


Abstract
Muscle injury induces strong changes in muscle cells and extracellular matrix. Muscle regeneration after injury has similarities to muscle development during embryogenesis and seems to follow the same procedure. The initial phase of muscle repair is characterized by inflammation and degeneration of the damaged tissue. Almost simultaneously, previous quiescent myogenic cells, called satellite cells, are activated, proliferate, differentiate and fuse to form multinucleated myofibers. Other non-muscle stem cells may also take part in this process. Secreted factors, such as hepatocyte growth factor (HGF), fibroblast growth factors (FGFs), transforming growth factor-βs (TGF-βs), insulin-like growth factors (IGFs), tumour necrosis factor α (TNFα) and others, are released during muscle repair and guide muscle regeneration, however, their exact functions and effects on muscle remodeling remain unknown. Intensive research is currently addressing the regenerative mechanisms which are involved in acute muscle injuries and chronic muscle diseases.
- Muscle injury
- muscle satellite cells
- myogenic regulatory factors
- growth factors
- extracellular matrix
- review
One-half of the body's mass is composed of skeletal muscles. Most of these muscles are linked to bones by tendons through which forces generated during muscle contractions are transmitted to the skeleton, producing movements of the body and contributing to its stability. Therefore, impairment in muscle function could result in instability. Potential causes of impairment in skeletal muscle function include injury, disease and aging. Impaired muscle function influences the quality of life, preventing people from performing activities of daily living and being independent. Thus, factors that impact skeletal muscle structure, function and regeneration are of great importance and interest not only scientifically but also clinically (1).
Causes of Muscle Injury
As has been already mentioned, many factors can influence muscle structure. Injury to skeletal muscle may occur as a result of: (a) disease such as muscle dystrophy; (b) exposure to myotoxic agents, such as hypivacaine or lidocaine; (c) sharp or blunt trauma, such as punctures or contusions; (d) ischemia; (e) exposure to hot or cold temperatures; and (f) the muscle's own contraction.
Special attention has been paid to the cellular and molecular responses activated in exercise-induced muscle damage and regeneration (2-8). In particular, eccentric exercise, where the activated muscle is forcibly lengthened, has been used extensively as a model to study exercise-induced muscle damage, since contraction-induced muscle injury is most likely to occur during activities that involve predominantly the lengthening of muscle during its contraction (4, 9-11). As a result of eccentric contractions, the contractile system of muscle fibers sustains mechanical damage, characterized by disruption of the myofilament structures in sarcomeres, damage to sarcolemma and loss of fiber integrity (3, 7, 9, 12-14). The process of fiber regeneration appears to follow a common pathway regardless of the nature of the injurious event (1).
Skeletal Muscle Development
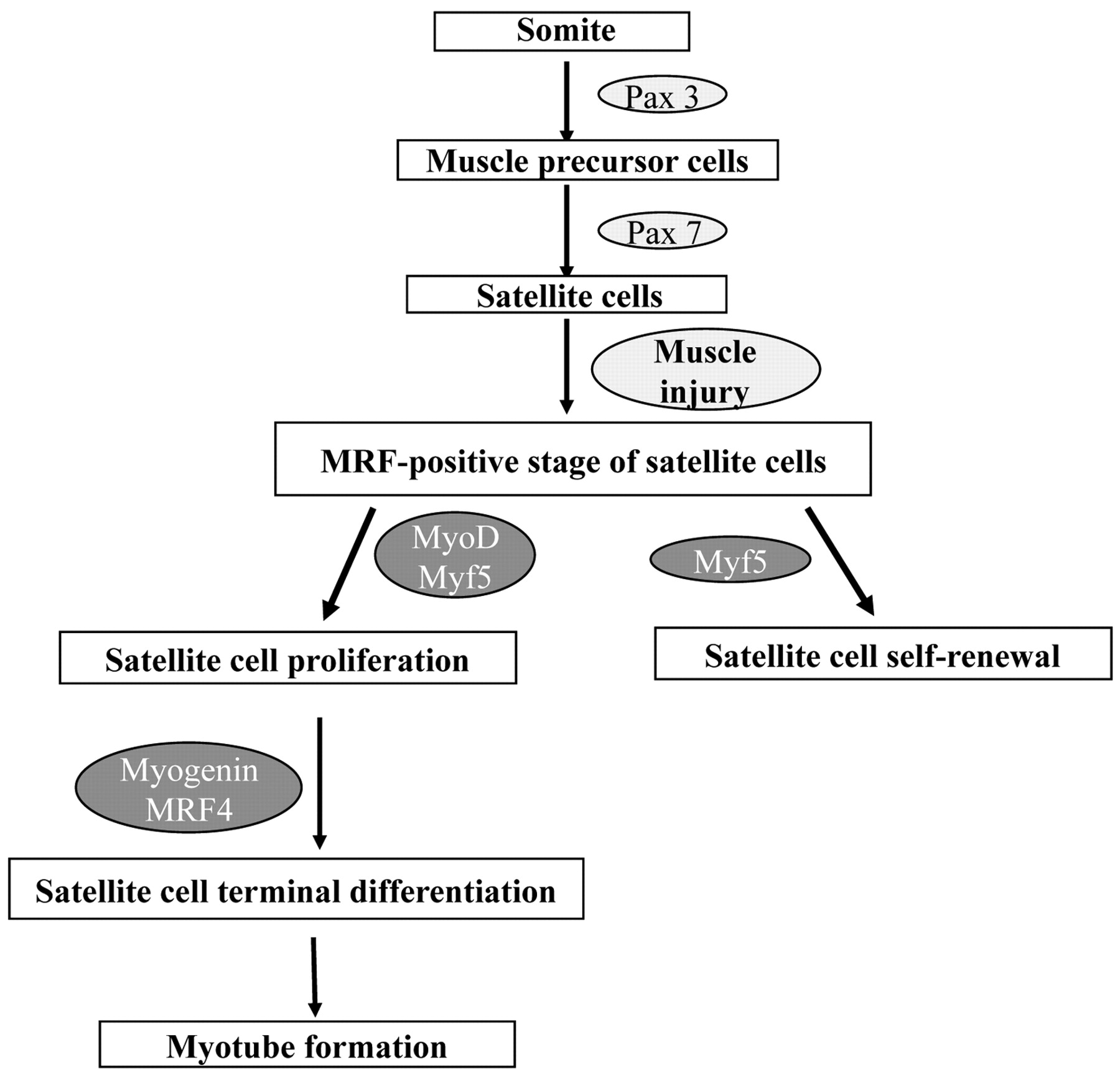
Muscle regeneration after injury has similarities to muscle development during embryogenesis. Thus, an overview of skeletal muscle development would help to understand the events during muscle repair. With the exception of head muscles, all skeletal muscles are derived from mesodermal precursor cells, which originate from the somites (epithelial spheres of paraxial mesoderm). Although the origin of muscle precursor (satellite) cells remains to be determined, it seems that the role of Pax3 and Pax7 genes for the specification of progenitor cells to the satellite cell lineage is very important (15). Pax3 and Pax7 are members of the paired box containing gene family of transcription factors and are co-expressed in the majority of myotomal cells of the somite during embryogenesis (16). Pax3 is essential for the migration of muscle precursors from the somite during development. Pax7 is possibly required for satellite cell specification (Figure 1). Mesodermal somatic cells located in the dorsal part of the somite (dermomyotome) receive signals from surrounding tissues, which induce the expression of the primary myogenic regulatory factors (MRFs), i.e. transcription factors of the myogenic lineage discussed in detail below, such as Myf5 and MyoD. The myogenic cells that express Myf5 and MyoD are called myoblasts (16). Up-regulation of the secondary MRFs (myogenin and MRF4) induces terminal differentiation of myoblasts into myocytes that now express not only myogenin and MRF4 but also important genes for muscle cells such as myosin heavy chain (MHC) and muscle creatine kinase (MCK). Eventually, mononucleated myocytes fuse to form multinucleated syncytium, which finally mature into contracting muscle fibers (Figure 1). During the late phase of embryonic myogenesis, a distinct population of myogenic precursor cells fails to differentiate and remains closely associated with myofibers as satellite cells in a quiescent undifferentiated state (8).
