


Abstract
Phenothiazines have their primary effects on the plasma membrane of prokaryotes and eukaryotes. Among the components of the prokaryotic plasma membrane affected are efflux pumps, their energy sources, energy providing enzymes such as ATPases, and genes that regulate and code for permeability aspects of the bacterium. The responses of multi-drug (MDR) and extensively drug resistant (XDR) Mycobacterium tuberculosis to the neuroleptic phenothiazine thioridazine are reviewed. The information collated suggests that this phenothiazine has the potential to cure XDR and MDR tuberculosis infections, a potential that has been recently demonstrated by its ability to cure 10 patients who presented with XDR TB infections. The mechanism by which this phenothiazine produces the desired effects within the infected macrophage is also discussed.
- Phenothiazines
- thioridazine
- pulmonary tuberculosis
- MDR TB
- XDR TB
- human macrophage
- enhanced killing activity by macrophage
- efflux pumps
- therapy with thioridazine
- review
The development of antimicrobial agents for the therapy of infection began in the late 19th century with the pioneering work of the German scientist Paul Erhlich. The studies conducted by Erhlich demonstrated that the newly created dye the phenothiazine methylene blue had activity against infectious bacteria and parasites (1). Previously, Bodoni had shown that the dye when administered to humans caused them to become lethargic (2) and although this study was not noticed for over 40 years, a similar conclusion was reached by others with the use of a phenothiazine antihistamine (3). However, the neuroleptic properties of the antihistamine were weak and attention was now focused on the methylene blue itself. Interest in this dye continued primarily as a result of the work by Perkin who had created the first chemically synthesized dye mauve some 50 years earlier and because of its commercial value, caused many to focus on the creation of additional dyes (4). Among the many discoveries obtained with the study of methylene blue was the development of dyes that could stain living tissue-vital stains. Because these dyes were derived from the phenothiazine methylene blue and they also inhibited motility of parasites, Roehl, a student of Erhlich, developed a canary model for malarial infection with Plasmodium yoelii and successfully treated the infected canary with a phenothiazinium salt created by Schulemann in 1932 by a chemical process that altered the side chain of methylene blue (5). However, the phenothiazinium salt was still blue and its use for human malaria infections would turn the patient an undesirable blue.
It remained for Charpentier in 1957 to develop a method by which the colour of methylene blue derivatives could be eliminated, and among these derivatives, was chlorpromazine which retained the neuroleptic properties of the original dye and which, for the next two decades was the primary chemotherapeutic agent for the control of psychosis (6). As a consequence of the wide use of chlorpromazine (CPZ) it was noted that the frequency of infections within psychiatric wards, oftentimes kept under improper hygienic conditions, were far fewer than that observed for other more sanitized areas of the hospital (7).
These observations prompted studies for the next 30 years after the introduction of CPZ that investigated the in vitro activities of CPZ against bacteria (8-27) and in vivo activities of CPZ (11, 12, 15-17, 28-37). CPZ was shown to inhibit the in vitro growth of a wide gamut of bacteria (9, 13, 15, 18-21, 23-25, 27, 38, 39), inhibit the growth of mycobacteria (9), inhibit the secretion of toxins (14-17, 21), cause the elimination of plasmids from infected Gram-negative bacteria (8), cause the lysis of bacteria (22), increase the permeability of bacteria to agents (25), reverse resistance to antibiotics (26) and inhibit bacterial enzymes (8, 23). Although all in vivo activities were produced by concentrations that were well beyond those that can be achieved clinically (40), and these high concentrations inhibited immune functions (13, 28, 30), the activities produced in vitro could be reproduced in vivo. Consequently, administration of CPZ to mice reduced production and effects of endotoxins by bacteria (11, 12, 15-17), cured the mouse of experimental infections (12, 29, 30-36). Although CPZ was undoubtedly shown to have potential for the therapy of bacterial infections, there was little interest during these 30 years post-introduction of CPZ due to the fact that this was the Golden Period of Antibiotics and there was no need for the use of an agent that was developed and extensively used for the therapy of psychosis. Moreover, because the FDA has a policy of ‘one drug for one disease’ the use of a neuroleptic as an antimicrobial agent was in effect a ‘no-no’.
The Threat of Multidrug-resistant Infections and the Chemotherapeutic Potential of Phenothiazines
Resistance to penicillin antibiotics was seen soon after the use of penicillin (40) and with the introduction of new antibiotics and antimicrobial agents and their extensive use and misuse, mono-resistance to these antibiotics rapidly followed (40-44). Resistance of a bacterium to two or more antibiotics (multi-drug resistance) was a rare event until the early 1980s (45, 46), frequent during the late 1980s (47-50) and common place during the 1990's (51, 52). With the piling up of evidence that the frequency of multidrug-resistance (MDR) would continue to increase (52), particularly in strains of Mycobacterium tuberculosis (Mtb) resistant to as many as 4 to 5 first-line of defence drugs (53) some focus on the wide gamut of the in vitro activities of phenothiazines against antibiotic susceptible (53-58) and resistant (9, 53, 59, 60) strains of Mtb were demonstrated. However, because these activities took place at concentrations that were clinically irrelevant (37, 61) interest in the antitubercular properties of phenothiazines remained mild. The demonstration by Crowle's (61) group that the phenothiazine CPZ enhanced the killing of intracellular Mtb increased interest in phenothiazine. However, because the side-effects of this phenothiazine are serious and frequent (62), no attempt to use this compound for the therapy of pulmonary tuberculosis ensued. Nevertheless, interest was renewed with the demonstration that the milder and phenothiazine thioridazine (TZ) was equally effective against mono-resistant Mtb, MDR and extensively drug-resistant (XDR) (resistant to 5 primary defence drugs) strains as was CPZ (59, 60). Moreover, other milder phenothiazines such as antihistamines were almost as effective as TZ (60). Following the macrophage protocol of Crowle, exposure of macrophages containing phagocytosed MDR Mtb resulted in the enhanced killing of the bacterium at concentrations in the medium that were below those used for the chronic therapy of psychosis (63). These studies were soon followed by demonstrations that mice infected with antibiotic-susceptible (64) and antibiotic-resistant Mtb could be readily cured with TZ (65). The curative ability of TZ and its much milder side effects coupled with over 40 years of safe usage, propose that at the very least, XDR TB patients whose prognosis of mortality was certain may be considered for therapy with TZ under ‘a compassionate rationale’ (66, 67).
Before discussion of the mechanism by which phenothiazines enhance the killing of intracellular bacteria, the targets of phenothiazines will be discussed.
Targets of Phenothiazines
1. CPZ inhibition of energy-dependent membrane functions of eukaryotes and prokaryotes. Most of the studies on the antimicrobial activity of phenothiazines have been conducted with CPZ. Nevertheless, the mechanisms defined for CPZ have been essentially demonstrated for other phenothiazines when studied. Therefore, for practical purposes we will at this time confine ourselves to the mechanisms by which CPZ affects bacteria in vitro, ex vivo (intracellular) and in vivo.
CPZ inhibits the binding of calcium to calcium-binding proteins of eukaryotes (60, 68, 69) and prokaryotes (58, 70-76). As a consequence of its calcium-binding properties, calcium-dependent enzymes of eukaryotes are inhibited. Among these are enzymes that provide energy (77-79) such as kinases (80-83) and phosphatases (84-87). The inhibition of energy-producing enzymes, most of which are on or close to the plasma membrane of eukaryotes, reduces or obviates numerous plasma membrane functions. Among the plasma membrane functions inhibited are phagocytosis of bacteria (88, 89), ABC transporters of MDR normal and cancer cells (90-97), and transporters of calcium (80, 98-100) and potassium ions (101-105). With respect to prokaryotes, CPZ inhibits the binding of calcium to calcium-binding proteins (106, 107) and mycobacteria (108-111), inhibits bacterial kinases (87, 107, 112-114) and phosphatases (115), calcium (8) and potassium transport (116, 117). In other words, the activity of the phenothiazine on energy-dependent membrane functions of eukaryotes and prokaryotes is very similar.

