


Abstract
Background: The aim of this study was to investigate the effect of fetal exposure to 4-n-octylphenol (OP) on the induction of mammary tumors induced by 7,12-dimethylbenz[a]anthracene (DMBA) in rats. Materials and Methods: All rats which showed vaginal plugs or sperm by vaginal smear test after mating were fed a normal diet only, or a diet mixed with 10, 100, 1000 ppm OP throughout the pregnancy period and from day 13 of pregnancy to the end of the pregnancy. No abnormal pregnancy was seen in any of the rats. Pups were given 10 mg DMBA by gastric intubation 50 days after birth. Results: Uterine weight decrease was observed in pups with fetal exposure to 1000 ppm OP throughout the pregnancy period. No endocrine disrupting conditions, such as persistent estrus, anovulatory ovaries or abnormal lactation in the mammary glands were seen in pups with fetal exposure to OP. However, fetal exposure to 100 and 1000 ppm OP throughout pregnancy period enhanced the early incidence and number of mammary carcinomas (MCs) while it did not enhance the incidence and number of benign proliferative lesions (PLs) which consisted of solid masses (fibroadenoma and lobular hyperplasia) and gross cysts. Conclusion: These results suggest that fetal exposure to the very weak estrogenicity of OP could enhance the induction of MCs but not affect the induction of PLs.
In humans, the incidence of breast cancer has been increasing worldwide (1, 2), and it is essential for its prevention to accurately determine the cause. In general, the risk of developing breast cancer is mainly related to carcinogens, daily diet, lifestyle, environment and genetics. The relationship of breast cancer risk to environmental agents, such as chemical exposure, has been widely studied (3-5). Exposure to certain chemicals and hormone-mimicking or endocrine disruptors is suspected to contribute to the increasing incidence of breast cancer as well as precocious puberty in the United States (6).
We have reported that neonatal administration of diethylstilbestrol (DES) (7-9), 17β-estradiol (E2) (10) and testosterone propionate (11-14) affected the induction of mammary tumors (MTs) induced by 7,12-dimethylbenz [a]anthracene (DMBA) in female rats while neonatal single administration of 4-n-octylphenol (OP) in female rats showed no effects of persistent estrus, anovulatory ovaries or the induction of MTs (15). We have also reported that rat mammary dysplasia (MD) induced by DMBA is morphologically similar to that in humans, and is characterized by gross cysts and solid masses with microscopic features including fibrotic adenosis (FA) and acinar adenosis (AA) (9, 11, 13, 14, 16). MD, FA and AA are relevant to benign proliferative lesion (PL), fibroadenoma and lobular hyperplasia in Fisher rats, respectively (17). Recently, we also reported that DES affects pregnancy and fetal exposure to DES induces endocrine disrupting conditions and enhances the induction of mammary carcinomas (MCs) and benign proliferative lesions (PLs) in rats (18). However, it remains to be determined whether high dose of OP, to which humans are daily exposed and has weak estrogenicity (19-21), affects pregnancy, and whether fetal exposure to OP affects the induction of MTs in rats.
Octylphenols, compounds derived from nonylphenol, are estrogenic endocrine disrupting agents. They have been detected in the soils of rivers and may have hazardous effects on certain fish and shellfish species, being passed along the biological food chain. They stimulate the growth of estrogen receptor-positive human mammary carcinoma cells, and also increase the uterine weight and height of the endometrial epithelium in ovariectomized rats (22). The estrogenicity of OP was very weak and thought to be approximately 1/1000 that of DES. In this study, the dose of OP was from 10 to 1000 ppm as 100 ppm OP reduced DMBA-induced mammary tumor multiplicity in female c-Ha-ras proto-oncogene transgenic rats (22).
Huggins et al. previously showed that a single administration of 20 mg DMBA 50 days after birth induced visible mammary cancer in rats by 110 days after birth (23). This animal has since been a useful model for studying the carcinogenesis of human breast cancer. DMBA is known to be a polycyclic hydrocarbon carcinogen forming DNA adducts (24). The presence of estrogen and progesterone receptors in DMBA-induced MCs has been well documented in rats (25, 26). DMBA-transformed cells (intraductal papillary proliferative lesions) were able to exist in unpalpable lesions long after the administration of DMBA under conditions in which estrogen was present but progesterone was absent (12).
The mammary line stage is observed in 6-14 mm crown-rump length embryos between day 11 and 13 of pregnancy, and then hillocks following the mammary line stage are observed on day 13 of intrauterine life in 17-18 mm crown-rump length fetuses (27). Mammary hillocks persist through the 16th day, and then become less evident, although they apparently occupy the position of the future nipples (27).