Skeletal Muscle Regeneration
Mammalian skeletal muscle has an impressive ability to regenerate itself, which it does on a daily basis as well as in response to injury (17). Skeletal muscle repair is a highly synchronized process involving the activation of various cellular and molecular responses, where the coordination between inflammation and regeneration is crucial for the beneficial outcome of the repair process following muscle damage (8, 18). Muscle tissue repair following damage can be considered as a process consisting of two interdependent phases: degeneration and regeneration, where, apart from the role of growth and differentiation factors, the degree of damage and the interactions between muscle and the infiltrating inflammatory cells appear to affect the successful outcome of the muscle repair process. Muscle regeneration depends on a balance between pro-inflammatory and anti-inflammatory factors that determine whether the damage will be resolved with muscle fiber replacement and reconstitution of a functional contractile apparatus, or with scar formation (18, 19). Although the phases of the repair process are similar after different causes of damage, the kinetics and amplitude of each phase may depend on the particular muscle damaged, the extent of damage, or the damage model used (2, 8, 20-22).
Degeneration. Initially, a rapid necrosis of myofibers is observed. This event is characterized by disruption of the myofiber sarcolemma and results in increased myofiber permeability. As a consequence of the disruption of the myofiber's integrity, the serum levels of muscle proteins, such as creatine kinase, are increased since these proteins are normally restricted to the myofiber cytosol. Increased serum levels of creatine kinase can be found after mechanical stress as well as during muscle degenerative diseases such as muscular dystrophies (23-25). Moreover, as the cytosolic proteins do not necessarily reflect the amount of structural damage, structurally bound proteins such as MHC and troponin have been used as better markers of damage to the contractile apparatus (26, 27). In particular, it was found that skeletal troponin I is an initial and specific plasma marker of skeletal muscle damage after exercise (26).
The early phase of muscle injury is usually accompanied by the infiltration of the damaged muscle by inflammatory cells. It should be mentioned that an important consideration is the role of the infiltrating cells, not only in mediating damage but also in the activation of repair processes necessary for successful recovery from the damage. The activation of mononucleated cells, principally inflammatory cells and myogenic cells is regulated by factors which are released by the injured muscle (24).
The first inflammatory cells to invade the injured muscle are the neutrophils. Recent reports suggest that an important increase in their number is observed 1-6 h after muscle damage (24). About 48 h after the injury, the inflammatory cells which predominate at the site of injured muscle are macrophages. These cells migrate to the damaged area through the bloodstream. Thus, in the case of significant damage of the blood supply to the injured area, regeneration cannot take place until new blood vessels penetrate the area (2). After the infiltration, macrophages phagocytose cellular debris and remove the disrupted myofilaments, other cytosolic structures and the damaged sarcolemma. Apart from that, macrophages also activate myogenic cells (28). It appears that the main histopathological characteristics of the early phase of muscle injury are muscle fiber necrosis and an increased number of non-muscle mononucleated cells within the injured area.
Regeneration. During muscle degeneration a muscle repair process is activated. An important event necessary for muscle regeneration is cell proliferation. It has been documented that new muscle fibers are formed as a result of the myogenic proliferation phase. Myogenic cells differentiate and fuse to existing damaged fibers for repair or to one another for new myofiber formation (29). Some characteristics of muscle regeneration are: (a) newly formed fibers are basophilic and this reflects a high protein synthesis. Moreover, they express embryonic forms of MHC, which means that fibers are formed de novo (30); (b) during regeneration, cell fusion is focal to the injured site rather than diffused within the muscle tissue (31); and (c) the fibers are split, which is probably due to the incomplete fusion of fibers regenerating within the same basal lamina (31, 32). When fusion of myogenic cells is completed, the size of the newly formed myofibers increases and myonuclei move to the periphery of the fiber. Finally, the new muscle tissue is the same as uninjured muscle, not only morphologically but also functionally.
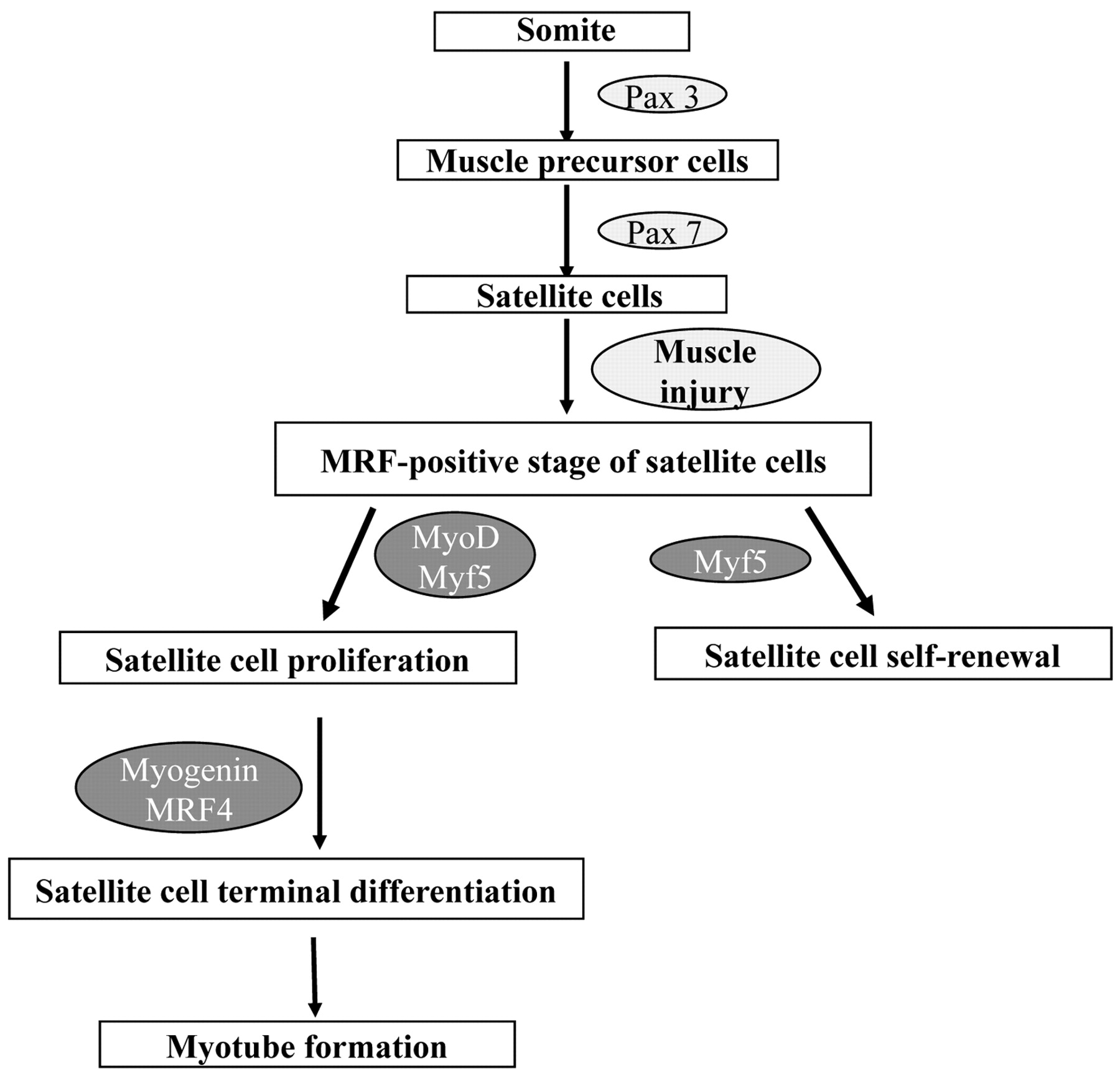
Satellite cell specification, activation and differentiation during muscle development and regeneration.
