


Abstract
The aim of this study was to investigate whether a total of twenty benzo[b]cyclohept[e][1,4]oxazines and their S-analogs, and 2-aminotropone derivatives affect the function of activated macrophages. These compounds inhibited the production of pro-inflammatory substances such as nitric oxide (NO) by lipopolysaccharide (LPS)-activated mouse macrophage-like RAW264.7 cells to different extents. Among them, benzo[b]cyclohept[e][1,4]oxazin-6(11H)-one [5] and 7-bromo-2-(4-hydroxyanilino)tropone [16] showed the highest inhibitory effects at concentrations that did not affect cellular viability (selectivity index=74.89 and 54.15, respectively). Western blot and RT-PCR analyses showed that [16] inhibited the expression of both inducible NO synthase (iNOS) and cyclooxygenase (COX)-2 at both protein and mRNA levels, whereas [5] inhibited only iNOS protein expression. Electron-spin resonance (ESR) spectroscopy revealed that both [5] and [16] scavenged nitric oxide (generated from NOC-7) and superoxide anion (generated by HX-XOD reaction) only at much higher concentration. These data suggest that [16] but not [5] exerts its anti-inflammatory action against macrophages via the inhibition of iNOS and COX-2 protein expressions.
- Benzo[b]cyclohept[e][1,4]oxazines
- 2-aminotropones
- RAW264.7 cells
- macrophage
- NO
- COX-2
- anti-inflammatory activity
Heterocylic compounds are known to display diverse biological activities (1). Hinokitiol and its related derivatives with a tropolone skeleton (2-4) have been reported to exhibit various biological activities such as antimicrobial (5), antifungal (6) and phytogrowth-inhibitory activity (7, 8), cytotoxic effects on mammalian tumor cells (9, 10), and inhibitory effects on catechol-O-methyltransferase (11) and metalloproteases (5). Hinokitiol acetate did not exhibit cytotoxic activity (10), antimicrobial activity nor metalloprotease inhibition (5), suggesting that these biological effects of hinokitiol-related compounds may result from the formation of metal chelates by the carbonyl group at C-1 with the hydroxyl group at C-2 in the tropone skeleton.
A total of 173 azulene, tropolone and azulenequinone-related compounds have been investigated for their ability to induce tumor-specific cytotoxicity and to inhibit nitric oxide (NO) and prostaglandin (PG) E2 production by lipopolysaccharide (LPS)-stimulated mouse macrophage-like RAW264.7 cells (12-25). We have previously reported that tropolone, 2,4-dibromo-7-methoxytropone (14), diethyl 2-chloroazulene-1,3-dicarboxylate, 1,3-difluoroazulene (12), 3,7-dibromo-1,5-azulenequinone and 3-morpholino-1,5-azulenequinone (16) potently inhibited NO production by LPS-activated RAW264.7 cells. However, the extent of inhibition of inducible NO synthase (iNOS) expression differed considerably from compound to compound (12, 14, 16). Furthermore, among these 6 compounds, only diethyl 2-chloroazulene-1,3-dicarboxylate effectively inhibited PGE2 production by LPS-stimulated RAW264.7 cells. These data demonstrated the lack of correlation between the extent of inhibition of iNOS protein expression by tropolone or azulene derivatives and that of PGE2 (24).
