


Abstract
Metal ions are released from casting alloys and cause damage to cell structures and local inflammation. However, the cytotoxic mechanism and the type of cell death induced in human gingival fibroblast (HGF) by contact with dental metals have not been well characterized. Here the cytotoxicity of eight metals against HGF was investigated. Cytoxicity of metals against HGF was in the following order: Ag(NH3)2F (most cytotoxic)>AgCl>CuCl2>CuCl, CoCl2> NiCl2>FeCl2, FeCl3 (least cytotoxic). None of the metals showed any apparent hormetic growth stimulation at lower concentrations, except for Ag(NH3)2F at 20 or higher population-doubling level of HGF. The sensitivity of HGF against Ag(NH3)2F was reduced during in vitro aging, similar to previous report with sodium fluoride. Contact with Ag(NH3)2F for only one hour induced irreversible cell death, whereas longer duration of contact with AgCl or CuCl2 was necessary to induce irreversible cell death. These metals induced neither DNA fragmentation nor caspase-3 activation. Pan-caspase inhibitor (Z-VAD-FMK) and autophagy inhibitors (3-methyladenine, bafilomycin) did not apparently affect the cytotoxicity of metals, when corrected for the effect of inhibitor alone on growth. We also found that Ag(NH3)2F induced much higher cytotoxicity than AgCl in mouse osteoblastic cell line MC3T3-E1, possibly inducing necrosis. These data suggest the importance of cautious application of Ag(NH3)2F to the oral cavity.
Dental casting alloys have been used throughout the history of dentistry, but studies have shown that the metal ions released from the casting alloys may damage cell structure and induce local inflammation (1-4). Gold (Au), silver (Ag) and palladium (Pd) have caused allergic reaction in the oral cavity, albeit infrequently (5). Ions released from nickel-based dental casting alloy have been reported to interfere with cellular energy metabolism and function of human gingival fibroblast (HGF) (6, 7). Recently, we have established an assay system for the interaction between metal and cultured cells (8-12). We found that contact with Cu plate induced rapid cell death and cysteine oxidation in HGF, and that the inclusion of Au at more than 10% in the Cu alloy completely eliminated both the cytotoxicity and oxidation (Yamazaki, unpublished data). At present, it is not yet clear whether the cytotoxicity induced by Cu plate is caused by Cu ion release by direct contact with the surface of the Cu plate. In the present study, we dissolved a total of eight metals in culture medium and added them to HGF cells to compare their relative cytotoxicity. It has been reported that metals, radiation and chemotherapeutic agents induce growth stimulation at lower doses/concentrations (so-called ‘hormesis’) (13). Therefore, we also investigated whether dental metals induce such a hormetic effect in HGF cells.
Materials and Methods
Materials. The following chemicals and reagents were obtained from the indicated companies: alpha minimum essential medium (α-MEM), Dulbecco's modified Eagle's medium (DMEM) (GIBCO BRL, Grand Island, NY, USA); fetal bovine serum (FBS) (JRH Bioscience, Lenexa, KS, USA); CuCl, CuCl2·2H2O, AgCl, FeCl2·5H2O, FeCl3·6H2O, NiCl2·6H2O, CoCl2·6H2O, bafilomycin (Wako Pure Chem Co., Tokyo, Japan); Ag(NH3)2F (Bee Brand Medico Dental Co., Osaka, Japan); MTT [3-(4, 5-dimethyl-thiazol-2-yl)-2,5-diphenyltetrazolium bromide], 3-methyladenine (Sigma-Aldrich, St. Louis, MO, USA); pan-caspase inhibitor (Z-VAD-FMK) (Biomol, Enzo Life Science, Plymouth Meeting, PA, USA). Culture plastic dishes and plates (6-well, 96-well) were purchased from Becton Dickinson, Franklin Lakes, NJ, USA.
Cell culture. HGF cells were established from the first premolar extracted tooth in the lower jaw and periodontal tissues of a twelve-year-old girl, according to the guideline of the Intramural Board of Ethics Committee after obtaining informed consent from the patient. These cells were cut into small pieces by surgical blade, and placed onto a 80-mm plastic dish to allow outgrowth during 2 weeks' culture in α-MEM supplemented with 20% heat-inactivated FBS, 100 units/ml, penicillin G and 100 μg/ml streptomycin sulfate at 37°C under a humidified 5% CO2 atmosphere. The cells were then used as primary culture with population-doubling level (PDL) zero. Cells were then harvested by treatment with 0.25% trypsin-0.025% EDTA-2Na in phosphate-buffered saline without calcium and magnesium [PBS(−)] (pH 7.4).