Factors Contributing to Muscle Regeneration
As skeletal muscle regeneration is a highly orchestrated process, it is clear that several factors contribute to the repair process in response to tissue injury. Muscle-specific genes, satellite cells, stem cells, trophic factors and extracellular matrix have significant roles in the reconstruction of myofibers.
Adult mammalian skeletal muscle is one of the few tissues capable of efficient regeneration after injury. This ability is due, at least partly, to a population of undifferentiated mononuclear myogenic cells named satellite cells because of their location at the periphery of mature skeletal myofibers (33). Today, it is known that satellite cells are arrested at an early stage of the myogenic program. Under unstressed conditions, they are quiescent. Following injury, or in response to increased functional demands or the need for routine maintenance, satellite cells are activated, proliferate and differentiate to give rise to myoblasts. Myoblasts then fuse with each other to form multinucleated myotubes, which give rise to adult muscle fibers. Thus, satellite cells play an important role during skeletal muscle repair after injury. Moreover, they can re-establish a residual pool of quiescent satellite cells that are able to support additional rounds of regeneration (34).
Following damage of the myofiber, quiescent satellite cells are activated to enter the cell cycle and proliferate, allowing for expansion of the myogenic cell population. After proliferation, satellite cells differentiate and contribute to the formation of new myofibers as well as to the repair of the damaged fibers. As satellite cell activation is not restricted to the damaged site, injury activates satellite cells all along the myofiber, leading to the proliferation and migration of satellite cells to the regeneration site (35). The process of satellite cell activation and differentiation during muscle regeneration is regulated by a family of muscle-specific, basic helix-loop-helix transcription factors called MRFs, including MRF4, myogenin, MyoD, and Myf5. After muscle injury, Myf5 and MyoD are typically the first MRFs to be expressed in the regenerating muscle cells, followed by myogenin, and finally MRF4 (36, 37). As a result of the exposure to signals from the damaged myofiber, quiescent satellite cells are activated and start proliferating. At this stage, the satellite cells are called myogenic precursor cells (8). The proliferative phase is followed by the differentiation and fusion of myoblasts with the damaged myofibers, for the repair of the fibers, or to each other, for new myofiber formation. Activated satellite cells are characterized by high expression of MyoD and Myf5. Hence, differentiation is accompanied by MRF expression and irreversible cell cycle exit (36, 38-41). By contrast, in quiescent satellite cells, neither MyoD nor Myf5 are expressed, suggesting that MRF expression is incompatible with reversible arrest (39, 40).
Upon satellite cell activation, MyoD up-regulation appears within a day of activation, although the complete process of regeneration to reform normal tissue architecture typically requires 3 to 4 weeks. It has been shown that some satellite cells enter the MRF-positive compartment by expressing either Myf5 or MyoD. This state is followed by coexpression of the two (42). However, it is supposed that MyoD and Myf5 play distinct roles during muscle regeneration. Although MyoD promotes satellite cell progression to terminal differentiation, Myf5 promotes satellite cell self-renewal (43, 44). As regards MyoD, it is clear that it is a key regulator of regeneration. It seems that one role of MyoD in the activated satellite cell is to directly or indirectly activate the high level of MRF4 expression found in the later stages of regeneration (36). Moreover, transcription factor Slug (Snai2) is a direct downstream target of MyoD. It has been shown that MyoD binds specifically to the Slug putative promoter region in myotubes and activates it during muscle differentiation. It is important to note that Slug expression is dramatically increased in the late phase of muscle regeneration (45). It appears that the Slug protein is required for efficient muscle regeneration; however, there are a number of possible molecular abnormalities that could give this result. It has been proposed that because Slug is a known transcription factor, a failure of myogenic differentiation found in Slug null mice may be due to downstream consequences of loss of appropriate Slug expression. Alternatively, they could also show a paucity of satellite cells (45).
The up-regulation of myogenin and MRF4 characterizes myoblast terminal differentiation. In regenerating adult muscle and in activated satellite cells, MRF4 appears after myogenin, while myogenin appears to be required for activation of MRF4 expression. The delay between myogenin and MRF4 expression in normal muscle regeneration suggests that, apart from myogenin, other factors may also be required to activate MRF4 expression in skeletal muscle. It has been shown that myogenin is associated with terminal differentiation and fusion of myogenic precursor cells to new or existing fibers. Previous studies also suggest that increased MRF4 promotes early differentiation and myotube formation. Moreover, MRF4 acts in a positive feedback loop to regulate its own expression in regenerating muscle (36, 42).
Moreover, myocyte nuclear factor (MNF) is a winged helix transcription factor that is also expressed selectively in satellite cells and acts to regulate genes which coordinate the proliferation and differentiation of satellite cells after muscle injury (46). MNF proteins are not essential for the correct patterning of skeletal muscle groups during embryonic and fetal development, nor for establishing the satellite cell population within adult muscle tissues. Rather, they appear to be required for correct temporal orchestration of molecular and cellular events necessary for muscle repair. In the absence of MNF proteins, satellite cell function is affected as regards the expression timing of myogenic determination genes and genes that control cell cycle progression. As a consequence of this dysregulation, muscle repair becomes ineffective. Specifically, in the absence of MNF, satellite cells retain an ability to proliferate in response to muscle injury, but the subsequent events that characterize the regenerative process are delayed and ineffective.
Studies have shown that there are two isoforms of MNF, MNFα and MNFβ (46). Upon satellite cell activation following muscle injury, MNFβ is down-regulated, whereas MNFα becomes more abundant. This pattern is reversed again as muscle regeneration proceeds and the quiescent satellite cell population is re-established. Because of this reciprocating pattern of expression, and because the two MNF isoforms share an identical DNA-binding domain, it is attractive to speculate that MNFα activates the same set of target genes that is repressed by MNFβ. In this way, MNF could serve to coordinate the timing of important events required for muscle repair and satellite cell renewal. In conclusion, the MNF factor is expressed selectively in quiescent satellite cells, which do not express known regulators of the myogenic program. Following muscle injury, MNF is present transiently in proliferating satellite cells and in centralized nuclei of regenerating myofibers, but its expression declines as these fibers mature, until only the residual stem cell pool continues to express detectable levels of MNF.
At the early stage of muscle regeneration, the activation of the signal transducer and activator transcription 3 (STAT3) protein is also detected first in the nuclei of activated myogenic precursor cells, and then continues to be activated in proliferating satellite cells expressing MyoD proteins. STAT3 is a protein which, in response to cytokines and growth factors including interferons (IFNs), epithelial growth factor (EGF), interleukin (IL)-5, IL6 and hepatocyte growth factor (HGF), is phosphorylated by receptor-associated kinases. They then, form homo- or heterodimers that translocate to the cell nucleus where they act as transcription activators. STAT3 mediates the expression of a variety of genes in response to cell stimuli, and so plays a key role in many cellular processes such as cell growth and apoptosis. As muscle regeneration progresses, STAT3 signaling is no longer activated in differentiated myoblasts and myotubes. It has been proposed that activated STAT3 may be an important signaling molecule that mediates essential functions for complete muscle regeneration, such as protection of activated satellite cells, proliferating myoblasts and surviving myofibers from apoptotic cell death and inhibition of myoblast differentiation at the early stage of skeletal muscle regeneration (47, 48). Nevertheless, it was recently found that another STAT3 pathway, consisting of JAK2, STAT2, and STAT3, is required for early myogenic differentiation (49). Overall, the activation of STAT3 signaling is an important molecular event that induces the successful regeneration of injured skeletal muscles.