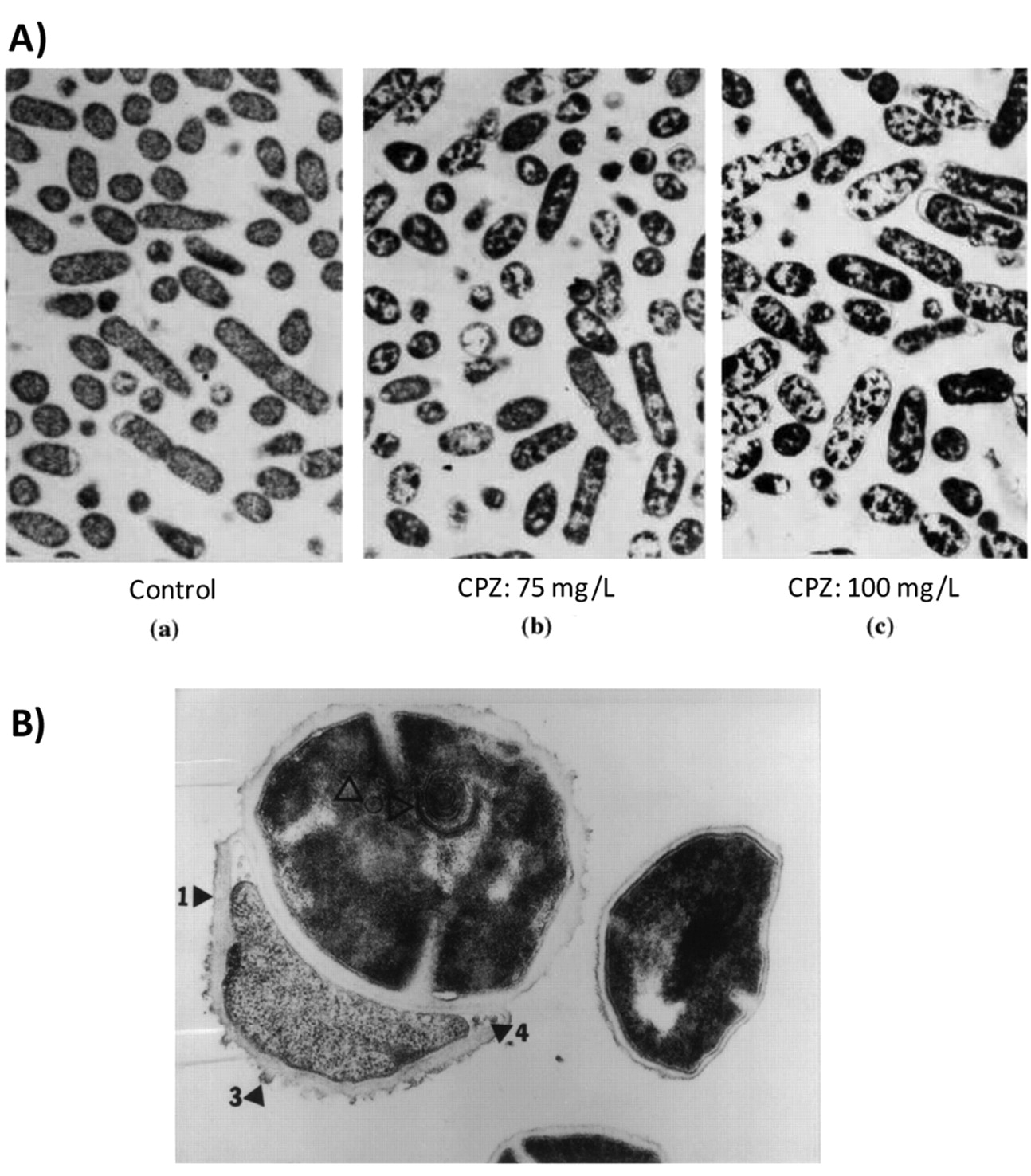
A) The effect of increasing concentrations of CPZ on ultrastructure of Salmonella typhimurium (×10000). a: Control with homogenous cytoplasm; b: with chlorpromazine at 75 mg/l; c: with chlorpromazine at 100 mg/l. (121). B) The effect of chlorpromazine on the ultrastructure of S. aureus exposed to 30 mg/l for 6 h. Note thickened cell wall (1 and 3), thickened cross wall (4), and lamellae (2) (pointers inside cell) (×82,000) (ref. 6).
2. CPZ and the bacterial cell envelope. The earliest activity of a phenothiazine such as CPZ on bacteria takes place at the level of the cell envelop inasmuch as this structure is first encountered by the agent. The activities affected are primarily those that are involved in energy dependent transport processes as shown in the previous section. In vitro exposure of bacteria to CPZ increases the permeability of the organism to antibiotics (118-121) and as a consequence, reduces antibiotic resistance (118-121). An example of the effect of CPZ on the cell envelope of Gram-negative and Gram-positive bacteria is provided in Figure 1. Briefly, the cell envelope which is crenated in the control Salmonella becomes smooth after a 16 hour exposure to a sub-inhibitory concentration of CPZ; in vitro exposure of Staphylococcus aureus to the same phenothiazine causes deterioration of the cell envelope and subsequent lysis of the organism. The effects on the cell envelop are preceded by elongation and filamentation of the Gram-negative bacterium as shown in Figure 2. The phenothiazine also has other cell envelope effects that include an ability of the phenothiazine to reduce antibiotic resistance or to reverse resistance to two or more antibiotics (122-124). Resistance is achieved through the inhibition of a multi-drug efflux pump system of the organism which insures that antibiotics and other noxious agents are extruded prior to their reaching their intended targets. This subject is discussed in the following section.
3. Bacterial efflux pumps and their inhibition by phenothiazines 3a. Bacterial efflux pumps involved in multi-drug resistance. The efflux pump systems of bacteria are genetically classified into five groups and sub-classified as to the immediate source of the protons that energize the pump (122). Efflux pumps that utilize a proton from the hydrolysis of ATP directly are members of the ABC transporter super family (125). These transporters consist of 6 or 10-12 domains that span the plasma membrane; one domain recognizes the agent to be extruded and another has a binding site for ATP and its subsequent hydrolysis that results in two protons (126, 127). The protons generated energize the pump and the substrate is exported to the outside of the cell. Bacterial ABC efflux pumps that transport potassium and calcium are readily inhibited by phenothiazines (128-130). The substrates extruded by ABC efflux pumps are presented in Table I.
Efflux pumps that are energized by protons under the control of the proton motive force (PMF) are genetically classified into four families: the major facilitator superfamily (MFS) which contains over 1000 transporters and therefore is the largest of the families; the multi-antibiotic toxin extrusion family (MATE); the small multi-drug resistance family (SMR); and the resistance nodulation division family (RND). The PMF results from metabolic activity of the bacterium that generates protons which are transported to and distributed to the surface of the cell (131). With respect to Gram-positive bacteria, the concentration of surface bound protons is greater than that at the medial side of the plasma membrane due to their binding to proton loving lipids of the lipopolysaccharide of the cell envelope thereby resulting in a proton gradient (132, 133). The protons on the surface are made available to the MFS, MATE and SMR efflux pumps all of which can extrude antibiotics and other noxious agents (134-136). With respect to Gram-negative bacteria, which have a more complex cell envelope consisting of an outer membrane and an inner membrane (plasma membrane) separated by the periplasmic space, the protons on the surface of the cell envelop are also attracted to the lipopolysaccharide components that love protons and these are translocated to the periplasmic space and used for the energizing of the RND efflux pump. The concentration of surface bound protons is much greater than that in the periplasm and therefore an electrochemical gradient is maintained that is defined as the PMF. The electrical potential of the PMF can be maintained at extreme pH and temperature (137-140).
Relationship of transporters of bacterial efflux pumps to antibiotics extruded. Adapted from VanBambeke et al. (223).
3b. Structure of an RND mdr efflux pump. The structure of RND efflux pumps of Gram-negative bacteria are similar and consist of three protein units: a TolC protein that provides a conduit from the transporter to the surface of the cell; a transporter protein that is attached to the plasma membrane and exports substrates from the outer leaflet of the plasma membrane directly to the attached TolC unit (141, 142); and, two fusion proteins that flank the transporter unit and assist the transport of the substrate through the TolC channel by peristaltic action (141, 142). Figure 3 depict the structural relationship of the units of the AcrAB-TolC efflux pump and the peristaltic mechanism behind the extrusion of the substrate. Because the most studied RND efflux pump of Gram-negative bacteria is the AcrAB pump present as the main efflux pump of E. coli (142) and Salmonella (143, 144) we will focus our discussion on the AcrAB system.