The number of the viable cells that excluded trypan blue was determined by hemocytometer under light microscopy and the total number of viable cells calculated. Subculture of HGF cells was carried out every week with 1:4 split ratio and the medium changed in between the subcultures. HGF had an in vitro lifespan (cumulative cell population doubling number) of 47, regardless of the culture medium (either DMEM or α-MEM). The saturation density of the cells declined with in vitro senescence in both media, but it was higher in DMEM/10%FBS than in α-MEM/10% FBS (14).
MC3T3-E1 mouse cells derived from the calvaria of C57BL/6 mice were sub-cultured as adherent cells in α-MEM supplemented with 10% heat-inactivated FBS, as described previously (12). Cells were washed with [PBS(−)] and detached by 0.25% trypsin-0.025% EDTA-2Na in PBS(−) for each experiment.
Determination of viable cell number. Cells were inoculated at 4×103/ml in 96-microwell plates and incubated for 48 hours to allow complete attachment. The cells were then incubated for 24 hours with fresh culture medium containing different concentrations of each metal. The viable cell number was then determined by MTT method. In brief, cells were incubated for 4 hours with 0.2 mg/ml MTT in fresh DMEM with 10%FBS. The formazan formed during incubation was dissolved with 0.1 ml of dimethyl sulfoxide, and the absorbance at 540 nm of the lysate was determined by using a microplate reader (Multiskan, Biochromatic, Labsystem, Osaka, Japan).
Assay for DNA fragmentation. HGF cells (16×104) were seeded on 6-well plates and incubated for 48 hours to allow complete attachment. Cells were then incubated for a further 6 hours with the indicated concentrations of metals. After washing twice with PBS(−), cells were collected by scraping with a rubber policeman on ice and spun down in an eppendorf tube. Cells were lysed with 50 μl lysate buffer [50 mM Tris-HCl (pH 7.8), 10 mM EDTA, 0.5% (w/v) sodium N-lauroylsarcosinate]. The solution was incubated with 0.4 mg/ml RNase A and 0.8 mg/ml proteinase K for 2 hours at 50°C and then mixed with 50 μl NaI solution (40 mM Tris-HCl (pH 8.0), 7.6 M NaI, 20 mM EDTA-2Na) followed by 250 μl of ethanol. After centrifugation for 20 minutes at 20,000×g, the precipitate was washed with 1 ml of 70% ethanol and dissolved in TE buffer [10 mM Tris-HCl (pH 8.0), 1 mM