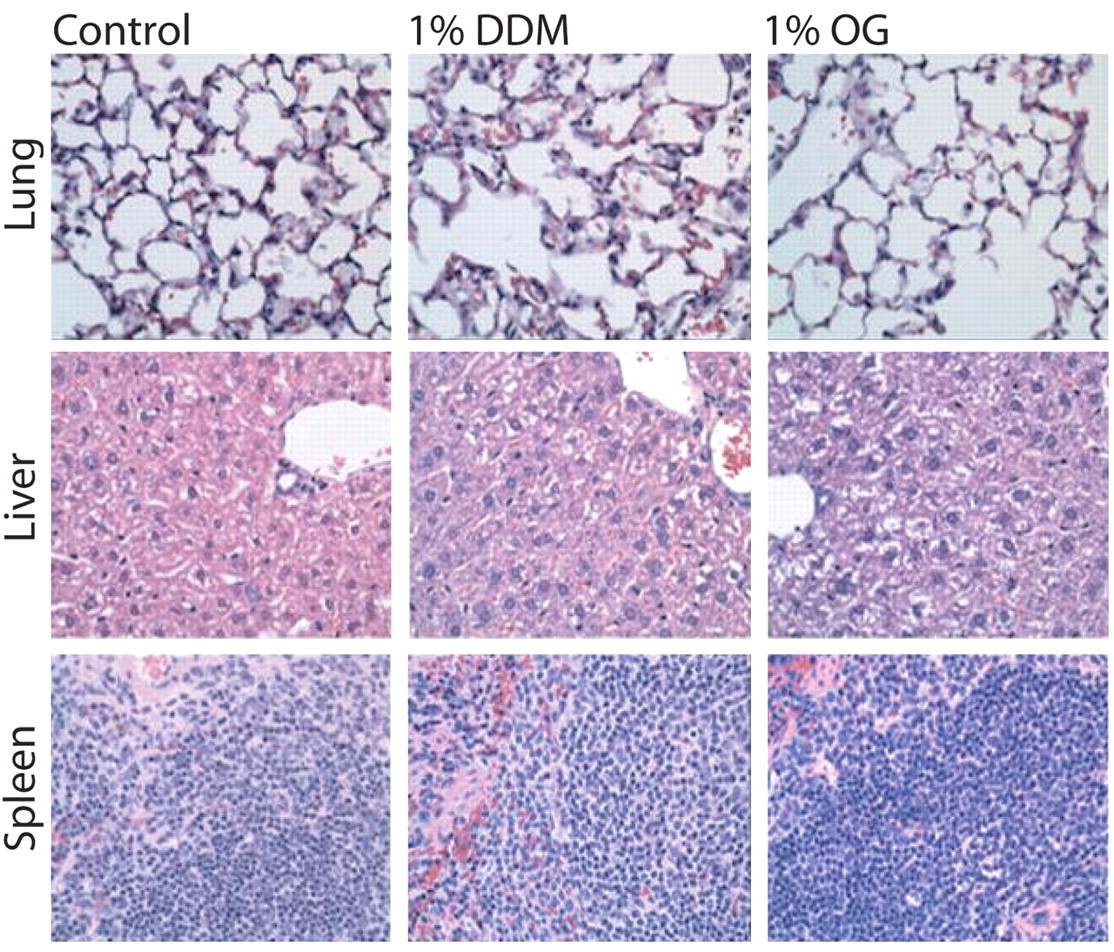
Histopathological evaluation of the toxicity resulting from the aerosolization of mouse lungs with 1% OG or DDM. Tissue specimens from the 0.1 and 0.2% groups were also evaluated (data not shown). Lungs, livers, and spleens exhibited no toxic response to OG or DDM and there was no discernable damage to the lung structures.

Cell counts (A) and protein concentration (B) in BAL from mouse lungs aerosolized with variable concentrations of OG (G) and DDM (M). The BAL differential showed 100% macrophages in control and 97% in 1% OG and DDM groups which also exhibited 3% lymphocytes but no eosinophils or neutrophils. The values are similar to the saline control and indicate no toxic response.