3c. Genetic regulation of RND efflux pumps. Permeability of Gram-negative bacteria to antibiotics is controlled by the combined roles of porins and efflux pumps (145-150) and the lipopolysaccharide layer of the outer membrane (151-155). Porins are tribarrel structures that provide a conduit through the outer membrane to the periplasm of the cell. Lipophobic antibiotics use these channels for penetrating and eventually reaching their intended targets. The number of porins used by lipophobic antibiotics of some clinical MDR Gram-negative isolates are reduced (148, 150) suggesting that the MDR nature of these bacteria is due to down-regulation of porins, primarily of the outer membrane protein F (ompF).
The MDR phenotype of an increasing number of Gram-negative clinical isolates is due to the over-expression of the main efflux pump of the organism (156, 157). The efflux mediated Gram-negative MDR phenotype is believed to be the result of mis-use or over-use of antibiotics (158-162). Experimentally, exposure of E. coli to increasing concentrations of tetracycline results in increasing resistance to tetracycline and other unrelated antibiotics, accompanied by an increased activity of the acrB transporter gene (163). Induced resistance can be reversed with a phenothiazine and by transfer to drug-free medium (163, 164). Evaluation, by real-time RT-PCR, of stress, local regulators and transporter coding genes indicates a cascade of genes that are increased in activity at each level of induced resistance to the antibiotic (164). Similar results have been obtained by others who employed the same approach for ciprofloxacin-induced resistance of Salmonella (165).

Elongation of E. coli exposed to 100 mg/l of chlorpromazine for 24 h. The example shown in this figure is indistinguishable from the response of E. coli to β-lactams to which it is sensitive (6).
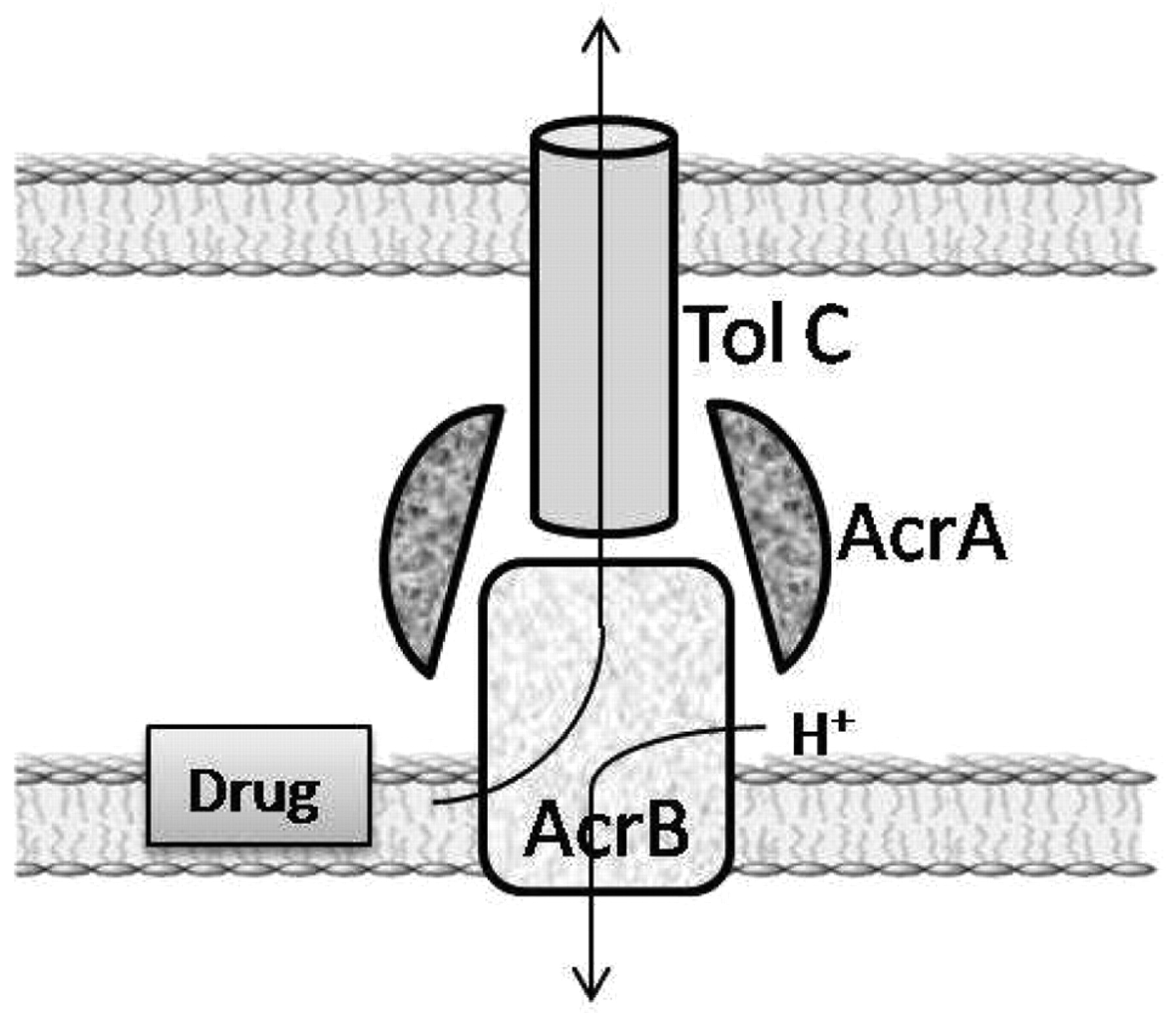
Schematic of structure of AcrAB-TolC efflux pump and the relationship of its components to the cell envelope. Proton of periplasm activates transporter and is subsequently translocated to the cytoplasm.
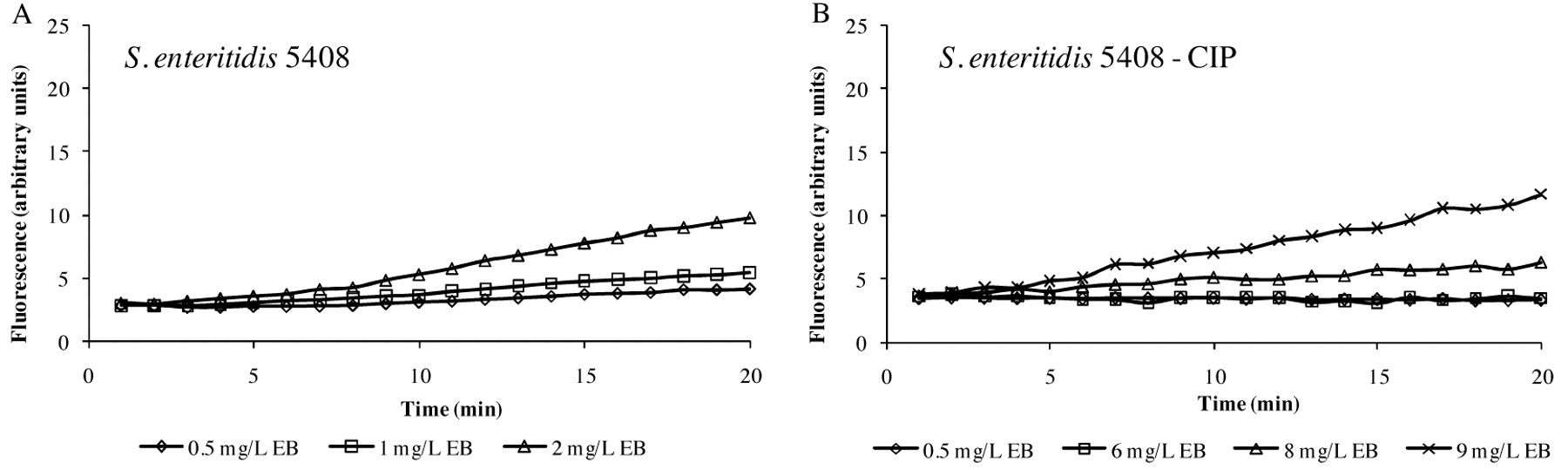
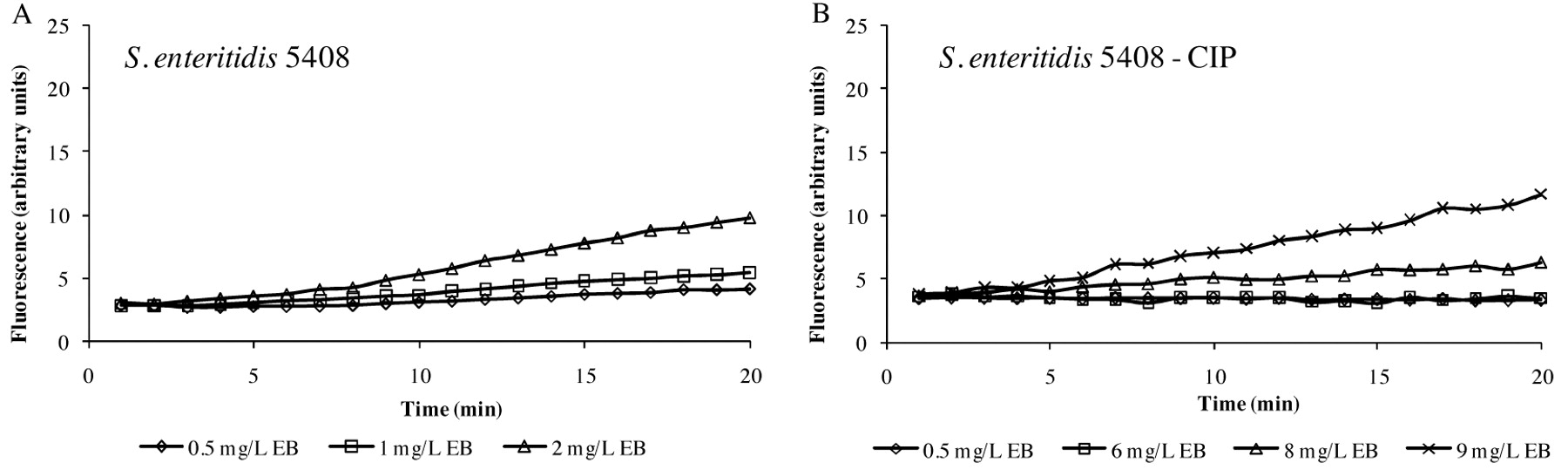
Accumulation of different concentrations of EB by S. enteritidis 5408 strain (A) and its progeny adapted to ciprofloxacin - S. enteritidis 5408CIP (B). The data are presented the difference between the accumulation data of the strain more EB and the EB alone, which has a background especially at higher concentrations.