EDTA-2Na]. Each sample (10-20 μl equivalent to 5×105 cells) was applied to 2% agarose gel electrophoresis in TBE buffer (89 mM Tris-HCl, 89 mM boric acid, 2 mM EDTA-2Na). After staining with ethidium bromide, the DNA was visualized by UV irradiation, and photographed as described previously (15). DNA from apoptotic HL-60 cells induced by UV irradiation (6 J/m2/min, 1 min) (16) was run in parallel as positive controls.

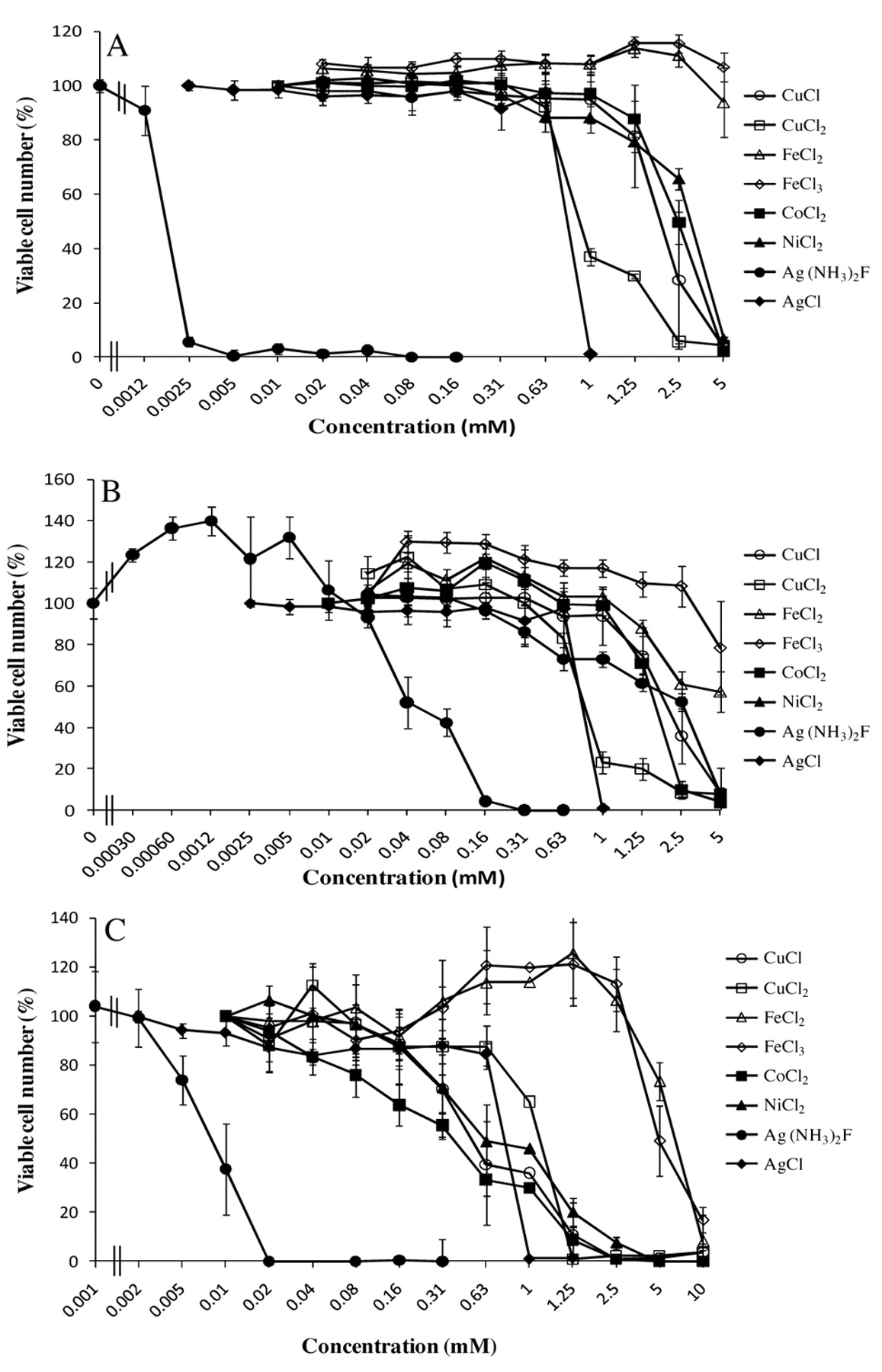
Cytotoxicity of eight metals against HGF and MC3T3-E1 cells. HGF cells at 14 PDL (A) or 20 PDL (B), and MC3T3-E1 (C) cells were incubated for 24 hours with the indicated concentrations of each metal and the relative viable cell number was then determined by MTT method. Each value represents the mean±S.D. of triplicate assays. Reproducible results were obtained in another two independent experiments.
Assay for caspase-3 activation. HGF cells (5×105) were seeded on 50-mm dishes, and incubated for 48 hours to allow complete adherence. Cells were then incubated for a further 6 hours in fresh medium containing the indicated concentrations of metals. Cells were washed twice with PBS(−) and lysed with 100 μl of lysis solution. Cells were collected by scraping with a rubber policeman and transferred to an eppendorf tube. After standing for 10 min on ice and centrifugation for 5 min at 10,000×g, the supernatant was collected. Lysate (50 μl, equivalent to 200 μg protein) was mixed with 50 μl lysis solution containing substrates for caspase-3 (DEVD-pNA (p-nitroanilide)). After incubation for 4 hours at 37°C, the absorbance at 405 nm of the liberated chromophore pNA was measured by microplate reader as described previously (15).
Effect of apoptosis and autophagy inhibitors. HGF cells (1×104) were inoculated on a 96-well plate, and incubated for 48 hours to allow complete adherence. Cells were then preincubated for 60 minutes with either 50 μM caspase inhibitor (Z-VAD-FMK), 10 mM 3-methyladenine (phosphatidylinositol 3 kinase inhibitor that inhibits the formation of autophagosome) (17) or 100 nM bafilomycin (V type proton pump inhibitor that inhibits the formation of secondary lysosome) (18), 60 minutes before the addition of each metal: 0.04 mM Ag(NH3)2F, 0.8 mM AgCl, 1 mM CuCl2. Cells were then incubated for a further 24 hours and then assayed for viable cell number by MTT method.