Therefore, in this study, pregnant rats were fed on a diet mixed with OP throughout their pregnancy period or from day 13 of pregnancy until the end of the pregnancy. The aim of this study was to investigate how OP exposure to pregnant rats before or after the development of mammary hillocks affects the induction of MTs induced by DMBA in female pups.
Materials and Methods
Animals. The animals were inbred Sprague-Dawley female adult rats, our own breeding animals. Their age and body weights (BWs) were 7-11 months of age (mean value 8.0±1.5) and 214.4-265.4 g (mean value 238.5±12.1), respectively. The animals were maintained in filtered air laminar flow at the Institute of Laboratory Animal Sciences, Frontier Science Research Center, Kagoshima University, and given a commercial diet (CE-2; CLEA Inc., Tokyo, Japan) and tap water ad libitum. CE-2 contains 4-5 ppb estradiol/kg, 25.1% crude protein and 4.8% crude fat. Crude protein is derived from soybean meal, white fish meal and yeast, and crude fat is derived from germ oil and soybean oil. Soybeans contain 0.072-0.249 isoflavones/100g of dry weight (28). The room temperature was maintained at 25°C±2°C and relative humidity at 55%±10%, with a 12 h light/dark cycle. The use of animals in this research complied with all relevant guidelines set by Kagoshima University.
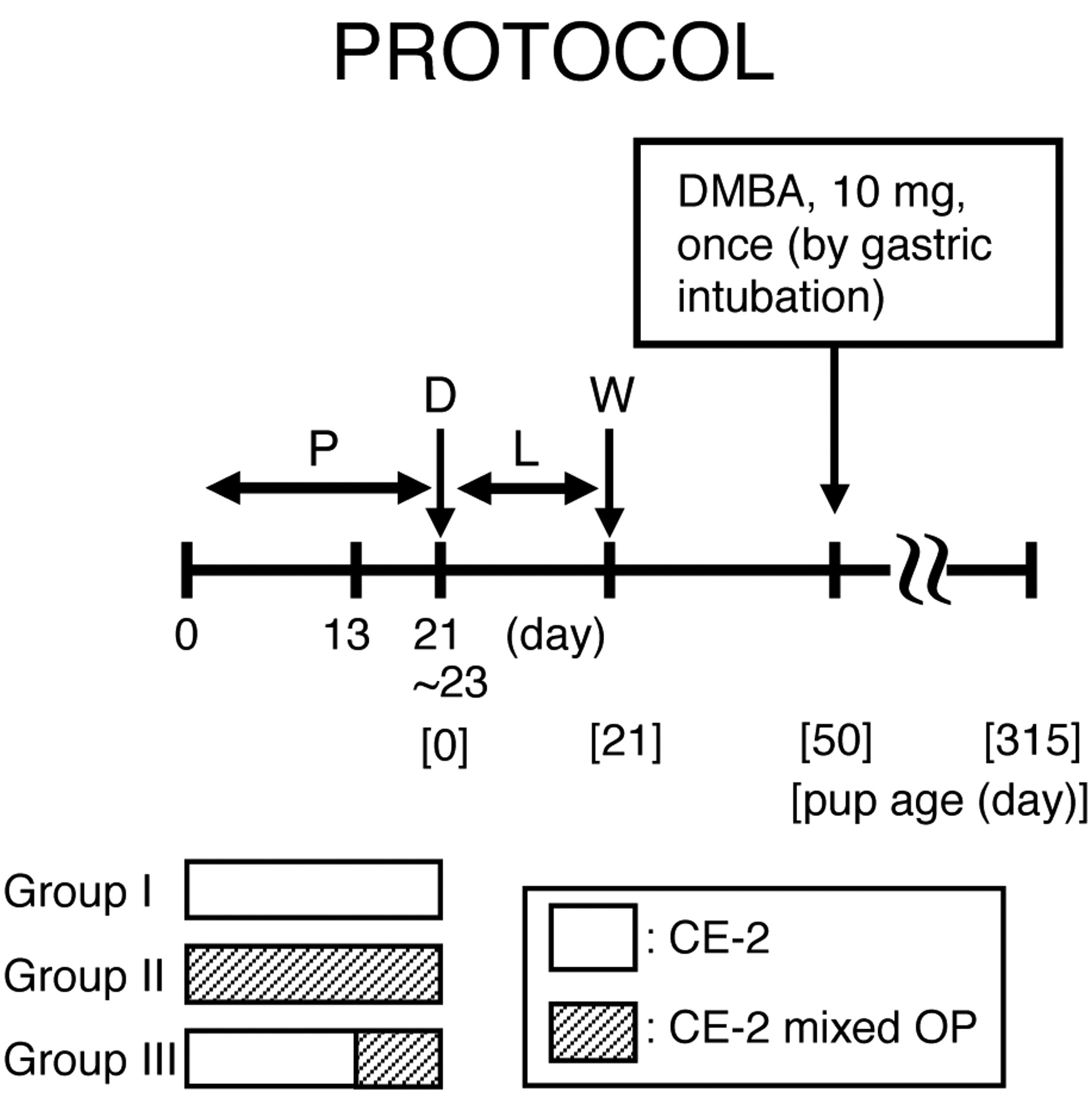
Experimental design. All rats in which vaginal plugs were observed or sperm by vaginal smear test after mating were divided into three groups (Figure 1). Rats in groups I (n=3), II (n=13) and III (n=10) were fed only CE-2, CE-2 mixed OP (Aldrich Chemical Company, Inc., Milwaukee, Wisconsin, USA) throughout the pregnancy period, and CE-2 mixed OP from day 13 of pregnancy until the end of the pregnancy. OP in subgroup a, b and c was mixed into the diet at 10, 100 and 1000 ppm, respectively. The number of animals