Galectin-1, a soluble carbohydrate-binding protein with a particularly high expression in skeletal muscle, is another factor that has been implicated not only in skeletal muscle development but also in adult muscle regeneration. After muscle injury, galectin-1 immunoreactivity is increased within the cytoplasm of activated satellite cells. Thereafter, differentiated myoblasts lose galectin-1 immunoreactivity, however, galectin-1 expression associated with basement membranes is detected in myotubes (50). It has been proposed that galectin-1 is a novel factor that promotes both myoblast fusion and axonal growth following muscle injury and consequently regulates myotube growth in regenerating skeletal muscles (50, 51). It was also found that human fetal mesenchymal stem cells readily undergo muscle differentiation in response to galectin-1 through a stepwise progression similar to that which occurs during embryonic myogenesis (52).
Satellite Cells
Satellite cells are located within the basal lamina surrounding individual myofibers, between the plasma membrane of the muscle fiber and the basement membrane. In comparison to adult myofibers, they have unique morphological characteristics, including abundant cytoplasm, a small nucleus with increased amounts of heterochromatin and reduced organelle content. These features reflect the fact that satellite cells are mitotically quiescent and transcriptionally less active than myonuclei (8). Although satellite cells are present in all skeletal muscles, their percentage is different in each one of them. More satellite cells are also found close to slow muscle fibers compared with fast muscle fibers within the same muscle. Moreover their number is higher at the neuromuscular junctions (NMJ). Although these differences in satellite cell location among muscles are established, the reason for such a phenomenon is still unknown. It is well established that there is a decrease in the number of satellite cells as a function of age. The reasons why this happens are not clear. It was proposed that the self-renewal capacity of satellite cells is restricted. Thus, the exhaustion of the satellite cell pool after several rounds of regeneration may contribute to the clinical deterioration observed in the elderly or in patients with myopathies (8, 53). Muscle satellite cells appear as a distinct population of muscle precursor cells during the 10th-14th week of human limb development. As was mentioned earlier, the role of Pax3 and Pax7 genes for the specification of progenitor cells to the satellite cell lineage seems to be very important (15) (Figure 1). To date, in vivo and in vitro data demonstrate a critical role of these genes in satellite cell development, although it is not clear whether they contribute only to the specification, or to the survival of satellite cell progenitors as well (54).
Muscle satellite cells fuse to each other to form muscle fibers and so the procedure of muscle regeneration is completed. In the course of such a tissue process, intercellular junction structures which contribute to the adhesion between cells and regulate intracellular cytoskeleton architecture are required. Such molecules are M-cadherin and M-calpain. M-cadherin is a calcium-dependent intracellular adhesion molecule that is expressed in skeletal muscle cells and myoblasts (55). It plays an important role in skeletal muscle development, particularly in the fusion of myoblasts into myotubes during embryonic myogenesis and muscle regeneration. Upon muscle injury, M-cadherin expression is induced, suggesting a possible role for this protein in the muscle repair process. In addition, M-cadherin expression is up-regulated in activated satellite cells following injury (56). It remains to be determined if other cadherins play an additional role in the fusion of myoblasts during muscle regeneration. M-calpain is also required to reorganize the cytoskeletal architecture during myoblast fusion. M-calpain is a calcium-dependent intracellular nonlysosomal cysteine protease that is markedly increased during fusion of myoblasts (57). This protease produces the modification of membrane and cytoskeleton organization for myoblast fusion (58). It is worth mentioning that, in contrast with M-calpain, calpastatin, which is a specific inhibitor of M-calpain, prevents myoblast fusion (59). Further analyses are essential to determine the specific role of M-calpain in satellite cell fusion during muscle regeneration.
Skeletal muscle has the capacity for complete regeneration and repair after repeated injuries. This ability shows that the satellite cell pool is renewed after every regenerative process. To date, there are several hypotheses about how this self-renewal process might take place. Among them, the asymmetric division hypothesis is generally accepted as the most reasonable explanation of the maintenance of the satellite cell pool. According to this hypothesis, satellite cells undergo asymmetric division during mitosis resulting in one daughter cell committed to differentiation and another one that either continues to proliferate or becomes quiescent (60, 61). Numb is a plasma membrane-associated protein believed to participate in this process, as it is asymmetrically segregated during the mitosis just before differentiation (61). Numb is thought to depress the Notch-1 signaling pathway, which is known to promote the proliferation of myogenic precursor cells. These data indicate that asymmetric mitotic division and differences in Numb levels and Notch-1 activity may be responsible for satellite cell renewal. Lower levels of Numb in one daughter cell maybe permit up-regulation of the Notch-1 pathway and lead to ongoing proliferation (62). Another possibility is that satellite cell renewal is caused by different patterns of gene expression. Upon injury, activated satellite cells co-express Pax7 and MyoD. After some rounds of proliferation, some of them down-regulate Pax7 but maintain MyoD expression. These Pax7-/MyoD+ cells are committed to differentiation. The rest of them maintain Pax7 but lose MyoD expression. These Pax7+/MyoD- cells continue to proliferate slowly, or become quiescent. Thus, self-renewal might be caused by the regulation of gene expression and the withdrawal from the terminal myogenic program (54).
Myf5 is also thought to participate in satellite cell renewal. Several studies have shown that activated satellite cells first express either Myf5 or MyoD, before the co expression of these two MRFs and their progression to terminal differentiation. It is, therefore, hypothesized that the expression of Myf5 alone defines a developmental stage during which self-renewal can take place (3). Alternative mechanisms of self-renewal also include the following: (a) the population of satellite cells is heterogeneous and consists of cells ready for immediate differentiation after injury and stem cells that divide mitotically and maintain the pool (63); and (b) non-satellite cell types from muscle interstitium and bone marrow may also participate in the maintenance of the satellite cell pool, although this is not yet well established (60).
Histopathological analysis has shown that muscle satellite cells exclusively differentiate into myotubes and myofibers, and there has been no evidence that these cells are able to differentiate into non-muscle cells in vivo. However, both primary cultured mouse myoblasts and the immortalized mouse myoblastic cell line C2C12 differentiate into osteoblasts and adipocytes as well as myotubes under appropriate culture conditions (64). Specifically, it was found that satellite-derived primary myoblasts, expressing myogenic markers such as MyoD, Myf5, Pax7 and desmin, differentiate into osteocytes and adipocytes following treatment with bone morfogenetic proteins (BMPs) or adipogenic inducers (65). In contrast to myogenic differentiation, commitment of satellite cells to the osteogenic lineage is accompanied by the suppression of myogenic determination genes, including those for MyoD and myogenin. Nonmyogenic determination genes, including Runx2, are already expressed in undifferentiated myogenic cells before BMP2-induced osteogenesis, suggesting that osteogenic differentiation of muscle satellite cells might not be transdetermined (64, 66).
Stem Cells
Muscle satellite cells were once considered the only source of myogenic cells in muscle repair. Recent findings have shown that there are multipotential stem cells in various adult tissues and tissue-specific stem cells are predetermined to a specific tissue lineage. Progenitor cells isolated from bone marrow, the adult musculature, the neuronal compartment and various mesenchymal tissues can differentiate into the myogenic lineage (67-71). Specifically, bone marrow (BM) and muscle adult stem cells can differentiate into muscle cells in vitro and contribute to muscle regeneration in vivo (16).
Recently, BM-derived cells (BMDCs) have been shown to be part of several adult tissues, such as epithelia, liver, heart, brain and skeletal muscle (17). There are reports that demonstrate that BMDCs not only can give rise to muscle-specific stem cells (satellite cells) but also can fuse under physiological conditions to form mature myofibers (17). It is maintained that either all BMDCs first adopt characteristics of muscle satellite cells and then regenerate skeletal muscle fibers, or that a proportion of these cells fuses directly with myofibers and that both mechanisms coexist in the same tissue. Specifically, it has been found that BM-derived multipotent adult progenitors (Cd13+/Scal-/Flk-/Cd45-/Cd31-) are capable of forming differentiated myotubes in vitro and in vivo following delivery into murine muscle (16). It has been also demonstrated that the green fluorescent protein (GFP)-labeled BMDCs become satellite cells that are capable of myogenesis (16).