Interestingly, inducing MDR in E. coli with increasing concentrations of a given antibiotic, results in the over-expression of the ompF gene that is accompanied with a reduction of ompF units (164). Because of an increased activity of genes that code for the main proteases of E. coli, the reduction of porin F units is not the result of a down-regulation of ompF, but of the degradation of ompF prior to it being inserted into the porin unit (164). The regulation of genes that affect the permeability (porin and efflux pumps) of bacteria to antibiotics will be further discussed in section 3d.
3d. Physiology of the AcrAB efflux pump. The demonstration of an efflux pump system of individual bacterial species and strains has been routinely conducted with the use of the fluorochrome ethidium bromide (EB) (166-172), a universal efflux pump substrate. A new automated system that employs EB for the study of a given efflux pump system has been developed and provides a real-time assessment of accumulation and efflux of EB under physiological conditions (125, 173, 174). Consequently, one may program for the desired temperature for conducting the assay (isolates obtained from poultry may have temperature optima that differ from bacterial isolates from the human host); and employ medium whose condition may have an effect on accumulation/efflux (ionic strength, pH, inhibitors, etc.). This assay has previously been described in detail (125, 173, 174).
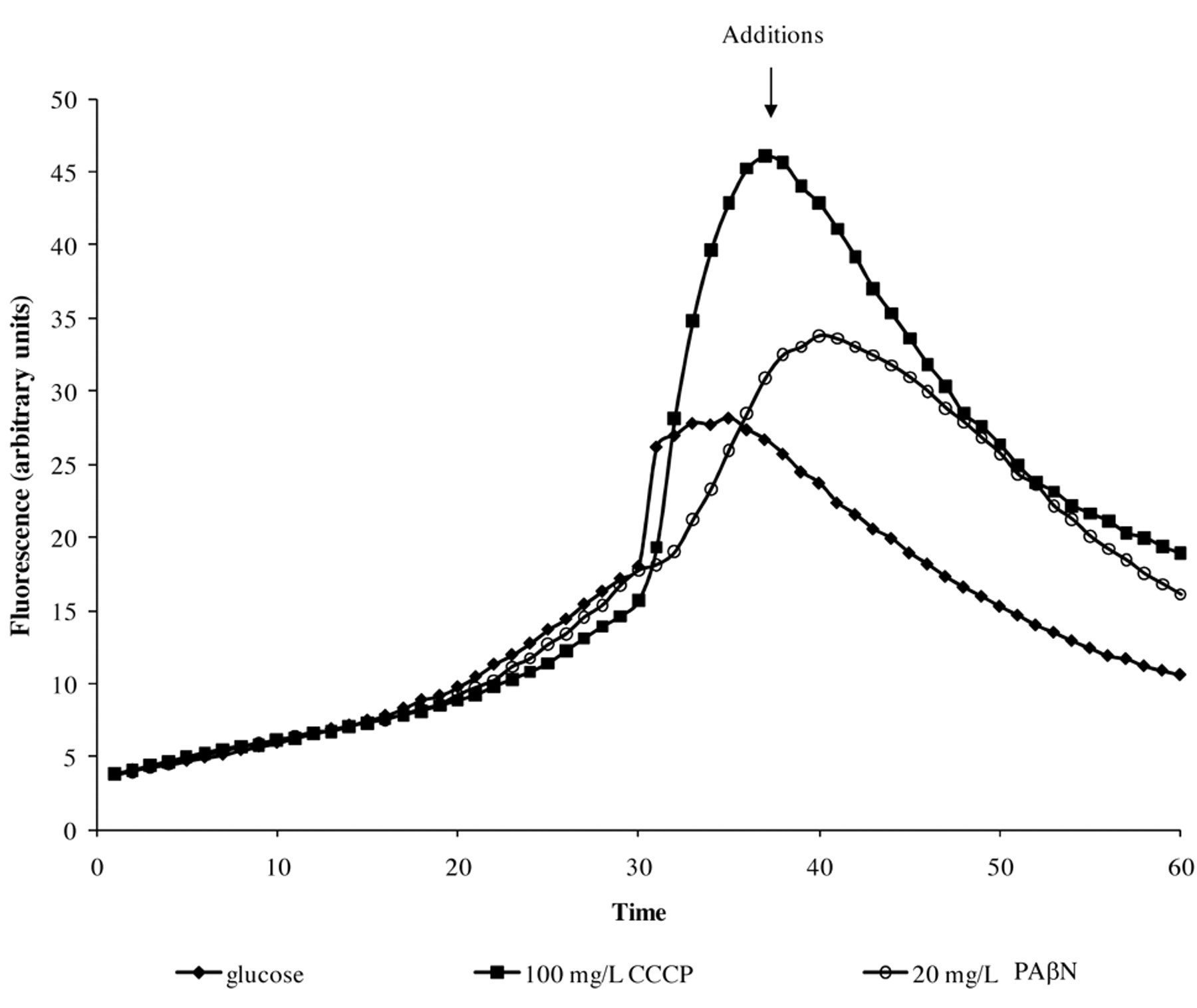
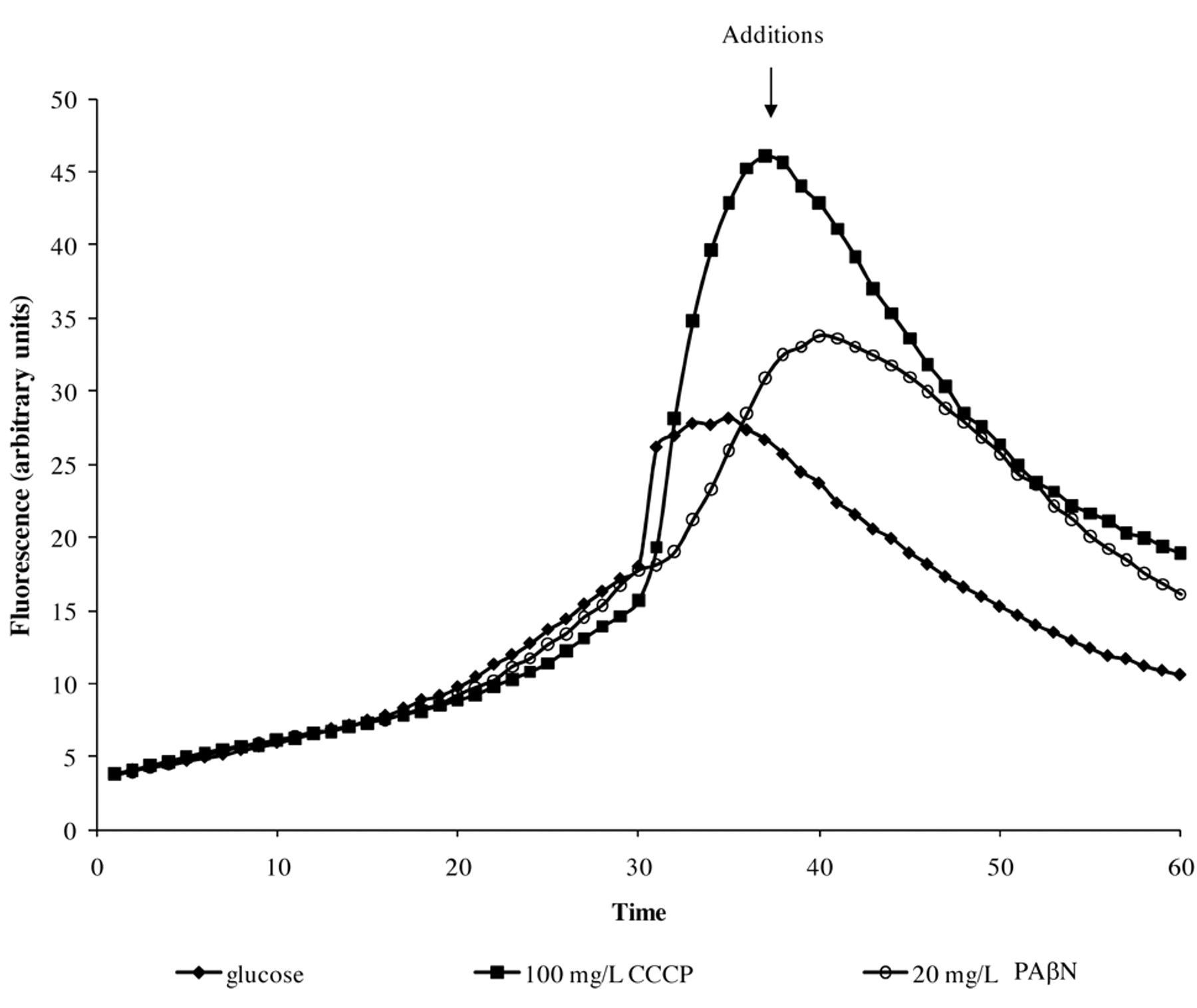
The first consideration that may be explored via the automated EB system is the different abilities of individual strains of a given species to extrude EB. As shown in Figure 4, glucose-containing saline of pH 7, the maximum concentration of EB that the S. enteritidis 5408 can barely handle (extrude) is 1 mg/l of EB (Figure 4A), whereas the S. enteritidis 5408CIP strain that has been induced to high level resistance to tetracycline can handle up to 5 mg/l of EB after which EB begins to accumulate (Figure 4B). The effect of pH is to modulate the accumulation of EB as shown in Figure 5. In this case, the example given is for E. coli K-12 AG100 in glucose-containing medium of pH 5, 7 and 8 and 1.0 mg/l of EB. As evident in Figure 5, at pH 8 accumulation of EB is greatest, however, at pH 5, accumulation of EB is absent (flat). The absence of accumulation at pH 5 is due to an active intrinsic efflux pump system. The following experiment was conducted: after 15 minutes of incubation of E. coli K-12 AG100 in glucose-free medium of pH 5 and 1 mg/l of EB, the instrument was paused, the proton ionophore carbonyl cyanide m-chlorophenylhydrazone (CCCP) was added, and the instrument restarted. As evident in Figure 6, the addition of CCCP immediately increases sharply the amount of fluorescence-thereby demonstrating the presence of an efflux pump system that is fully operational at pH 5. Similar results have been obtained with Salmonella, Enterococcus faecalis and Enterobacter aerogenes (125).