in groups IIa-c and IIIa-c was 4, 5, 4, 4, 3 and 3, respectively. Animals in groups IIIa-c were fed only CE-2 during days 0-13 of pregnancy, comparable to the control group. All pups were weighed at birth. All lactating dams were fed only CE-2. All pups were weaned 21 days after birth and all dams underwent necropsy after weaning. At 50 days after birth, all female pups were given 10 mg DMBA (Wako Pure Chemical Industries Ltd., Osaka, Japan) dissolved in 1 ml sesame oil by gastric intubation. All DMBA-administered animals, except those intermediately sacrificed due to moribund state by tumor growth, were examined once weekly by palpation to detect mammary tumors from 50-315 days after birth. All palpable mammary masses were weighed, and the longest diameter and shortest diameter of the surface were measured. The size (cm2) was calculated by multiplying these lengths. The estrus cycles of all animals were examined once daily from 50 to 70 days after birth by a vaginal smear test. The body, ovaries, uterus, adrenal glands and pituitary gland of all animals were weighed.
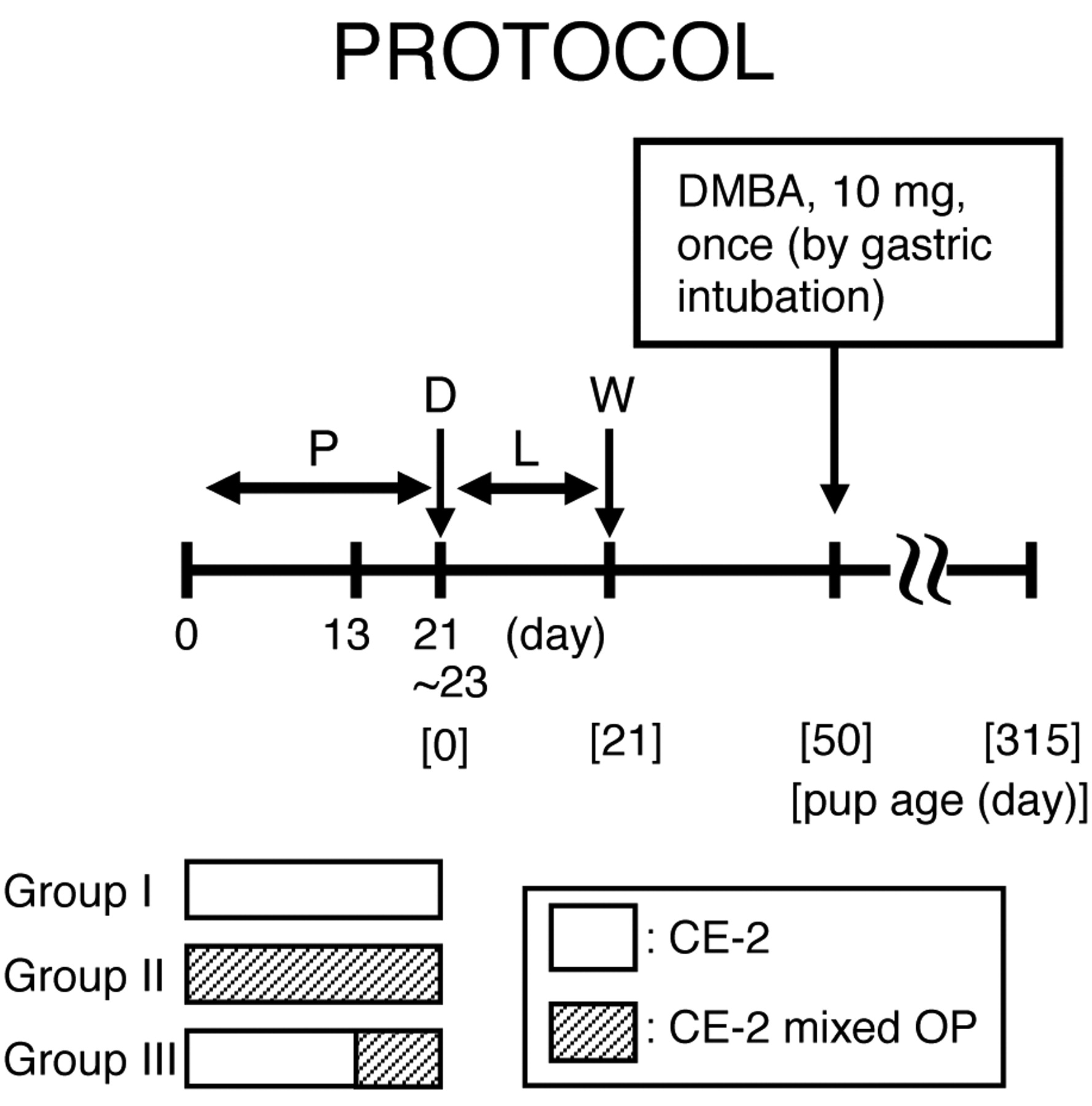
Summarized protocol. P: pregnancy, D: delivery, L: lactation, W: weaning.
Histopathological examination. Right abdominal mammary glands (R4-6), ovaries, adrenal glands, pituitary gland, uterus and vagina in dams were fixed in 10% phosphate-buffered formalin, dehydrated, and embedded in paraffin. All mammary tumors, residual mammary glands (L1-6 and R1-6), ovaries, adrenal glands, pituitary gland, uterus and vagina in female pups were fixed in 10% phosphate-buffered formalin, dehydrated, and embedded in paraffin. They were sectioned at 5 μm, stained routinely with hematoxylin and eosin (HE) and then examined histopathologically.