It is now well accepted that a single hematopoietic stem cell (HSC) can give rise to cells that reconstitute all of the lineages of the blood as well as contribute to mature muscle fibers (16). It has been shown by single-cell transplantation experiments that HSCs can give rise to progeny that reconstitute the blood and integrate into regenerating myofibers. In addition to this, HSC derivatives that have the capacity to regenerate muscle fibers exist in the pool of hematopoietic cells known as myelomonocytic progenitors. By contrast, the results of Shi's study indicate that mature progeny of myelomonocytic progenitors (macrophages) do not fuse spontaneously with myofibers and do not contribute to muscle regeneration (16). Moreover, it has been shown that neural stem cells, which generate neurons, glia and blood cells, can also produce myotubes and undergo various patterns of differentiation depending on their exposure to appropriate epigenetic signals in mature tissues, including skeletal muscle (68). These facts suggest an important role of non-muscle stem cells in skeletal muscle repair. Nonetheless, further studies are necessary to establish the optimal cellular and environmental conditions that promote myogenic conversion of non-muscle stem cells for therapy.
Like BMDCs, a great population of adult stem cells is capable of myogenesis and the formation of myofibers. Muscle side population (mSP) cells constitute a stem cell/progenitor cell population that resides in adult tissues including BM and skeletal muscle. mSP cells contribute to myogenesis both in vitro and in vivo (72, 73). Previous studies have established that mSP cells increase in number following muscle injury, have a distinct molecular signature and participate in muscle regeneration (16). Furthermore, mSP cells may represent satellite cells progenitors (72, 73). The in vitro conversion of mSP cells to the myogenic lineage requires the cooperation of myoblasts (73). But there is evidence that there are progenitor cells with myogenic potential other than satellite cells within skeletal muscle. However, mSPs represent a cellular population separate from satellite cells; they may represent satellite progenitors and myogenic progenitors capable of direct myogenic fusion (72). It is important to characterize muscle-derived stem cells taking into consideration the cell surface markers, in order to have more information about cellular characteristics and origin. Thus, it is of much interest that most of the mSP cells express Sca-1 in contrast to a small percentage which expresses CD45 consisting of both Sca-1+ and Sca-1-. Although both CD45+ and CD45- mSP cells contribute to myogenesis, myogenic potential appears to be greater in CD45- than CD45+ muscle-derived cells (73, 74). An additional stem cell population that has been isolated from adult skeletal muscle is that of muscle-derived stem cells (MDSCs) which are CD34+/Sca1+). After muscle regeneration involving vascular damage, CD34+/Sca1+ cells migrate from blood vessels to become part of regenerating myofibers. These data do not clarify, however, if the CD34+/Sca-1+ cells represent progenitors of satellite cells (16, 75). Further studies would be necessary to define the relationship of these stem cell/progenitor cell populations to satellite cells and their capacity to participate in the growth, maintenance and regeneration of skeletal muscle in response to injury and disease.
The Role of Secreted Factors in Muscle Regeneration
Skeletal muscle repair is a highly orchestrated process that involves the activation of quiescent satellite cells to proliferate and differentiate. This activation requires the controlled up-regulation of muscle transcription factors and muscle-specific genes. The role of various secreted growth factors has been shown to be very important during this process (Figure 2). Muscle injuries cause the release of biologically active molecules into the extracellular space. These molecules may be endogenous to the injured tissue itself or may be synthesized and secreted by other cell types at the wound site, including neutrophils and macrophages. All these factors are thought to play a role in the different stages of muscle regeneration.
Hepatocyte growth factor (HGF). HGF or scatter factor is a protein which is bound to the extracellular matrix in muscle tissue and is released in response to injury. HGF is believed to play a fundamental role during muscle regeneration. Its mitogenic, motogenic and morphogenic activities, especially during the initial phase of muscle repair, are considered to be essential for effective muscle regeneration (76). HGF is believed to promote the quiescent satellite cells to enter the cell cycle (77). There are two theories about how this action is mediated. It has been shown that HGF activates p38 mitogen-activated protein kinase (MAPK) and phosphatidylinositol 3-kinase (P13K) signaling pathways (16). Alternatively, HGF down-regulates caveolin-1 protein expression. This leads to the up-regulation of the ERK pathway, which is required for the satellite cell activation (78).
It has also been proposed that HGF inhibits the differentiation of myogenic precursor cells (76), probably by regulating Twist protein and p27. Twist is an inhibitor of MyoD expression and, therefore, of the progression to terminal differentiation. P27 is a cyclin-dependent kinase inhibitor. Twist and p27 are coordinately regulated. As satellite cells proliferate, p27 levels slowly increase to a threshold level. Cells then exit the cell cycle, turn off Twist expression and undergo differentiation. HGF induces Twist and reduces p27 levels and both these regulations are required for the inhibitory activity of HGF on cell differentiation (77). The exact effects on gene expression leading to this inhibitory activity in vivo are not yet well understood. Nevertheless, in vitro studies suggest that the binding of HGF receptor, c-met, results in the silencing of MyoD and myogenin gene expression and inhibits the synthesis of muscle-specific structural proteins (e.g. MHC) as well as myotube formation (79). Finally it should be mentioned that HGF could stimulate satellite cells and myogenic precursor cells migration to the site of injury. The activation of either Ras-Ral or ERK pathway mediates this action (8).

The role of secreted factors and extracellular matrix in muscle regeneration.
There are several mechanisms through which HGF acts on satellite cells. HGF is produced and released by regenerative myotubes (80). Upon injury, it is also released from the ECM by an NO-dependent mechanism (16). Moreover, soon after injury its expression is up-regulated in the spleen (81). These data indicate that the actions of HGF during muscle repair may be a result of autocrine, paracrine, or endocrine mechanisms. HGF transduces its signals in the cells via c-met. C-met is a tyrosine kinase receptor expressed broadly in the quiescent satellite cells of skeletal muscle. Its presence is very important for both muscle development during embryogenesis and muscle repair after injury, underlying the important role of HGF and c-met during these two processes (79).
Fibroblast growth factor (FGF). Fibroblast growth factor-6 (FGF-6) belongs to a family of cytokines that control cell proliferation, cell differentiation and morphogenic events. Several members of the FGF family are expressed in developing skeletal muscle, but FGF-6 is the only one that seems to participate in muscle repair (82, 83). The high amounts of FGFs, which are released within the short period of inflammation after tissue disruption, may induce satellite cells to proliferate and partly cause the chemotaxis of further muscle precursor cells (84). Today, it is known that the expression of FGF-6 is stimulated after skeletal muscle injury (16). FGF-6 induces strong morphological changes, alters satellite cell adhesion and compromises their ability to differentiate into myotubes (85). Moreover, like other FGFs, FGF-6 is thought to possess angiogenic activity. On the other hand, FGF-6(-/-) mutant mice show increased fibrosis and myotube degeneration after injury (82). In conclusion, these data suggest that FGF-6 stimulates the proliferation of satellite cells and induces the expression of genes required for their terminal differentiation (e.g. MyoD and myogenin) (82, 83).
However, it should be noticed that other FGFs are potential redundant factors to FGF-6 in vivo (86). For instance, FGF-1 and -2 have been shown to stimulate proliferation and repress differentiation of myogenic precursor cells. The high level of FGF-1 related to the increasing rate of myogenic precursor cell proliferation suggests a possible intracrine role for this factor in the early phase of muscle regeneration (84). FGF-2 is also thought to regulate satellite cells activity in regenerating muscle. In vitro, FGF-2 can induce satellite cells to enter the cell cycle, although it cannot accelerate their transit from proliferation to differentiation (83). Moreover, its mitogenic influence is limited to the initial days after injury. Thus, the role of FGF-2 in muscle repair seems to be important but not yet well understood. The FGFs transmit their signals to the cells through transmembrane tyrosine kinase receptors (FGFRs), for which four distinct genes have been discovered (FGFR 1-4). FGFR-1 and 4 are the most prominent transcripts in satellite cells. The expression of the correlative genes is up-regulated during muscle repair. To date, there is evidence that FGFR-1 may regulate ongoing proliferation of satellite cells, whereas FGFR-4 is possibly involved in their differentiation (83).