An EB microplate assay has been developed that utilizes the same concept behind the EB agar method (175, 176) but rather than agar, an equivalent broth medium is used in conjunction with a 96 microwell plate (177). Figure 7 provides evidence that the accumulation of EB by E. coli strains that differ by the degree of AcrAB efflux pump expression is modulated by pH (178).
Other methods have been developed for the study of efflux of EB. As an example, the EB agar method (177) has been modified to accommodate as many as 12 strains for purposes of distinguishing differences in their efflux capacities at differing pH. As evident in Figure 8, pH modulates accumulation of EB by salmonella strains that differ in the expression of their efflux pump AcrAB.

pH modualtion of accumulated EB by AG100 and the effect of glucose.
3e. Effect of the phenothiazine TZ on genes that control the permeability of Salmonella. In vitro exposure of Salmonella to CPZ inhibits growth for the first 8 hours after which the organism becomes increasingly resistant (121). As evident in Figure 9, TZ also produces the same effect. Further studies to investigate the growth inhibition and resistance of the organism to the agent at the gene level, were carried out using real-time RT-PCR. The effect of TZ on the stress genes soxS and rob, the global regulator of permeability ramA, the local regulator of efflux pumps marA, the transporter gene acrB, the two-step regulon pmrA and pmrB, and the two step regulon phoQ and phoP of Salmonella after zero time, 0.5; 1.0; 4.0, 8.0 and 16 hours was determined. As evident in Figure 10, the first gene that is significantly activated within an hour relative to that of the control (not exposed to TZ) is the stress gene soxS, after which time it is relatively inactive for the duration of the culture. The activity of the global regulator gene ramA is increased within an hour and continues to increase for the next 7 and a half hours. By the end of the culture period, ramA activity is equal to that of the control. The local regulator marA is highly active but only for a brief period (after 4 hours). The transporter gene acrB achieves a very high level of activity by the end of 8 hours and although its activity decreases by the end of the culture period, it remains the most active gene. After 8 hours of exposure to TZ the strain begins to grow. The activities of the pmrA and pmrB commensurate with the sensor role known for pmrB which precedes the increased activity of pmrA (179). Clearly, the sequential and coordinated activities of soxS, ramA, marA and acrB are responsible for the ensued resistance of Salmonella to phenothiazine.

pH modulation of CCCP effects on efflux by E. coli AG100. Legend in the graph corresponds to the conditions of the additions. Accumulation (first 25 min) was carried at pH5 in absence of glucose.

pH modulates accumulation of EB by E. coli strains that differ in the expression of their AcrAB efflux pump.