Statistics. Statistical analyses were performed using SPSS II (SPSS Japan Inc., an IBM company, Tokyo, Japan) for Windows, as described previously (8, 15). Mean differences were evaluated by Student's t-test. Data are shown as the mean±standard deviation (SD). The incidences (percentages) were tested using a four-fold contingency table (Chi-square test).
The Effects of 4-n-octylphenol (OP) on food volume in pregnancy.
Results
There were no significant changes in the mean daily food intake during pregnancy (Table I). Delivery occurred in all bred rats, while one mother rat in group IIIb and one mother rat in group IIIc ate all pups within 24 hours after delivery. The pregnant periods of all delivered rats were 21-23 days (mean value 22.6±0.6). No statistically significant changes were seen in the BW gain of bred rats during pregnancy (Figure 2). A dam with 5 male and female pups each in group IIIa died accidentally during the lactation period. Four female pups, 4, 3, 6 and 1 in groups I, IIb, IIc, IIIa and IIIb, respectively, died during the lactation period. No animals with persistent estrus (PE) were seen from 50 to 70 days after birth, and the patterns were regular with successive estrus cycles of 4 or 5-day duration. There were no significant changes in the number and BW of both male and female pups at birth (Table II).
The absolute weights (AW) and AW/BW of the uterine of pups in group IIc at necropsy were significantly lower than in group I (control) (Table III).
Pathology of mammary tumors (MTs) induced by DMBA. Single or multiple mammary masses, diagnosed as MC (adenocarcinoma) or PL, were seen in rats; no metastasis was recognized in any rat. These masses were localized in all areas (L1-6 and R1-6) of mammary glands. MCs were briefly subclassified into the papillary type, solid type or secretory type in this study, as described previously (7, 29, 30). PLs consisted of solid masses, which included fibroadenoma and lobular hyperplasia, and GCs, as described previously (7, 15, 16).