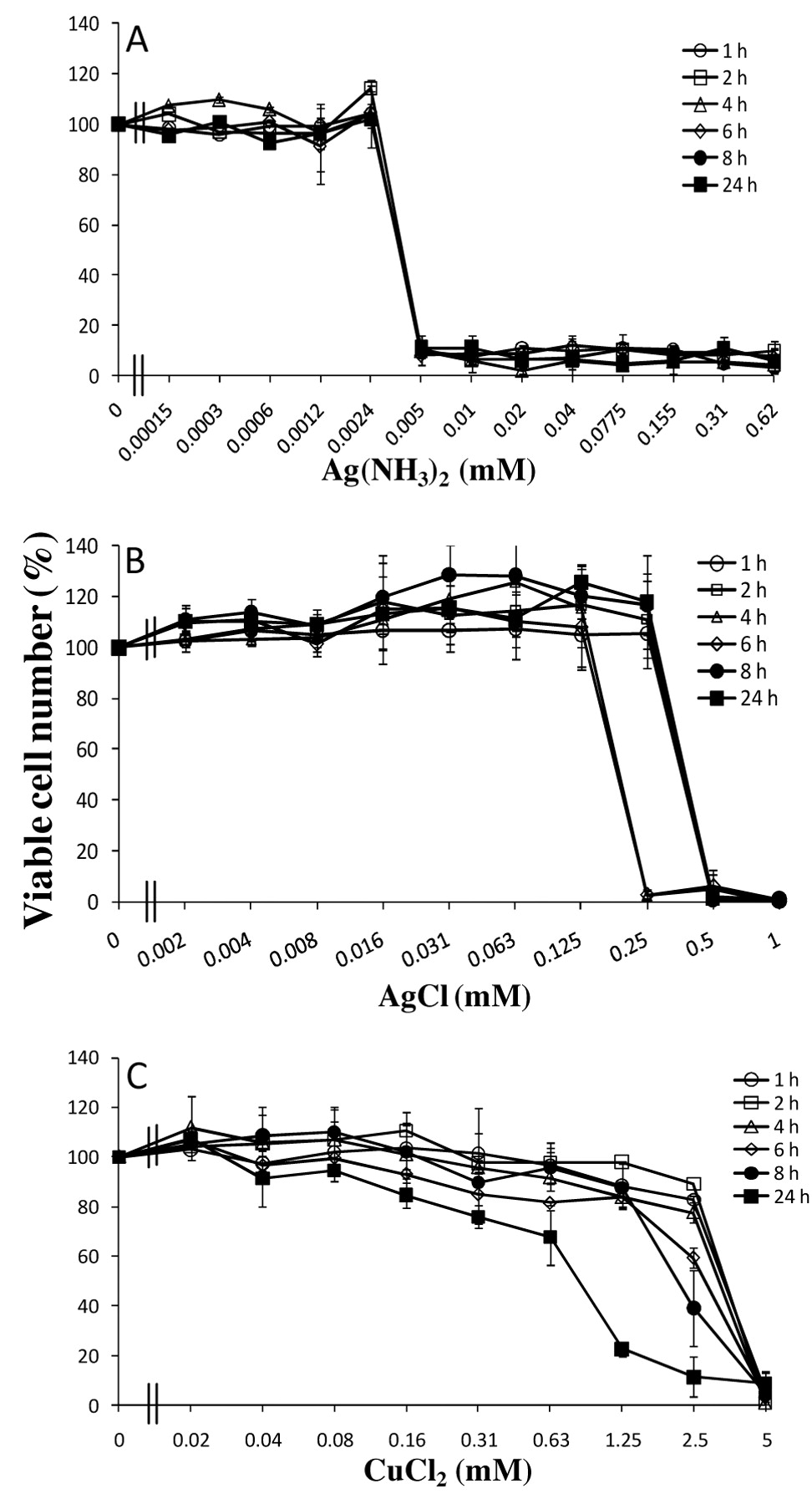
Determination of minimum exposure time necessary for cell death induction by metal. HGF cells (6 PDL) were incubated for 1, 2, 4, 6, 8 or 24 hours with different concentrations of Ag(NH3)2F (A), AgCl (B), CuCl2 (C) and then resuspended with fresh culture medium, and then further incubated for 23, 22, 20, 18, 16 or 0 hours before MTT assay (Total incubation time=24 hours). Each value represents the mean±S.D of three determinations.