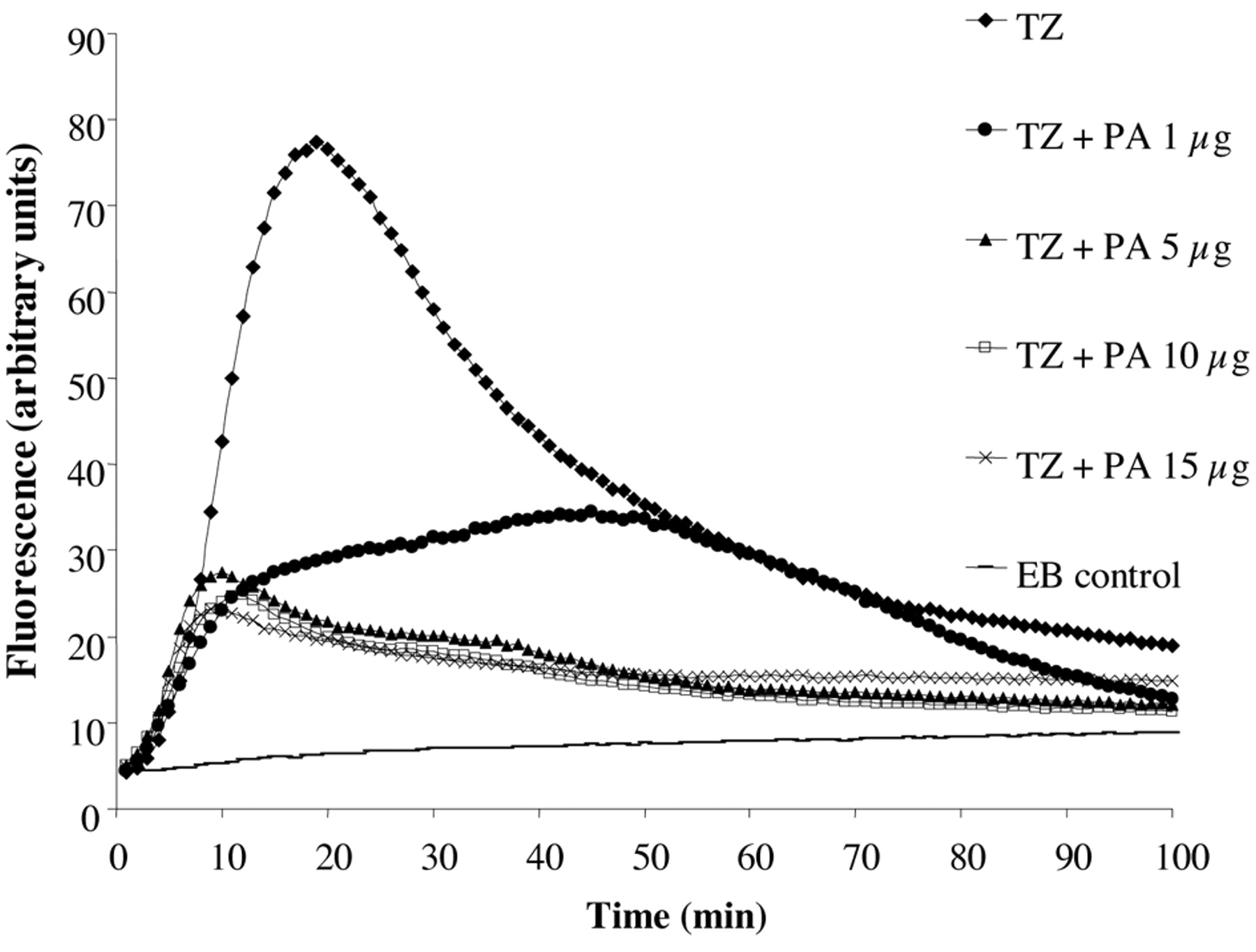
The application of the automated EB method for the assessment of the TZ effects on the accumulation of EB by salmonella was conducted in glucose free saline pH 8 (179). As shown in Figure 11 exposure of Salmonella 104 to 50 mg/l of TZ immediately promotes accumulation of EB. Surprisingly, approximately after 20 minutes a peak of accumulation is reached, followed by a decrease in fluorescence-indicating efflux. Because no metabolic energy was present in the glucose-free saline, the efflux response noted could not be readily related to any significant change in gene expression. Consequently, the question was asked as to the source of energy that afforded efflux. Salmonella, when grown in the absence of glucose, will switch its source of energy to fatty acids. Therefore, the assay that demonstrated TZ promoted accumulation followed by efflux was repeated with additions of increasing concentrations of palmitic acid. Figure 11 serves to illustrate that the accumulation of EB could be significantly reduced and hence, the process of efflux was not needed. Because increasing concentrations of EB do significantly increase accumulation at pH 8 which is not followed by efflux, we can conclude that the efflux noted after TZ promoted accumulation is not the result of the activation of the efflux pump as a consequence of accumulated EB. Interestingly, if at the peak of TZ accumulation CCCP or the common inhibitor of an RND efflux pump PAβN (phe-arg-napthylamide) is added, TZ promoted efflux is not affected (Figure 12).

Effect of pH on the minimum concentration of EB that produces fluorescence of Salmonella strains. Note that sal 5408 does not fluoresce with concentrations of EB as high as 3 mg/l.

Effect of concentrations of thioridazine on the growth of Salmonella. Growth curve of Salmonella enteritidis 104 in the presence of TZ 100 mg/L (Mueller-Hinton) at pH 7.4.
3f. Antimicrobial activities of phenothiazines. The activity of TZ against Salmonella discussed in the previous section takes place at sub-inhibitory concentrations. At much higher concentrations, the growth of the organism is inhibited after a 16 to 24 hour period. Comparison of the minimum inhibitory concentration (MIC) of CPZ, the phenothiazine most studied with respect to its antimicrobial properties against bacteria, is summarized by Table II. Among the most resistant species to CPZ are the Gram-negative bacteria, with Salmonella strains being the most resistant followed by Enterobacter, E. coli, Shigella and Klebsiella. Among the less resistant bacterial species are Gram-positive ones such as Mycobacteria, Staphylococci, Enterococci and Vibrio. Regardless of the degree of sensitivity to the phenothiazines, the MICs are well beyond the level that can be achieved in the patient, namely, 0.5 mg/L of plasma (181-183).
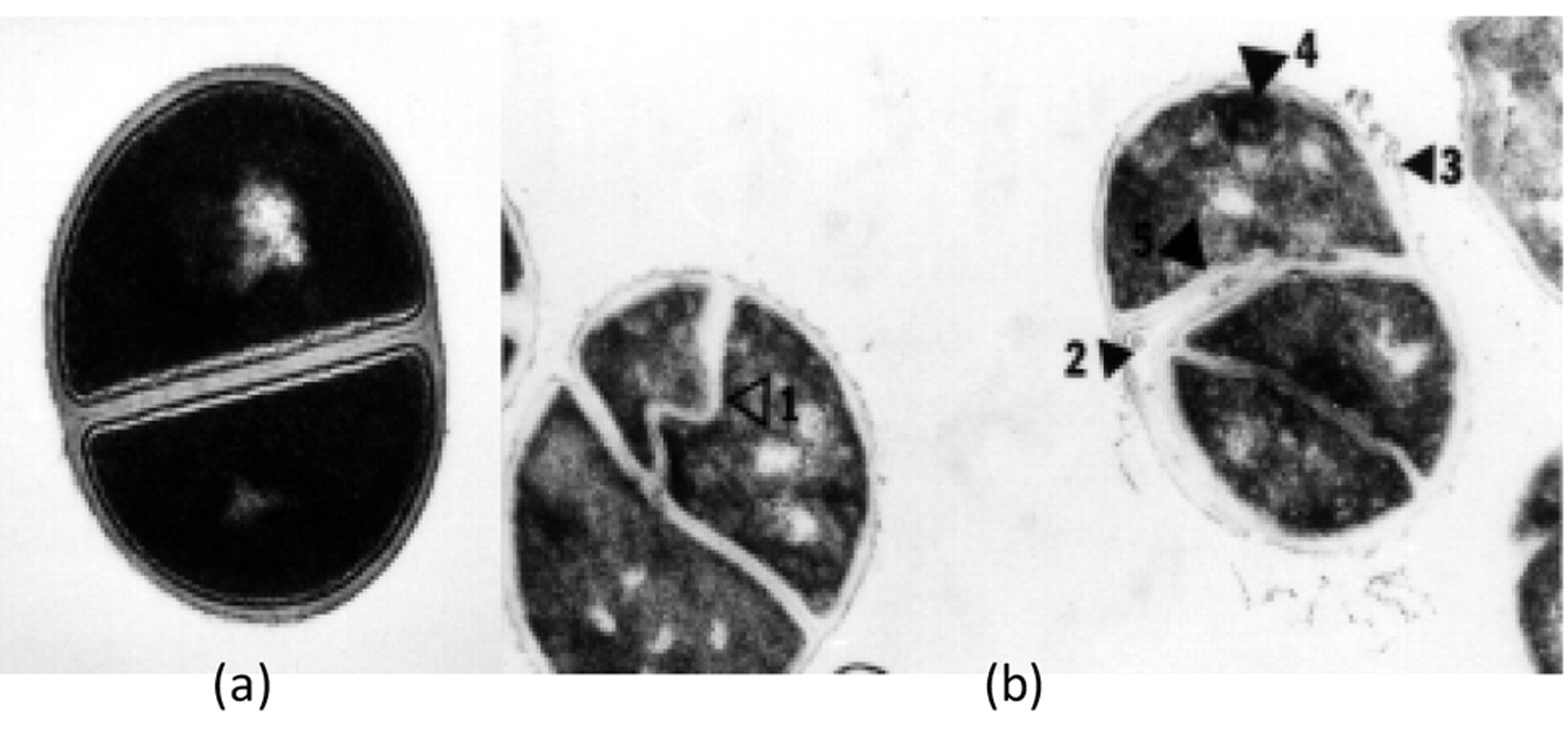
The earliest morphological response of Gram-negative bacteria to CPZ is the development of filamentation (184) as illustrated in Figure 2. Filamentation that is caused by exposure to sub-inhibitory concentrations of a beta-lactam has been shown to be the result of the antibiotic binding to penicillin binding protein 3 (185). However, filamentation can be promoted by exposure to non-beta lactam antibiotics such as nalidixic acid, novobiocin, oxolinic acid and nitrofurantoin (184, 186) by cyclic AMP (187-189), by heat (190), and by many other environmental conditions which threaten the organism. This suggests that the reponse of filamentation is one of adaptation that affords survival (191). The response of Gram-positive bacteria to CPZ is similar to the response to beta-lactams as illustrated in Figure 13. Beta-lactams at subinhibitory concentrations inhibit the hydrolytic enzymes that would normally cause the separation of the replicated cells from each other (192, 193). For filamentation, the end product is the containment of as many as 64 genomes within a common cytoplasm; for the cluster, as many as 64 cells have been shown to make up the cluster. The mechanism by which a phenothiazine induces filamentation and cluster formation of Gram-negative and Gram-positive bacteria are not known. However, at considerably higher concentrations of the phenothiazines, the agent readily intercalates between nucleic bases, primarily between areas of the DNA rich in guanosine and cytosine bases (194, 195). Moreover, similar concentrations of the phenothiazine inhibit gyrase (196) and promote direct relaxation of supercoiled DNA (196). At first it is difficult to reconcile the effects on gyrase with that on supercoiling. Quinolones that target gyrase and affect supercoiling of DNA also promote filamentation, therefore it may be assumed that filamentation promoted by CPZ involves gyrase and relaxation of supercoiled DNA. Because relaxation of supercoiled DNA affords transcription (197) and a number of genes are involved in induced filamentation of E. coli (188) it is reasonable to expect that at least the nucleotide sequence of fic and fic1 genes are targets of the phenothiazine. Recently published studies have shown that TZ reduces the transcription of the MecA element of MRSA and its coded product PBP2a (198) and that CPZ reduces the activity of the global regulator ramA (97). These findings and the fact that TZ, as discussed in section 3d, has effects on the activity of a collection of genes involved in the regulation of permeability of a Gram-negative bacterium indicate that the relationship of specific genes to the induced phenothiazine morphological responses of bacteria will be a subject of intense study.