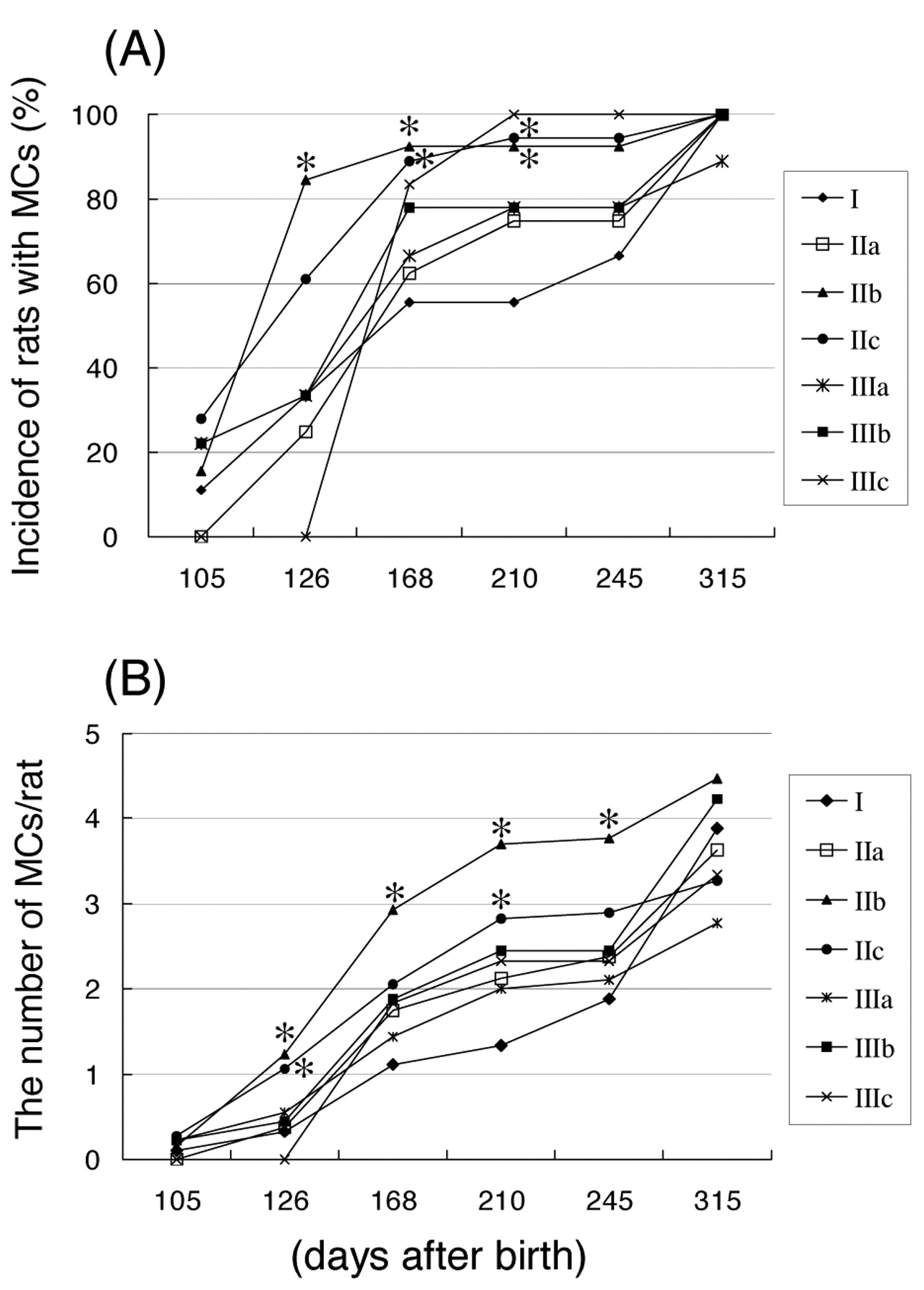
Figure 3 shows that the incidence of MCs in group IIb at 126, 168 and 210 days after birth, and group IIc at 168 and 210 days after birth was significantly higher than in controls. The number of MCs/rat in groups IIb at 126, 168, 210 and 245 days after birth, and group IIc at 126 and 210 days after birth was significantly higher than in controls. Table IV shows that the mean day of detection (latent period) in groups IIb and IIc was significantly shorter than in controls.