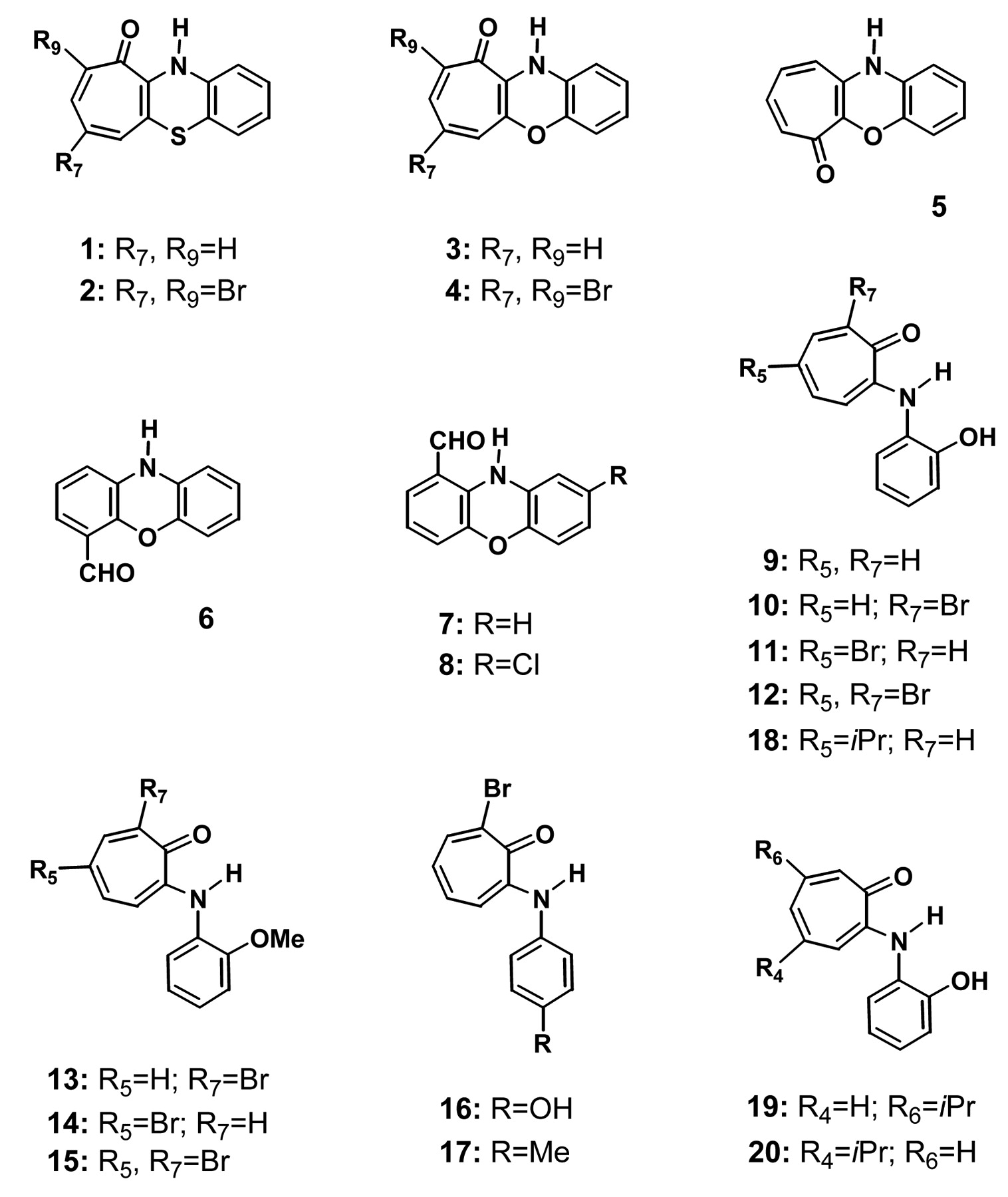
Differing from unreactive heterocyclic-annulated tropylium compounds, benzo[b]cyclohept[e][1,4]oxazines and their S-analogs are generally very reactive, especially towards 1,4-difunctional nucleophiles such as O-phenylenediamine, ethylenediamine and their S- and O-analogs (26). We have recently reported the tumor-specific cytotoxicity of a total of twenty benzo[b]cyclohept[e][1,4]oxazines and their S-analogs, and 2-aminotropone derivatives (25). In order to confirm the anti-inflammatory potency of these twenty compounds (Figure 1), we investigated whether they inhibit NO production by LPS-stimulated RAW264.7 cells. In the arachidonic acid (AA) cascade, cycloxygenase (COX) catalyzes the rate-limiting reactions for prostaglandin (PG) synthesis comprising the bis-cyclooxygenation of AA to form PGG2 and the peroxidative reduction of this intermediate to PGH2 (27). Since COX has two isoforms, COX-1 (constitutive isoform) and COX-2 (inducible isoform) (28), we also investigated whether these compounds inhibit the expression of COX-2 as well as iNOS at protein and mRNA levels.

Chemical structure of benzo[b]cyclohepta[e][1,4]benzoxazines and their S-analogs [1-8], and 2-aminotropones [9-20].