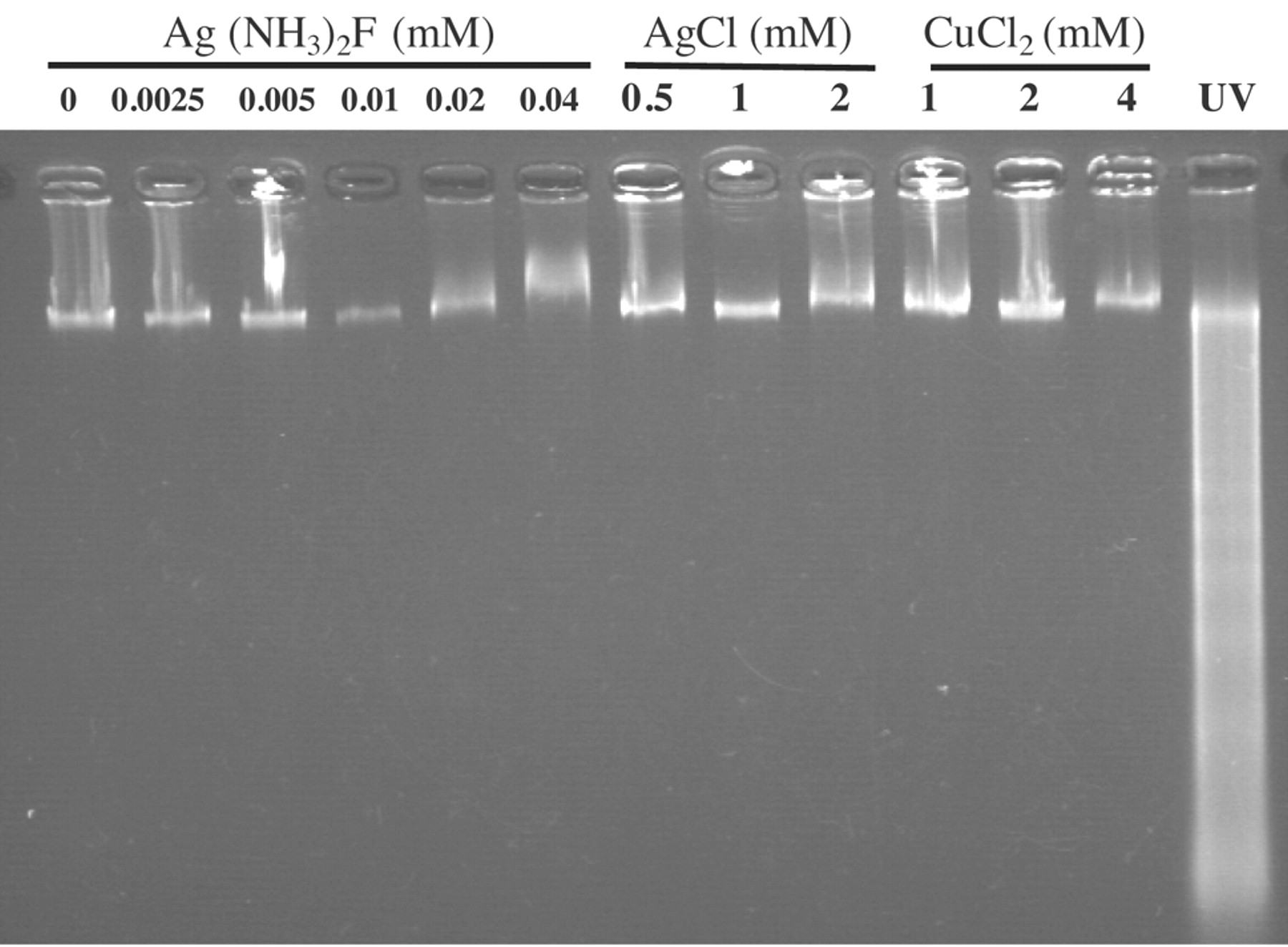
Metals failed to induce internucleosomal DNA fragmentation. HGF cells (8 PDL) were incubated for 6 hours with the indicated concentrations of Ag(NH3)2F, AgCl, CuCl2, and then assayed for DNA fragmentation by agarose gel electrophoresis.