Assessment of stress, global and local regulators of efflux pumps, acrB transporter and two-step PmrA/B genes of Salmonella cultured in medium containing 100 mg/L of thioridazine by real-time RT-PCR. Quantification of relative expression of regulator genes, stress genes, acrB, pmrA and pmrB genes of S. enteritidis 104 after 0, 0.5, 1, 4, 8, 16 h of exposure to TZ 100 mg/L. Changes in gene expression are relative to the untreated cells, and normalized against the house-keeping gene 16S rRNA.
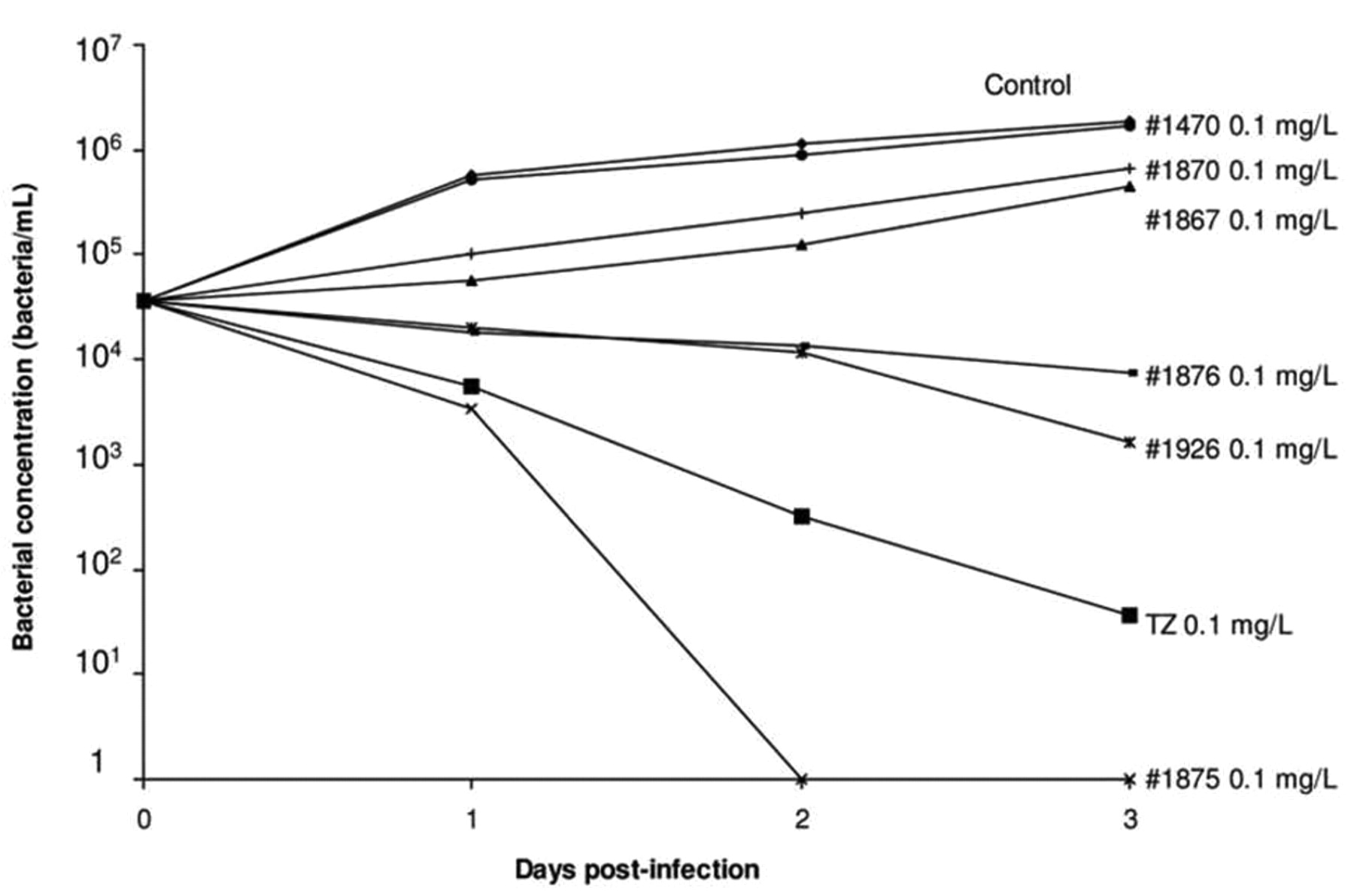
4. The mechanism by which TZ enhances the killing of intracellular Mtb, MDRMtb and XDRMtb. The TZ-enhanced killing of intracellular antibiotic susceptible M. tuberculosis, MDR Mtb and extensively drug resistant XDR Mtb non-killing human macrophages takes place at concentrations of TZ in the medium that are similar to those used for the therapy of psychosis (65). At first, because CPZ is concentrated by lysosomes of the macrophage (199), killing was attributed by our group (65), as Crowle's group (63), to be the result of the phenothaizine being concentrated to a level compatible with that bactericidal concentration demonstrated in vitro (65). TZ inhibits efflux pumps of both prokaryotes and eukaryotes (200), and among the pumps affected are those responsible for the transport of K+ and Ca++ ions that are essential for the acidification of the phagolysosome that results in the activation of lysosome hydrolases (201, 202). The effect of other inhibitors of K+ and Ca++ transport on the killing of intracellular mycobacteria was also examined (203). As shown in Figure 14, these common inhibitors of K+ and Ca++ transport enhanced the killing of intracellular mycobacteria. The mechanism by which TZ enhances intracellular killing was previously proposed (131). Briefly, the binding of the mycobacterium to the plasma membrane of the macrophage results in the invagination of that part of the plasma membrane ultimately resulting in the phagosome. The efflux pumps that transport K+ and Ca++ into the cell are present in the plasma membrane in large numbers (204) and, after the formation of the phagosome, these transporters are now transporting these ions from the phagosome to the cytoplasm of the cell. Exposure of the macrophage that contains the phagocytosed bacterium to TZ, results in the uptake of the phenothiazine via the process of pinocytosis (205). The vacuole that contains TZ and the lysosome fuse with the phagososome. TZ, now diffused throughout the phagolysosome, inhibits the transport of K+ and Ca++ thereby building up the concentration of these ions within the phagosome, and subsequent activation of V-ATPases (206). The activation of the V-ATPases generates H+ protons which decrease the pH of the phagolysosome and the latent hydrolases are now activated (207) and promote the degradation of the bacterium (207-209).