FGFs are thought to have neurotrophic activity. FGF-2 can interact with other growth factors to stimulate the synthesis and secretion of nerve growth factor (NGF). Moreover, FGF-5 is a muscle-derived survival factor for cultured spinal motoneurons, which might be important for the reinnervation process in generating muscle. FGF-6 is also expressed in adult muscles and during embryogenesis. FGF-7 regulates the division and differentiation of myoblasts in the developing myotubes and is active during the early development of limb musculature (84). In conclusion, the main action of FGFs is the continuous activation of muscle precursor cell proliferation, which provides enough cells to allow regeneration to occur (84-86). FGFs might induce satellite cells to proliferate, possibly through an indirect differentiation inhibiting mechanism, involving insulin-like growth factor II (IGF-II). They might also repress differentiation by negatively regulating the expression of the myogenic determination gene MyoD and by suppressing the activation of the muscle gene program, even in the presence of constitutive expression of myogenic determination genes (84). Furthermore, it is likely that FGF-6 acts by stimulating the entry of satellite cells in the cell cycle rather than by directly promoting MyoD expression.
Transforming growth factor-beta (TGFβ). The TGFβs are a small family of multifunctional growth factors, consisting of TGFβ1, β2 and β3. Recently, myostatin was also identified as a member of the TGFβ family. A common characteristic of all TGFβs is their ability to bind to extracellular proteins and to be stored in the ECM until activation by a physiological process, such as wound healing (87). Despite the similarities of their actions in vitro, it seems that each of the TGFβs has a different and discrete role in vivo (88). After injury, TGFβ is released by degranulating platelets at the site of injury and autoinduces its own production by resident cells, such as smooth myogenic cells. It has been shown that TGFβ3 is synthesized by myoblasts and inhibits myogenic differentiation (84). Following the denervation and the ischemia of skeletal muscles, TGFβ1 and TGFβ3 are expressed by regenerating muscles within the first days after a trauma (84)
In vitro, TGFβs can depress satellite cell proliferation and differentiation in a dose-dependent manner (89). This might prevent the activated satellite cells from leaving the proliferative state and so continue their replication (84). However, it seems that TGFβs can inhibit, induce or have no effect on satellite cells proliferation in vivo, depending on whether platelet-derived growth factor (PDGF), FGF or IGFs are present or not (88, 90). TGFβs are also believed to regulate the immune response, as well as motoneuron survival after injury (88). Indeed, it is chemotactic not only to macrophages but also to leukocytes. It stimulates the synthesis of fibronectin, collagens, proteoglycans and novel matrix proteins and it induces angiogenesis. TGFβ is also chemotactic and activating for monocytes, which are potent inducers of angiogenesis and secrete FGF, tumor necrosis factor (TNF) and IL-1 (84). TGFβ2 regulates where and when myoblasts fuse to form myotubes and TGFβ1 promotes fibroblasts to differentiate and may inhibit myogenic development, regulating the formation of epimysium and perimysium (88). Apart from this, it appears that TGFβs have an influence on ECM reorganization throughout the regeneration period of muscle. It seems to be responsible for the reconstruction of the basement membrane and ECM that surrounds the damaged myofibers and the activated satellite cells. TGFβs also stimulate the production of PDGF. Finally, excessive TGFβ-induced deposition of ECM at the site of injury can lead to fibrosis (84).
Myostatin (MSTN). MSTN, a member of the TGFβ superfamily, plays an important role in regulating skeletal muscle growth. MSTN(-/-) mutant mice have a dramatic and widespread increase in skeletal muscle mass (91). Skeletal muscle development and regeneration are highly paralleled processes under balanced regulation. MSTN is well positioned as a negative regulator of these processes (92). It is specifically expressed during embryogenesis, as well as in adult skeletal muscles during regeneration. High levels of MSTN are detected within necrotic fibers and connective tissue during the degenerative phase of muscle repair. On the contrary, regenerative myotubes contain no MSTN during enlargement and fusion (93). This expression profile suggests that MSTN acts like an inhibitor of muscle growth, perhaps via repressing satellite cells proliferation during muscle regeneration. It has been shown that MSTN inhibits cell proliferation and protein synthesis in C2C12 muscle cells (94), as well as the expression of MyoD and Pax3 (95), while it stimulates the expression of genes that take part in ubiquitin-mediated proteolysis. These in vitro data are also consisted with MSTN being a negative regulator of muscle growth and hypertrophy (96). Moreover, MSTN function was implicated in both hypertrophy and hyperplasia of muscle, since mice completely lacking MSTN showed a profound increase in skeletal muscle growth with both an increase in myofiber size and an increase in myofiber number (97). The question of how MSTN negatively regulates myogenesis has not been answered. A hypothesis is that MSTN may regulate fiber size and number during skeletal muscle development. MSTN is produced and secreted by muscle cells and therefore it may be autocrinically or paracrinically involved in the control of myogenic cells proliferation and differentiation (16).
Insulin-like growth factor (IGF). It is very clear that the growth hormone (GH)-IGF axis plays a major role in controlling the growth and differentiation of skeletal muscle, as it does virtually in every tissue. Recently, the activity of this axis during muscle repair has become more apparent. GH acts directly on skeletal muscle during regeneration. Although the presence of GH receptor is established, most studies have failed to demonstrate direct binding of GH to muscle cells (90). On the contrary, the role of IGFs in muscle repair is evident. IGF-I is known to induce muscle hypertrophy by increasing myotube size, and DNA (98) and protein synthesis (99, 100). It also causes biochemical changes, including both activation of enzymes and increase of products of anaerobic glycolysis (99). Moreover, IGF-I can promote muscle growth by inhibiting protein degradation and suppressing the expression of two muscle-specific atrophy-related ligases: muscle ring-finger 1 (MuRF1) and atrogin-1 (also known as muscle atrophy F-box (MAFb) (101, 102). Thus, IGF-I prevents loss of muscle mass and contributes to an increase of muscle strength (98, 100).
During muscle regeneration, IGF-I is unique among growth factors, as it stimulates both proliferation and differentiation of muscle cells (34, 90, 103). IGF-I increases the proliferation potential of satellite cells (104) by enhancing the expression of intracellular mediators, such as cyclin-D (104-106). It also stimulates terminal differentiation by inducing myogenin gene expression (90, 106-107). It is very interesting that in the time course of muscle regeneration, IGF-I initially reduces the expression of myogenic factors and induces cell proliferation as mentioned. This pattern is later reversed, when IGF-I increases myogenin gene expression and down-regulates cell-cycle markers. Thus, IGF-I first enhances proliferation and subsequently induces differentiation of muscle satellite cells (103). Moreover, an attractive aspect has been developed in the literature about the differential expression and implication of IGF-I isoforms [IGF-IEa, IGF-IEb and IGF-IEc (also known as mechano growth factor, MGF)] in the regulation of muscle fiber regeneration (106, 108). In particular, it was found that IGF-IEa and IGF-IEc isoforms have different expression kinetics and they probably act as different growth factors, with apparently different function. IGF-IEc is rapidly activated and subsequently depressed in damaged muscles, and it was suggested that this isoform is responsible for satellite cell activation, prolongation of myogenic cell proliferation and for depression of their terminal differentiation into myotubes (109, 110). In contrast, IGF-I Ea appeared to have a more delayed expression profile, increase the mitotic index, enhance terminal differentiation and to promote fusion of the myogenic cells. However, both, these isoforms appear to up-regulate muscle protein synthesis (109, 110).