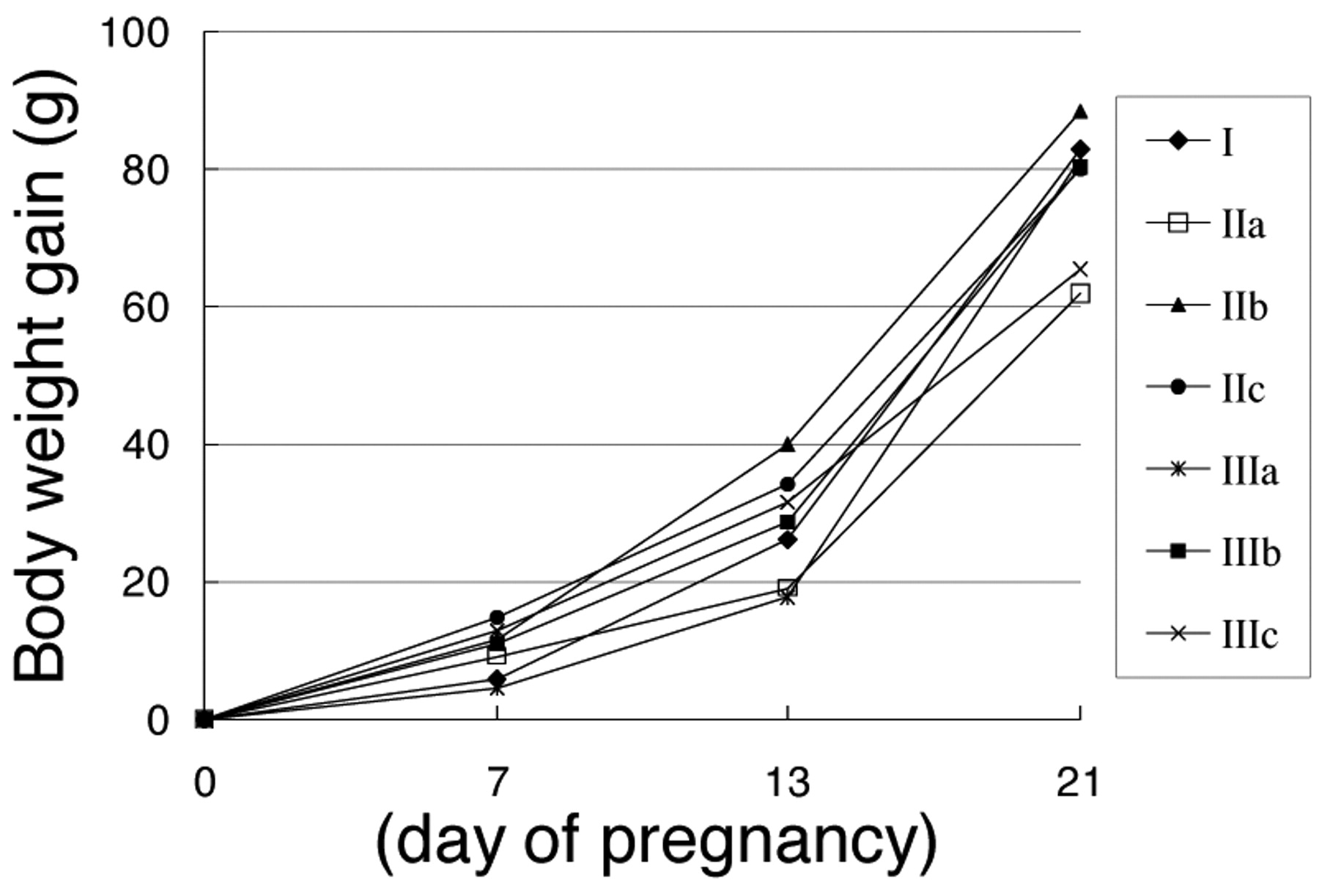
Body weight gain (g) on day 7, 13 and 21 of pregnancy.
In benign PLs, Figure 4A-4B, Tables V and VI show no evidence of promotion or inhibition. Table VII shows the observed benign and malignant mixed MTs among masses which diagnosed as adenocarcinoma. The latent period of mixed MTs was longer than that of MCs. These mixed MTs showed various histological features, such as a mass composed of adenocarcinoma, hyperplasia and fibroadenoma (Figure 5).
Histopathological changes in mammary glands and other endocrine-related organs. Physical active lactation was seen in the mammary glands of all dams. No lactation was seen in the mammary glands of any female pups. Corpus lutea (CL) were seen in the ovary of all dams and female pups. No abnormal changes, such as prolactinoma, were seen in the pituitary gland. Similarly, no abnormal changes such as atrophy and neoplasm were observed in the uterus and vagina, nor in the adrenal glands of all dams and female pups.
The effects of 4-n-octylphenol (OP) in the number and body weight (BW) of pups at birth.
The effects of 4-n-octylphenol (OP) on the body and organ weights in female pups at necropsy.
Effects of 4-n-octylphenol (OP) on induction of mammary carcinomas (MCs) in female pups.
Effects of 4-n-octylphenol (OP) on induction of benign proliferative lesions (PLs) in female pups.
Effects of 4-n-octylphenol (OP) on the number (incidence) of solid masses and gross cysts in benign proliferative lesions (PLs) in female pups.