Light microscopic image of acid fast stained cavity lung specimen from a patient with cavitary MTB. The bacteria have a similar appearance to those in the corded state grown in culture (cf Figure 4B).
The effective cellular permeabilizing properties of surfactants (12) nicely addresses the issue of the bacteria “hiding out” in the protective environment of macrophages and dendrites (19) because the surfactants will augment the entrance of antibiotics into these cells and into the bacteria that reside inside. Instead, the TDM armor represents a more formidable barrier that OG and DDM proficiently remove (Figure 4).
In this regard, it was shown that a poloxamer surfactant enhanced the bactericidal activity of antibiotics in mice infected with TB and enhanced the activity in a streptomycin-resistant strain of TB in murine infection (20). These results are thought to have their foundation from similar results obtained from TB-infected macrophages in which the surfactant/antibiotic were found effective. These studies clearly show that the surfactant alter the bacterial coat to enhance their permeability to antibiotics (21).
Human and bovine surfactants have been aerosolized into the lungs of TB patients and when combined with the conventional application of antibiotics the procedure significantly reduces therapy to two months (16). It was proposed that the added surfactants help restore their normal composition in the lungs. These findings are encouraging and concur with our studies. However, OG and DDM are expected to have much greater efficacy because of their higher CMC values; lung surfactants 10−10 M cf. OG and DDM ~10−2 M (22). Moreover, it is possible to exploit the synergism of the surfactant combined with antibiotics administered directly to the lungs.
Ex vivo studies utilizing a surgically exercised lobe containing cavitary lesions are in progress. This approach addresses the proficiency issue in an extraordinary timely manner. The sack-like cavity consisting of fibrotic tissue is devoid of type two pneumocytes that produce all the lung surfactants. Therefore, presumably the cavity has little or no DPPC and PG which might otherwise hinder the formation of cords that cover its wall (Figure 7). Surfactants are introduced into the cavity on a surgically removed specimen through the bronchial opening utilizing a catheter. The efficiency of DDM or OG in removing the cords from the cavity and killing those bacteria that remain can be evaluated by light microscopy the day of the procedure and the number of colony-forming units can be determined utilizing tissue specimens taken prior to and after treatment. A comparative study in which the front line antibiotics are included with the surfactants will also be evaluated (1). This treatment regimen permits a gauge of efficacy on the day of the procedure, which expedites the development of new treatment options. Promising results from these studies may lead to an attractive alternative to the surgical treatment.
In the broader picture, the removal of most of the bacteria that are not firmly embedded in the cavity wall may abrogate or substantially decrease the release of viable virulent MTB and prevent the spread in susceptible populations. In clinical trials, the absence of MTB in sputum after therapy will permit a rapid evaluation of the treatment. The use of surfactants combined with chemotherapy may have a major impact in the treatment of some of the most dreaded pulmonary diseases such as lung cancer, bacterial pneumonia, and MTB. The perturbation and destruction of the cellular structure of pathogens by surfactants affords a novel approach to treating the rapidly developing and highly resistant forms MDR-TB, XDR-TB (23) and other mutants that are not expected to have or be capable of developing a defense. These studies may achieve a milestone in which surfactants combined with other drugs greatly improve the treatment of human diseases and propel surfactants to the forefront of clinical medicine.
Acknowledgements
This study was supported by the Gillson Logenbaugh Foundation and Vanjan Inc., Houston, TX.
We thank Dr. Robert Brown for support and helpful discussions, Dr. William R. Jacobs, Jr. for providing the kasB mutant, Bheravi Patel for preparation of the manuscript, Dr. Jun Liu for assistance in preparing the figures, and Dr. Robert L. Hunter for helpful discussions and practical introduction to pulmonary aerosolization.
- Received March 12, 2010.
- Revision received July 16, 2010.
- Accepted July 16, 2010.
- Copyright © 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}