Moreover, IGF-I promotes muscle cell survival during the initial phase of their differentiation (111). It has been proposed that IGF-I can promote nerve sprouting and expression of nerve growth supporting molecules by activated interstitial cells (112). More recently, it has been shown to reduce fibrosis and modulate the inflammatory response following injury, by down-regulating pro-inflammatory cytokines (100). In this way IGF-I contributes to efficient muscle regeneration. The role of IGF-II in muscle repair has not been so well investigated. It is known that the expression of IGF-I and -II occurs at different stages of muscle regeneration, with IGF-I expression preceeding that of IGF-II (113). Moreover, myoblasts are shown to express autocrine IGF-II after a period of time (90). Thus, it seems that IGF-I and -II have different effects on gene expression and IGF-II appears to be more related to the formation of myotubes (114).
Tumor necrosis factor-α (TNFα). As previously mentioned, inflammation is a key response to muscle injury and is essential for muscle regeneration. It has long been known that TNFα, which is produced by activated leucokytes, plays several important roles in inflammation, including activation and chemotaxis of leucokytes, expression of adhesion molecules and regulation of the secretion of other pro-inflammatory cytokines (115, 116). More recently, it has been proposed that TNFα contributes to the degenerative and regenerative processes after muscle injury. TNFα seems to participate in the muscle protein loss during the degenerative phase of muscle regeneration. It has been shown that TNFα promotes the activation of nuclear factor κB (NFκB) in skeletal muscle cells. NFκB is a transcription factor, which activation alters gene expression and causes proteolysis.
In vitro and in vivo data indicate that TNFα promotes the expression of atrogin-1, leading to the catabolism of muscle proteins. This is a result of the activation of the ubiquitin/proteasome pathway in muscle fibers and is believed to be mediated via the p38 MAPK signaling pathway (102, 117). Moreover, TNF-binding to TNFR-1 and the induction of reactive oxygen species production seem to play an important role during this process (118). Equally important is the role of TNFα during the regeneration phase. After skeletal muscle injury TNFα is released not only by infiltrating macrophages, but also by injured muscle fibers (115). Its expression remains at high levels during the repair process and returns to normal levels several days post-injury (116). This expression profile, along with in vitro studies, suggests that TNFα has a double role during muscle regeneration: it is both a competence factor (activating satellite cells to enter the cell cycle) and a progression factor (enhancing satellite cell proliferation, once it has been initiated, perhaps via activating the expression of c-fos gene, well known as an important cell growth regulator) (117). TNFα may also take part in satellite cell terminal differentiation, as in vitro studies indicate that TNFα deficiency depresses MyoD expression, although it does not alter myogenin and MRF4 expression (116). However, its absence only moderately affects the muscle repair process, which would indicate that redundant mechanisms or factors might exist in vivo (115, 116).
Interleukin-6 (IL-6). IL-6 is a cytokine with a major role in the regulation of the inflammation process after muscle injury (119). The time course of IL-6 expression after muscle injury is similar to that of TNFα in vitro (116). Some in vitro studies show that IL-6 has trophic effects and may take part in muscle repair after injury. Other studies show that it regulates muscle protein degradation (120). These contradictory in vitro data indicate that in vivo IL-6 has a multiple role in muscle regeneration: it can be both a factor that induces proteolysis of damaged myofibers and a proliferation signal for satellite cells to replace the destroyed muscle tissue. To date studies suggest that skeletal muscle can produce IL-6 in response to exercise as well as inflammation and injury (121, 122). IL-1β is a possible regulator of IL-6 production in myotubes, probably by activating the MAPK signaling pathway and NFκB (120). Epinephrine (123) and TNFα (122) may also contribute to this regulation.
Leukemia inhibitor factor (LIF). Recent data suggest that LIF is produced by the regenerating muscle itself (122), as well as by other cells in culture (121) and seems to play a pleiotropic role during muscle regeneration (124). Its expression is regulated by other cytokines and growth factors such as IL-1α, IL-1β, TNF, FGF-2 and members of the TGFβ family. LIF acts via binding to specific membrane receptors (84). It has been shown to enhance the proliferation of myogenic precursor cells in vitro and to increase their number and size in vivo (125-127), but does not suppress myoblast fusion to myotubes (125, 127). LIF also seems to affect the amount of fibronectin, tenanscin-c, collagen type IV and laminin produced by the fusing myoblasts and activated fibroblasts. This suggests a major role for LIF, as the ECM remodeling is a basic aspect of muscle repair (127). Finally, it has been shown that following nerve injury, LIF expression increases rapidly at the site of injury and promotes both motor and sensory neuron survival (128, 129).
Nerve growth factor (NGF). NGF is essential for sympathetic neurons and neural-crest derived primary sensory neurons. Furthermore NGF is present in very low amounts in skeletal muscle (84).
Platelet-derived growth factor (PDGF). PDGF was identified as a growth-promoting activator in human platelets. Three isoforms exist: PDGF-AA, -AB and -BB. In skeletal muscle regeneration, only the BB isoform of PDGF has a significant effect. Myoblasts bind selectively to PDGF-BB isoforms provoking a mitogenic response. After PDGF is released from injured vessels, platelets and macrophages, it stimulates angiogenesis in vivo. It also causes cell migration, including that of adult muscle precursor cells. It is known that PDGF is first released from degranulating platelets and later by infiltrating activated macrophages. Therefore, platelet breakdown products stimulate muscle regeneration (84). To sum up, PDGF not only stimulates the proliferation of satellite cells but also inhibits their differentiation, while it is also chemotactic for them (8, 16, 84).
Extracellular Matrix Remodeling
For many years, the ECM was believed to play a passive role during muscle development and regeneration, acting simply as a scaffold for the arrangement of the cells within tissues. Today, it is believed that proteoglycans, collagen, metalloproteinases and other elements of the ECM might play an energetic key role during muscle development (130-132). Although the exact function of these other elements in muscle regeneration is not well understood, the similarities between skeletal muscle embryogenesis and regeneration might indicate a similar role of the ECM during these two processes. Satellite cells are surrounded by ECM components and remodeling of these ECM molecules has been seen in many myoblast and muscle cell responses such as migration, fusion and myotube maturation (133-135). Moreover, the ECM acts as a reservoir of growth factors. For instance, TGFβ binds to decorin, a heparan sulfate proteoglycan, and is stored in the ECM. Matrix degeneration after injury can release these growth factors, which can then transduce their signals to the cells and regulate the regeneration process (33, 136). The ECM may also participate in the regulation of the activity of various growth factors. Molecular studies have proven that FGF and TGFβ can bind to extracellular proteoglycans (87). FGF-binding to syndecan-1 and glypican seems to be essential for the function of FGF during muscle development and repair, while the binding of TGF to decorin inactivates TGF (87, 132). Therefore, the ECM may be involved in the precise regulation of the activity of growth factors during muscle regeneration (Figure 2). Moreover, it is notable that the ECM possibly participates in satellite cell migration to the site of injury (132, 137). Cell migration involves a series of complex cell ECM interactions, including adhesion to ECM through integrins, contraction of the cytoskeleton, translocation and release of cell-ECM contact (132). The right function of integrins is essential for the migration process and may also have a critical role during muscle repair. Furthermore, various studies suggest that interactions between neuron axon and Schwann cells as well as between Schwann cells and the ECM are essential for nerve growth during peripheral nerve regeneration. This suggests a possible pole for ECM in the reinnervation of injured skeletal muscle (138, 139). Besides, ECM may also participate in signaling pathways essential for muscle differentiation, as myogenin alone cannot stimulate cell terminal differentiation (130).