Results
Cytotoxicity of metals. The cytotoxicity of metals was determined by incubating the exponentially growing HGF cells with increasing concentrations of each metal. Cytoxicity of metals against HGF less than 14 PDL was in the following order: Ag(NH3)2F (CC50=0.0018 mM) >AgCl (CC50=0.77 mM) >CuCl2 (CC50=1.11 mM) >CuCl (CC50=1.98 mM) >CoCl2 (CC50=2.5 mM), NiCl2 (CC50=3.18 mM) >FeCl3 and FeCl2 (no apparent cytotoxicity) (least toxic) (Figure 1A). HGF at higher PDL had an almost similar order of sensitivity to these metals: Ag(NH3)2F (CC50=0.048 mM) >AgCl (CC50=0.77 mM), >CuCl2 (CC50=0.97 mM) >CuCl (CC50=1.48 mM) >CoCl2 (CC50=1.73 mM), NiCl2 (CC50=2.59 mM) > FeCl3 and FeCl2 (no apparent cytotoxicity) (least toxic) (Figure 1B). Similarly, Ag(NH3)2F showed the highest cytotoxicity towards mouse osteoblastic cells MC3T3-E1 cells (Figure 1C).
None of the metals had any apparent growth-stimulating effect (so-called ‘hormesis’) at lower concentrations, except for Ag(NH3)2F in HGF cells at 20 or higher PDL. Time course study demonstrated that contact with Ag(NH3)2F for only one hour induced irreversible cell death, whereas longer contact with AgCl or CuCl2 was necessary to induce the irreversible cell death (Figure 2).
Type of cell death induced by metals. Ag(NH3)2F, AgCl and CuCl2 induced neither clear-cut DNA fragmentation (either smear or internucleosomal) (Figure 3) nor caspase-3 activation (Figure 4). Pretreatment with 3-methyladenine, and bafilomycin did not affect the cytotoxicity of Ag(NH3)2F, AgCl and CuCl2, when corrected for cytotoxicity or growth stimulation by inhibitor alone (Figure 5).

Metals failed to activate caspase-3. HGF cells (10 PDL) were incubated for 6 hours with the indicated concentrations of Ag (NH3)2F, AgCl, CuCl2, and then lysed for the assay of caspase-3 activity. Apoptotic HL-60 cells induced by UV were was used as positive control. Each point represents the mean±S.D.of triplicate assays.
Discussion
The present study confirmed the previous finding that both Ag+ was the most cytotoxic among various metal cations tested (19, 20) and failed to induce internucleosomal DNA fragmentation (19). This suggests that Ag+ induced non-apoptotic cell death, but the type of cell death has not yet been clarified. We found that neither Ag(NH3)2F nor AgCl induced apoptotic markers such as internucleosomal DNA fragmentation and caspase activation, and that apoptosis and autophagy inhibitors did not modify the Ag+-induced cytotoxicity. All these data suggest that the type of cell death induced by Ag+ may be by means other than apoptosis and autophagy, probably necrosis. Since silver-impregnated collagen cuffs used in tunneled catheters may induce local cytotoxicity (21), the cytotoxicity of Ag+ against normal oral cells remains to be investigated in more detail for their future use without consequences.
Ag(NH3)2F, known as an anticaries and antibacterial agent, was one order more cytotoxic against HGF (CC50=0.0018-0.048 mM) than were AgCl (Figure 1) (CC50=0.77 mM) and NaF (CC50=5 mM) (22). Higher cytotoxicity of Ag(NH3)2F than AgCl was also found in mouse osteoblastic cells MC3T3-E1 cells (Figure 1C). This further supports the previous finding that the chemical synthesis of silver diamine fluoride through the interaction between fluoride and silver synergistically enhances cytotoxicity compared to that of the individual metals (23). We found similarity between Ag(NH3)2F and AgCl, in respect to the lack of apoptosis-inducing activity and inhibitor sensitivity. It has been reported that Ag is released from alloy elements (24), suggesting that the released Ag+ may significantly affect the cellular function. However, further studies are required to identify the point of action of Ag compounds.

Effect of several inhibitors on metal-induced cytotoxicity. HGF cells (12 PDL) were pretreated for 1 hour without (control), or with pan-caspase inhibitor (Z-VAD-FMK; 0.05 mM), 3-methyl adenine (3-MA; 10 mM) or bafilomycin (BAF; 100 nM), and then Ag(NH3)2F (0.04 mM), AgCl (0.8 mM) or CuCl2 (1 mM) was added. After incubation for 24 hours, the viable cell number was determined by MTT method. Each value represents the mean±S.D of three determinations.
We also found that the sensitivity of Ag(NH3)2F declined during in vitro aging of cells, similar to that of NaF (22). This suggests some modification of the transport system of these cells for these metals during aging.
- Received February 27, 2010.
- Revision received May 26, 2010.
- Accepted June 4, 2010.
- Copyright © 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.