Materials and Methods
Materials. The following chemicals and reagents were obtained from the indicated companies: Dulbecco's modified Eagle's medium (DMEM) (Gibco BRL, Grand Island, NY, USA); fetal bovine serum (FBS) (JRH Bioscience, Lenexa, KS, USA); LPS from Escherichia coli (Serotype 0111:B4), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) (Sigma Chem Co., St Louis, MO, USA); dimethyl sulfoxide (DMSO) (Wako Pure Chem. Ind., Osaka, Japan); 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxy1-3-oxide (carboxy-PTIO; a spin trap agent), 1-hydroxy-2-oxo-3-(N-3-methyl-3-aminopropyl)-3-methyl-1-triazene (NOC-7; an NO generator), diethylenetriaminepentaacetic acid (DETAPAC), 5,5-dimethyl-1-pyrroline-N-oxide (DMPO) (Dojin, Kumamoto, Japan).
Benzo[b]cyclohepta[e][1,4]benzoxazines and their S-analogs [1-8], and 2-aminotropolones [9-20] synthesized in this study.
Synthesis of benzo[b]cyclohept[e][1,4]oxazines and their S-analogs, and 2-aminotropolones. Benzo[b]cyclohept[e][1,4]oxazines and their S-analogs [1-8] and 2-aminotropone derivatives [9-20] (Figure 1, Table I) were synthesized as described elsewhere (25).
Cell culture. RAW264.7 cells (29) were cultured as an adherent monolayer culture in DMEM supplemented with 10% heat-inactivated FBS under a humidified 5% CO2 atmosphere, as reported elsewhere (23). When the cells were grown to confluency, they were detached by being splashed with the medium through the pipette, and seeded at lower cell density. We did not collect the cells by scraping them out from the plate with a rubber policeman, since the scraping procedure damaged the cells.
Determination of viable cell number. RAW264.7 cells were inoculated at 0.3×106/ml (100 μl) in a 96-microwell plate (Becton Dickinson Labware, NJ, USA) and incubated for 24 hours. Near confluent cells were treated for a further 24 hours with the indicated concentrations of test compound in phenol red-free DMEM supplemented with 10% FBS in the presence or absence of LPS (0.1 μg/ml). The culture supernatant was collected for the determination of NO as described below. The attached cells were used for the determination of viable cell number by MTT method. In brief, cells were incubated for 60 minutes with 0.2 mg/ml of MTT. After removal of the medium, the cells were lysed with DMSO, and the absorbance at 540 nm was recorded using a microplate reader (23).
Effect of benzo[b]cyclohept[e][1,4]oxazine and 2-aminotropone derivatives on NO production by LPS-stimulated RAW264.7 cells.
Assay for NO production. RAW264.7 cells were treated with test compounds as described above, and the concentration of NO released from the cells was determined by Greiss reagent, using the standard curve of NO2−. To eliminate the interaction between sample and Greiss reagent, we also measured the NO concentration in the culture medium without the cells and subtracted this value from that obtained with the cells. The concentration that inhibited the LPS-stimulated NO production by 50% (50% inhibitory concentration: IC50) was determined from the dose-response curve (23). The efficacy of inhibition of NO production was evaluated by the selectivity index (SI), which was calculated using the following equation: SI=CC50/IC50, where CC50 was the cytotoxic concentration causing death of 50% of the population of cells.