Effects of 4-n-octylphenol (OP) on induction of benign and malignant mixed mammary tumors (MTs) in female pups.
Discussion
Exposure of pregnant rats to 1-100 ppm DES previously affected pregnancy and resulted in sterility, abortion, and affected fetal growth and number of pups born (31). In the present study, however, exposure to 1000 ppm OP during pregnancy did not have any of these effects, which was thought to be due to the very weak estrogenicity of OP (20-22). Endocrine disrupting conditions, such as pituitary weight increase, absence of CL and abnormal lactations in mammary glands, were seen in female pups with fetal exposure to DES, while PE was not evident (18). In this study, no endocrine disrupting conditions, such as PE, anovulatory ovaries and abnormal lactations in mammary glands were seen in female pups with fetal exposure to OP. Fetal exposure to 10 ppm DES from day 13 of pregnancy has been shown to slightly decrease the uterus weight of pups, while causing no evident atrophy (18). In the present study, exposure to 1000 ppm OP throughout pregnancy period also significantly decreased the uterine weight of pups, without evidence of atrophy. The precise mechanisms of these findings remain to be resolved, but it may be considered that estrogenicity of OP during pregnancy affects the gonadotropin-secreting system of pups, resulting in the decreased uterus weight of pups. This is based on the observation that neonatal daily administrations of DES reduced uterine weight possibly due to a disturbance in the gonadotropin-secreting system (7, 9).

Incidence of rats with mammary carcinomas (MCs) and number of MCs/rat. * p<0.05: significantly different from Group I.
Fetal exposure to DES has been shown to enhance the early incidence and number of MCs (18). In the present study, fetal exposure to OP during pregnancy also enhanced the early incidence and number of MCs. It was speculated that OP exposure to pregnant rats before the development of mammary hillocks may be more sensitive to the induction of MCs. This speculation was based on the observation that fetal exposure to OP from day 13 of pregnancy did not enhance the induction of MCs. However, it is necessary to investigate how OP exposure to pregnant rats from 0 to day 13 of pregnancy affects the induction of MTs induced by DMBA in female pups. Terminal end buds (TEBs) in mammary glands 50 days