MIC of CPZ against pathogenic bacteria.
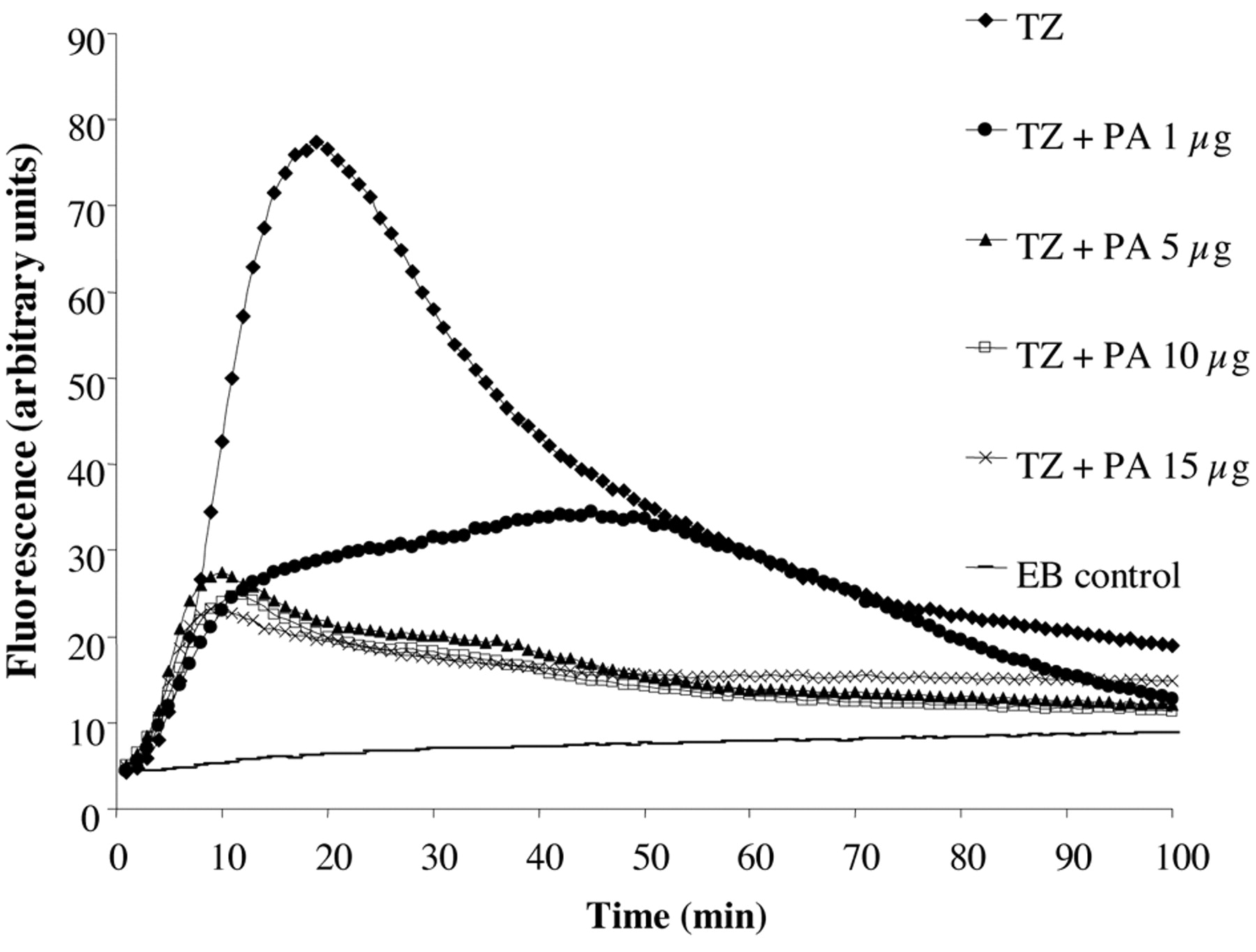
The effect of TZ on the accumulation of EB by Salmonella in glucose-free saline pH 8. Note: In the presence of increasing concentrations of palmitic acid (PA) the effects of TZ on accumulation are significantly reduced.
The effect of CCCP and PAβN on the TZ (50 mg/L) promoted efflux of EB by S. enteritidis 104 at pH8. Addition of CCCP and PAβN, at the peak of TZ induced accumulation, does not affect efflux.

S. aureus and 30 mg/L CPZ. Incubation 6 hours (×50 200). Arrows indicate five distinct types of abnormalities (221).

Effect of TZ and its derivatives in the killing activity of macrophages infected with M. tuberculosis H37Rv ATCC27294. Human macrophages (1×105 cells/mL) were infected with M. tuberculosis at a ratio of 1:10 (1 macrophage:10 bacteria). After 1 hour of phagocytosis, cells were washed to remove non-phagocytosed bacteria. Aliquots of the test compounds were added to the corresponding wells and cells incubated for 0, 1, 2 and 3 days. After incubation, supernatants of the cells were removed and plated in order to determine whether the addition of the agents cause the lysis of the macrophage and subsequent release of the bacterium. Adhered cells were lysed with SDS 0.01% in order to release all the intracellular bacteria and aliquots of the lysed cells were plated in 7H11 plates and incubated at 37°C. After 3 to 4 weeks, CFU were counted and bacterial concentrations calculated. The data presented are averages from three independent experiments each of which was conducted in triplicate. First published by Martins M et al. (203).
5. New concept for the therapy of MDR and XDR TB. Due to the fact that therapy of a tuberculosis infection takes place over many months, it often leads to the selection of antibiotic resistant mutants (210). Ineffective management by inexperienced physicians and patient non-compliance (211) are the primary reasons for the selection of MDR Mtb mutants. As a consequence of these factors over a prolonged period of time, the accumulation of mutations result in the development of XDR TB, an infection that is for all purpose, a lethal one, and even under the best conditions of case management (Super DOTS and same-day laboratory analyses) mortality is high (212, 213). Assuming that new drugs are created and are effective against all strains of Mtb, it will be a matter of time before resistance to the agent takes place (210).
The finding that TZ enhances the killing of intracellular Mtb strains by activating the killing machinery of the non-killing macrophage gives rise to a totally new concept of anti-TB therapy (214). Rather than target the bacterium, the targeting of the macrophage that contains the intracellular mycobacterium by-passes the mutational response expected with the targeting of the bacterium itself. The advantage of having a form of therapy that is immune to a mutational response by the bacterium, is self evident.
6. Will TZ be used for the therapy of multi-drug and extensively drug resistant infections by Mtb? The administration of 50 mg TZ in one day to a healthy human demonstrated that the QT interval was prolonged (215). This is a large initial dose and its effect on the QT interval is not surprising. However, initial therapy with TZ begun at 25 mg/day and increased slowly over time. Under these conditions, the patient soon acclimates to the drug and with a few exceptions, does not develop any notable changes in cardiac functions. TZ can be used safely and its use does not produce any side effects that differ from those produced by other non-phenothiazine neuroleptics (215). However, because approximately 6% of Eastern Europeans have a mutation in their p450 cytochrome (216), the metabolism of TZ will be slower, and with the build-up of TZ concentration, and the possibility of the heart slowing down to a level that cannot sustain life has some unknown probability (217-220). Although precautions involving cardiac monitoring prior to and during initial therapy with TZ have not been routinely taken in the past, it is reasonable that they be instituted if TZ is to be used, just to be on the safe side.
The above statements raise a question that each and every physician must pose when they have used all that is available and the XDR TB patient's prognosis of imminent mortality is certain. If precautions are taken and therapy will not harm the patient, will the use of TZ, an agent that is now proven to cure MDR TB infection in mice help the patient survive for longer, improve the quality of life, and provide a cure? Given the fact that TZ has been safely used for over 40 years and is still used in the USA and other parts of the world, what would the patient say given the choice: eventual mortality with high degree of suffering, or, try what may be the only option if quality of life is to be improved and a possible cure is achieved? This question has been recently answered by a group that demonstrated that administration of thioridazine to 12 XDR TB patients who complied with the therapy proved to be a successful treatment in 10 of the patients (224).
- Received November 5, 2009.
- Revision received April 4, 2010.
- Accepted April 14, 2010.
- Copyright © 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
-
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}