Assay for iNOS protein expression. RAW264.7 cells were inoculated at 1.0×106/ml (100 μl) in 96-well plates (Becton Dickinson) and incubated for 24 hours. Near-adherent cells were treated for 24 hours with different concentrations of test sample. The concentrations were selected by CC50 in the presence or absence of 0.1 μg/ml LPS. The cell pellets were lysed with 50 μl of lysis buffer [10 mM Tris-HCl (pH 7.6), 1% Triton® X-100, 150 mM NaCl, 5 mM EDTA-2Na and 2 mM PMSF] for 10 minutes on ice. The cell lysates were centrifuged at 16,000×g for 20 minutes at 4°C to remove insoluble materials and the supernatant was collected. The protein concentrations of the supernatant were measured by Protein Assay Kit (Bio Rad, Hercules, CA, USA). Equal amounts of the protein from cell lysates (10 μg) were mixed with 2×sodium dodecyl sulfate (SDS)-sample buffer [0.1 M Tris-HCl (pH 6.8), 20% glycerol, 4% SDS, 0.01% bromphenol blue, 1.2% 2-mercaptoethanol], boiled for 10 minutes, and applied to SDS-8% polyacrylamide gel electrophoresis, then transferred for 90 minutes to polyvinylidene difluoride (PVDF) membrane. The membranes were blocked with 5% skim milk in phosphate-buffered saline [PBS (-)] plus 0.05% Tween 20 for 60 minutes and incubated for 90 minutes at room temperature with anti-iNOS or anti-COX-2 (dilution: 1:1,000 and 1:4,000, respectively, Santa Cruz Biotechnology, Delaware, CA, USA) or anti-actin antibody (1:6,000; Sigma Chem Co.), and then incubated with horseradish peroxidase-conjugated anti-goat, anti-rabbit or anti-mouse IgG for 60 minutes at room temperature. Immunoblots were developed with Western Lighting™ Chemiluminescence Reagent Plus (21).
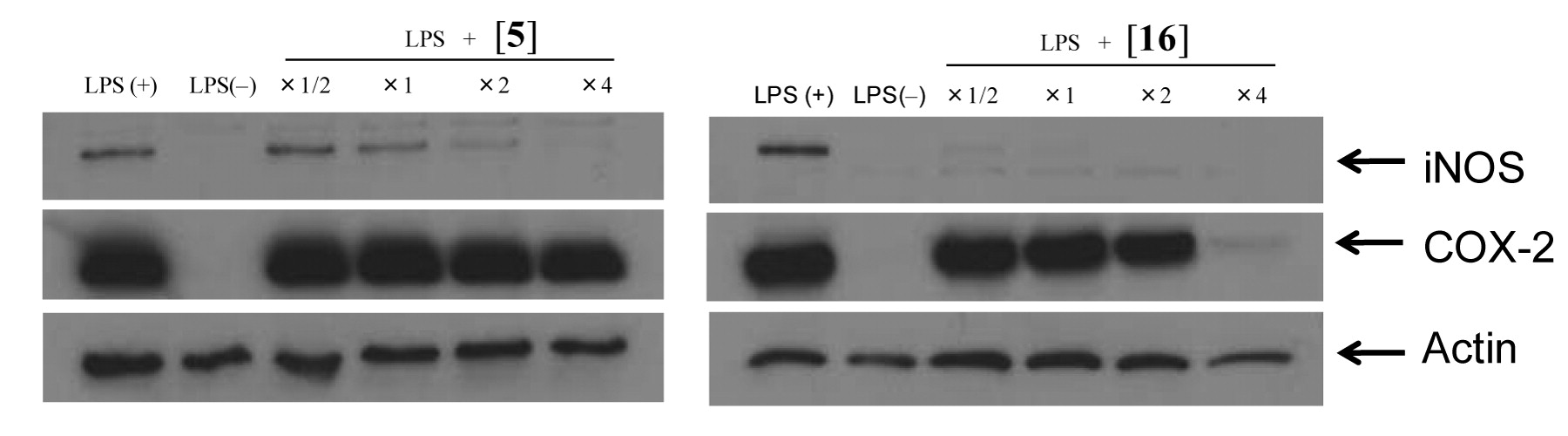
Effect of benzo[b]cyclohept[e][1,4]oxazin-6(11H)-one [5] and 7-bromo-2-(4-hydroxyanilino)tropone [16] on the LPS-stimulated expression of iNOS and COX-2 proteins. RAW264.7 cells (100×104 cells/ml, 1 ml) were inoculated on 24-well plate and incubated for 24 hours. RAW264.7 cells were then treated for 24 hours without (control) or with ×1/2, ×1, ×2 or ×4 of IC50 (concentration that inhibited the LPS-stimulated NO production by 50%) of benzo[b]cyclohept[e][1,4]oxazin-6(11H)-one [5] or 7-bromo-2-(4-hydroxyanilino)tropone [16] in the presence or absence of LPS (0.1 μg/ml) in the presence or absence of LPS (0.1 μg/ml), and then the expression of iNOS and COX-2 proteins were monitored by Western blot analysis.