after birth (DMBA administration period) in rats with neonatal administration of 0.1 and 1 μg DES increased, resulting in a stimulatory effect on the initiation of MCs (8). In the study presented here, it is possible that fetal exposure to OP increased TEBs 50 days after birth, resulting in a stimulatory effect on the initiation of MCs. It is necessary, however to further investigate whether fetal exposure to OP affects the development of TEBs.
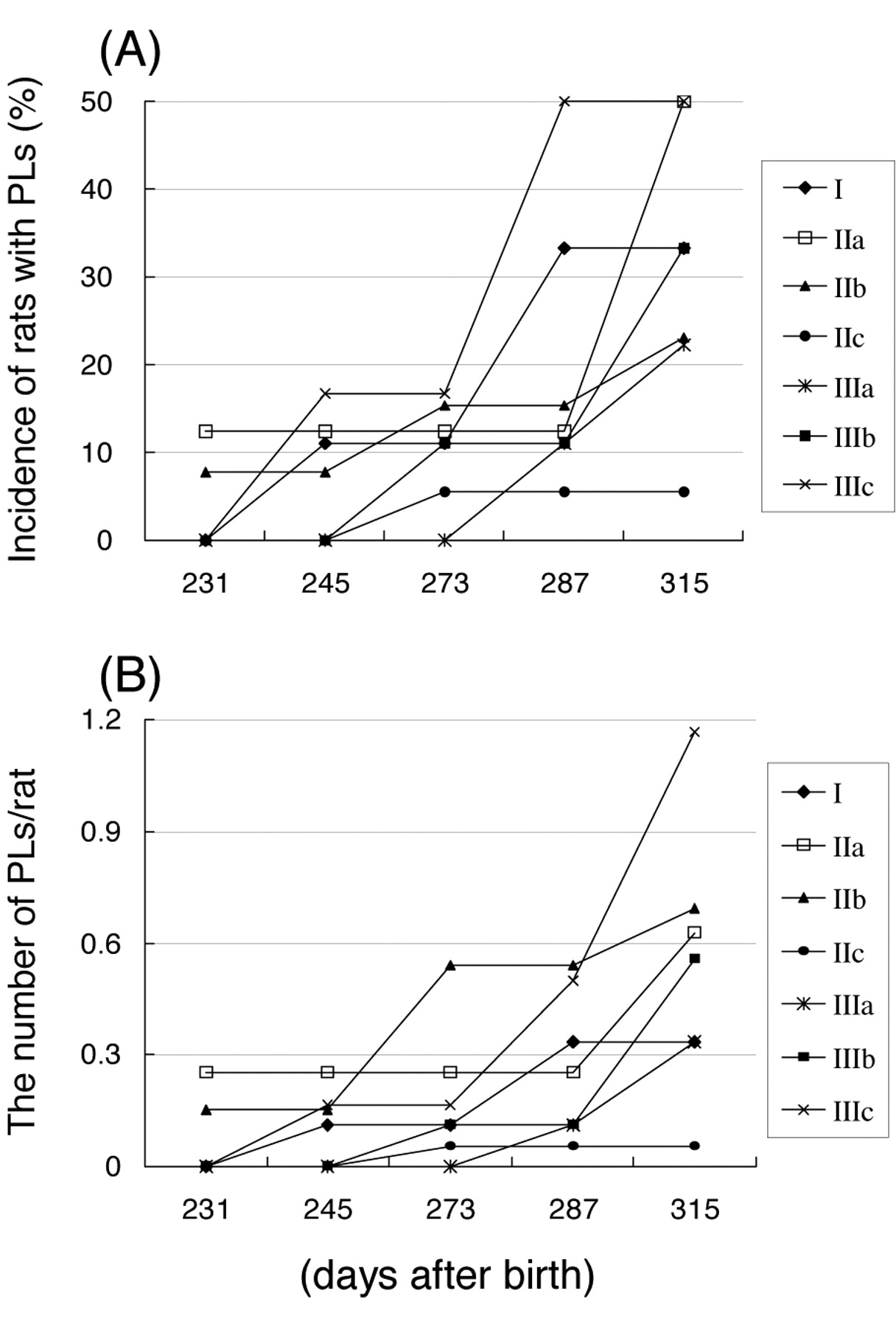
Incidence of rats with proliferative lesions (PLs) and number of PLs/rat.
Fetal exposure to DES enhanced the late incidence and number of PLs, possibly due to hormonal conditions with a relative excess of estrogen (18). In the present study, fetal exposure to OP did not induce abnormal hormonal conditions, such as a relative excess of estrogen, as there was no evidence of the promotion or inhibition of PLs.
Alternatively, TEBs differentiate to more mature structures, namely, alveolar buds (ABs) and lobules, which are less susceptible to carcinogens in rats (32, 33). TEBs that had already differentiated into ABs before DMBA administration did not develop carcinomas, but remained unmodified or underwent dilatation, giving rise to hyperplastic lobules (34). It is possible, however that dysplastic lobules may develop carcinomas because masses consisting of both MC and PL were found and had long latent periods.
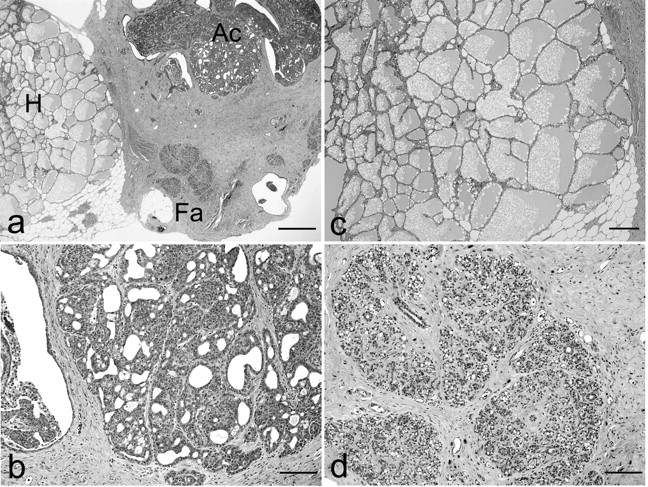
Benign and malignant mixed mammary tumors in group IIIb. A mass composed of adenocarcinoma (Ac), hyperplasia (H) and fibroadenoma (Fa) can be seen (a). Ac: Papillary and tubular proliferations without interstitial invasion can be seen in ducts (b). H: lobular hyperplasia consists of large lobular structures composed of acini and the lumens of acini are dilated and filled with acidophilic secretions (c). Fa: Rich hyalinized stroma and developed acini can be seen (d). Bar=500 μm (a), 100 μm (b, d) and 200 μm (c). HE stain.
In conclusion, these results suggest that fetal exposure to the very weak estrogenicity of OP can enhance the induction of MCs, with no effect on the induction of PLs.
Acknowledgements
We are grateful to Mr. T. Kodama for his valuable technical assistance.
- Received November 18, 2009.
- Revision received April 7, 2010.
- Accepted May 28, 2010.
- Copyright © 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.