Another important element of the ECM is the family of metalloproteinases (MMPs) and their natural antagonists, namely the tissue inhibitors of metalloproteinases (TIMPs). Matrix physiological remodeling requires the action of proteinases, among which MMPs appear to be major players, since the turnover of ECM is mediated via MMPs while inhibited by specific TIMPs (140). MMPs are a family of Ca- and Zn-dependent endopeptidases able to cleave most of the ECM components (138). Their expression is highly regulated during both muscle development and repair (132, 141), and it appears that, in such conditions, MMPs and plasminogen activators act in concert. Thus, plasmin activity together with MMPs is needed to complete a wound healing process; urokinase-type plasminogen activator (uPA)-uPAR complex participates in fibrinolysis while MMPs have the capacity to split fibrin by acting as pericellular fibrinolysins (142, 143). Moreover, some MMPs, together with plasmin and uPA, can activate several latent growth factors and proteases, such as TGFβ1 and basic fibroblast growth factor (bFGF), whose activities are purported to be crucial for cell migration and tissue remodeling process in vivo and in vitro (144-146). To date, data indicate that MMPs are involved in myoblast migration to the site of injury (138). The role of the TGFβ1/uPA bioregulation system has been implicated in several pathophysiological processes (147-159) and increasing evidence particularly supports an important role of uPA in promoting invasiveness, fibrinolysis and matrix remodeling in various physiological and pathological processes other than muscle regeneration, such as in optimizing the survival of metastatic cancer cells (136, 158, 160-163).
uPA and plasmin are implicated in several non-fibrinolytic processes, which lead to ECM degradation, either directly by proteolytic cleavage of ECM components, or indirectly through the activation of latent MMPs (37, 164, 165). Indeed, a proteolytic activation cascade initiated by uPA/plasmin is involved in MMP activation during muscle regeneration (37, 146), since MMPs are secreted in latent form and need to be converted to active form to have proteolytic activity (140). Plasmin can directly activate several MMPs in vivo through proteolysis and it appears that the activation of MMP-2 and MMP-9 during skeletal muscle regeneration could be mediated by plasmin (37, 146). These metalloproteinases appear to be differentially expressed at different stages of the degeneration and regeneration processes of experimentally damaged skeletal muscle (166). It has been proposed that MMP-9 expression is related to the inflammatory response and probably to the activation of satellite cells, since its expression is induced within 24 hours post damage and remains present for several days, while MMP-2 activation is concomitant with the regeneration of new myofibers (166).
Moreover, activated satellite cells can synthesize and secrete MMP-2 and 9 and may be involved in the ECM remodeling after injury (133, 138, 166). Other MMPs, such as MMP-7, are thought to take part in the fusion of myoblasts (138). Furthermore, MMPs are possibly involved in the transmission of growth factor signals during muscle repair (87, 130, 132, 138). To conclude, these data are suggestive of a multiple role, although not precisely clarified, of MMPs in muscle regeneration.
Furthermore, since ECM remodeling is a result of the balance between synthesis and degradation, it follows that an excess of TIMPs does not favour proteolysis, as TIMPs bind to MMPs and regulate their enzymatic activities. TIMP-1 and TIMP-2, inhibitors of MMP-2 and -9, were found to be up-regulated during the degeneration and regeneration phases respectively, following muscle damage (167). This result is augmented by the increased levels of circulating MMP-9 as well as TIMP-1 and TIMP-2 found following muscle damage, providing evidence of ECM remodeling possibly via the MMP/TIMP pathways following muscle-damaging exercise (168, 169).
Finally, collagen is one of the most basic components of the ECM. Its production upon injury should be carefully regulated in order to achieve muscle repair and to avoid excessive cicatrization. Type I collagen is the basic collagen type in normal skeletal muscle. Soon after injury, fibroblasts synthesize collagen type III. Almost a week post-injury, the production of type I collagen is increased and the type III/type I ratio decreases below normal. However, as the regeneration process proceeds, the type III/type I ratio returns slowly to normal. It is not yet understood why this reversion of the collagen type production is observed during the initial phase of muscle repair (170).
The Role of Degenerative Fiber Nuclei in Muscle Regeneration
Muscle regeneration has long been thought to be due to satellite cell activation. The effects of several growth factors and ECM components are being intensely investigated. However, the radical idea that post-mitotic myonuclei can possibly dedifferentiate after injury and reform myoblasts or stem cells is rather attractive, as the damaged myofibers themselves could then provide the essential molecules for muscle repair. It has long been known that cells of adult urodele amphibians have the ability to re enter the cell cycle after injury. On the contrary, mammalian cells were thought incapable of dedifferentiation because two processes must take place in order for this to be possible: formation of new membranes around myonuclei and induction of DNA synthesis. Neither of these has ever been observed until recently (171, 172). Electron microscope observations have indicated that the formation of new membranes around myonuclei in injured mouse myofibers is possible (172). Moreover, mammalian myotubes can dedifferentiate when treated with an extract from newt regenerating limbs (171). Msx1, a transcriptional factor, can also induce mouse myotubes to dedifferentiate to cells capable of re-differentiating into various cell lineages. Myoseverin, a microtubule-binding molecule, can cause myoblasts to be generated from mouse myotubes in the C2C12 cell line (8).
These studies suggest that the molecular pathways and mechanisms required for dedifferentiation are present in mammalian cells. Therefore, it is likely that their inability to undergo dedifferentiation in vivo is due to the lack of such essential signals. If this hypothesis is correct, the identification of factors inducing dedifferentiation may lead to a new approach to the regeneration process (171).
Conclusion
In conclusion, the results of intensive studies to date indicate that muscle regeneration is a far more complex process than was initially believed. The various cell populations involved, the precise regulation of gene expression, the multifunctional role of the known and unknown growth factors and connective tissue components suggest that a new field of research is opening up. Although studies concerning muscle regeneration after injury have been performed mainly in animal models, these models provide a way to an understanding of the cellular and molecular signaling pathways involved in muscle degeneration and regeneration and, hence, could potentially lead to clinical interventions and cell-based therapies. The use of such models could hold great promise in treating age-related muscle atrophy or congenital and acquired myopathies, such as Duchenne dystrophy, which are common and, as yet, untreatable and fatal. Thus, future studies are expected to further define the molecular pathways and interactions that are essential for effective muscle regeneration, which would contribute to the development of new therapies in humans.
- Received April 6, 2009.
- Revision received July 7, 2009.
- Accepted July 14, 2009.
- Copyright © 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
Jump to section
- Article
- Abstract
- Causes of Muscle Injury
- Skeletal Muscle Development
- Skeletal Muscle Regeneration
- Factors Contributing to Muscle Regeneration
- Satellite Cells
- Stem Cells
- The Role of Secreted Factors in Muscle Regeneration
- Extracellular Matrix Remodeling
- The Role of Degenerative Fiber Nuclei in Muscle Regeneration
- Conclusion
- References
- Figures & Data
- Info & Metrics
Related Articles
Cited By...
- Study of Therapeutic Effects of Losartan for Sarcopenia Based on the F344xBN Rat Aging Model
- RNASeq analysis of a Pax3-expressing myoblast clone in-vitro and effect of culture surface stiffness on differentiation
- Myofiber injury induces capillary disruption and regeneration of disorganized microvascular networks
- Characterization of Optimal Strain, Frequency and Duration of Mechanical Loading on Skeletal Myotubes' Biological Responses
- Deficient Skeletal Muscle Regeneration after Injury Induced by a Clostridium perfringens Strain Associated with Gas Gangrene
- Deficient skeletal muscle regeneration after injury induced by a Clostridium perfringens strain associated with gas gangrene
- Krüppel like factor 2 - deficient myeloid cells promote skeletal muscle regeneration after injury
- Skeletal Muscle Volume and Intramuscular Adipose Tissue Are Prognostic Predictors of Postoperative Complications After Hepatic Resection
- TGF-{beta} Family Signaling in Mesenchymal Differentiation
- Diet-induced obesity impairs muscle satellite cell activation and muscle repair through alterations in hepatocyte growth factor signaling
- Role of Phosphoinositide 3-OH Kinase p110{beta} in Skeletal Myogenesis
- Regulatory mechanisms of metabolic flexibility in the dark-eyed junco (Junco hyemalis)
- A Novel Isoform of Met Receptor Tyrosine Kinase Blocks Hepatocyte Growth Factor/Met Signaling and Stimulates Skeletal Muscle Cell Differentiation