Assay for iNOS and COX-2 mRNA expression. RAW264.7 cells were inoculated at 1.0×106/ml (100 μl) in 96-well plates and incubated for 24 hours. Near-confluent cells were treated for 24 hours with different concentrations of test compound in the presence or absence of LPS (0.1 μg/ml). The expression levels of iNOS and COX-2 mRNA were measured by RT-PCR. The primer sequence and size was: (i) iNOS: PCR product size 496 bp, sense primer 5’-CCCTTCCGAAGTTTCTGGCAGCAGC-3’, antisense primer 5’-GGCTGTCAGAGCCTCGTGGCTTTGG-3’; (ii) COX-2: Product size 371 bp, sense primer 5’-TTTGTTGAGTCATTCACCAG ACAGAT-3’, antisense primer 5’-CAGTATTGAGGAGAACAGA TGGGATT-3’; (iii) G3PDH: Product size 452 bp, sense primer 5’-ACCACAGTCCATGCCATCAC-3’, antisense primer 5’-TCCACC ACCCTGTTGCTGTA-3’ (21). After amplification, each reaction mixture was analyzed by 2% agarose gel electrophoresis and visualized by ethidium bromide staining under UV light. The relative amount of PCR product was normalized to the corresponding amount of G3PDH PCR product.
Radical-scavenging activity. The radical intensity of test samples was determined at 25°C, using electron-spin resonance (ESR) spectroscopy (JEOL JES REIX Tokyo, Japan; X-band, 100 kHz modulation frequency) (21, 23). The radical intensity of NO produced from the reaction mixture of 20 μM carboxy-PTIO and 60 μM NOC-7 was determined in 0.1 M phosphate buffer (PB; pH 7.4) in the presence of 30% DMSO (microwave power and gain were 5 mW and 400, respectively). When NOC-7 and carboxy-PTIO were mixed, NO was oxidized to NO2 and carboxy-PTIO was reduced to carboxy-PTI, which produces seven-line signals. Samples were added 3 minutes after mixing. The NO radical intensity was defined as the ratio of the signal intensity of the first peak of carboxy-PTI to that of MnO.
The superoxide anion (in the form of DMPO-OOH), produced by the HX-XOD reaction (total volume: 200 μl) [2 mM HX in 0.1 M (pH 7.4) (PB) 50 μl, 0.5 mM DETAPAC 20 μl, 8% DMPO 30 μl, sample 40 μl, H2O 30 μl, XOD (0.5 U/ml in PB) 30 μl] was determined (microwave power: 16 mW, gain: 630) (30).
Results
Inhibition of NO production. Twenty benzo[b]cyclohept[e] [1,4]oxazines and their S-analogs, and 2-aminotropone derivatives inhibited the production of NO by LPS-activated mouse macrophage-like RAW264.7 cells to different extents. Among them, benzo[b]cyclohept [e][1,4]oxazin-6(11H)-one [5] and 7-bromo-2-(4-hydroxyanilino)tropone [16] showed the highest inhibitory effects at concentrations that did not affect the cellular viability (SI=74.89 and 54.15, respectively, Table II).
Effect on iNOS and COX-2 expression. Western blot analyses showed that both compound [5] and [16] inhibited expression of iNOS protein (Figure 2). Compound [16] inhibited COX-2 protein expression at a concentration twice and four times that of the IC50 (50% inhibitory concentration of LPS-stimulated NO production). On the other hand, [5] did not completely inhibit the COX-2 protein expression even at concentrations 4 times that of the IC50.
RT-PCR analysis showed that [16] but not [5] inhibited iNOS and COX-2 mRNA expression at a concentration twice and four times that of the IC50 (Figure 3).

Effect of benzo[b]cyclohept[e][1,4]oxazin-6(11H)-one [5] and 7-bromo-2-(4-hydroxyanilino)tropone [16] on the LPS-stimulated expression of iNOS and COX-2 mRNA. RAW264.7 cells (100×104 cells/ml, 1 ml) were inoculated on 24-well plate and incubated for 24 hours. RAW264.7 cells were then treated for 24 hours without (control) or with ×1/2, ×1, ×2 or ×4 of IC50 (concentration that inhibited the LPS-stimulated NO production by 50%) of benzo[b]cyclohept[e][1,4]oxazin-6(11H)-one [5] or 7-bromo-2-(4-hydroxyanilino)tropone [16] in the presence or absence of LPS (0.1 μg/ml) in the presence or absence of LPS (0.1 μg/ml), and then the expression of iNOS and COX-2 mRNA was monitored by RT-PCR analysis.
NO-scavenging activity. There was a possibility that the reduced concentration of NO in the culture medium of the cells treated with either [5] or [16] may have been due to the NO-scavenging activity of these compounds. ESR spectroscopy revealed that both [5] and [16] scavenged NO generated from NOC-7 only at much higher concentrations (30-200 times higher than the concentration of IC50 for the inhibition of NO production; Figure 4A), demonstrating that this possibility was small. Since NO and O2- interact with each other, we investigated whether [5] and [16] scavenge the O2- generated by the HX-XOD reaction. ESR spectroscopy revealed that these compounds also scavenged O2- very poorly (Figure 4B).
Discussion
The present study demonstrated that all twenty benzo[b] cyclohept[e][1,4]oxazine and 2-aminotropone derivatives investigated showed an inhibitory effect on the NO production by LPS-activated RAW264.7 cells to different extents, and that among them, benzo[b]cyclohept[e] [1,4]oxazin-6(11H)-one [5] and 7-bromo-2-(4-hydroxyanilino)tropone [16] were the most potent inhibitors. However, these two compounds seem to inhibit LPS-stimulated NO production by different mechanisms.
Compound [16] inhibited iNOS protein expression more efficiently than that of NO production, and therefore the inhibition of NO production by [16] was due to the inhibition of iNOS production (Figure 2) resulting from impaired mRNA expression, not due to its NO-scavenging activity (Figure 4). Compound [16] also inhibited the expression of COX-2 protein (Figure 2) and mRNA expression (Figure 3), but at slightly higher concentrations (×2 and ×4 of IC50 of NO production). These data suggest the possible anti-inflammatory activity of [16]. Higher tumor specificity of this compound (25) makes the further investigation of [16] promising.
Similarly, the inhibition of LPS-stimulated NO production by compound [5] was due to the inhibition of the induction of iNOS protein expression, not due to its NO-scavenging activity. However, [5] failed to inhibit iNOS mRNA expression. It remains to be investigated whether [5] induces post-transcriptional modification of iNOS, and whether it induces the S-nitrosylation of COX-2 that is responsible for the decline of this enzyme's activity (31).
In conclusion, the present study suggests that the inhibition of NO production by macrophages is not an appropriate parameter for evaluating anti-inflammatory activity, supporting our recent report (24). Other markers, such as PGE2 production or COX-2 expression, should be monitored in addition to NO production.

Low scavenging activity of benzo[b]cyclohept[e][1,4]oxazin-6(11H)-one [5] and 7-bromo-2-(4-hydroxyanilino)tropone [16] against NO and O2-. Each value represents the mean±S.D. of triplicate assays.
Acknowledgements
This study was supported in part by a Grant-in-Aid from the Ministry of Education, Science, Sports and Culture of Japan (Sakagami, No. 19592156).
- Received March 17, 2009.
- Revision received June 3, 2009.
- Accepted July 2, 2009.
- Copyright © 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Enhancement of Cytotoxicity of Three Apoptosis-inducing Agents Against Human Oral Squamous Cell Carcinoma Cell Line by Benzoxazinotropone
- Anti-inflammatory Potential of Rikkosan Based on IL-1{beta} Network Through Macrophages to Oral Tissue Cells
- Quantitative Structure-Activity Relationship Analysis of Cytotoxicity and Anti-UV Activity of 2-Aminotropones
- Cytotoxic Activity of Benzo[b]cyclohept[e][1,4]oxazines
- Quest for Anti-inflammatory Substances Using IL-1{beta}-stimulated Gingival Fibroblasts
- Hormetic and UV-Protective Effects of Azulene-related Compounds
- Hormetic and Anti-radiation Effects of Tropolone-related Compounds
- Hormetic Response of Cultured Normal and Tumor Cells to 2-Aminotropone Derivatives