


Abstract
Mastic is a resinous exudate obtained from the stem and the main leaves of Pistacia lentiscus. We have reported the antiplaque effect of mastic-containing chewing gum on the oral cavity. We hypothesize that mastic may be a multifunctional food which has some beneficial pharmaceutical properties. The aim of this study was to assess the biological activity of solid and liquid types of mastic by cytotoxicity against fibroblasts, radical-scavenging activities and inhibitory effect on cell death of oral polymorphonuclear leukocytes (OPMNs). Mastic showed selective antibacterial action against Porphyromonas gingivalis and Prevotella melaninogenica, but no anti-HIV activity. Among a total of thirteen human cell types, promyelocytic leukemia HL-60 was the most sensitive to the cytotoxicity of mastic, followed by myeloblastic leukemia (ML-1, KG-1), erythroleukemia (K-562), oral squamous cell carcinoma (HSC-2, HSC-3, HSC-4), hepatocellular carcinoma (HepG2), glioblastoma (T98G, U87MG) and normal oral cells (gingival fibroblast, pulp cell, periodontal ligament fibroblast, most resistant). Mastic did not induce the differentiation of myelogenous leukemic cells into maturing cells with higher nitroblue tetrazolium-reducing activity, but induced apoptotic cell death, characterized by internucleosomal DNA fragmentation, caspase-3 activation and a decline in the intracellular concentration of putrescine. The cytotoxicity of mastic against leukemic cells did not diminish during its storage. On the other hand, mastic inhibited the spontaneous apoptosis of OPMNs. Mastic showed hydroxyl radical-scavenging activity. The selective antibacterial and apoptosis-modulating activity of mastic suggests its possible beneficial effects on oral health.
Certain plants used in folk medicine serve as sources of therapeutic agents containing antibacterial agents and other medicinal compounds. The effective substances isolated from these plants have been shown to have multipotential effects. Mastic is the extract of sap from Pistacia lentiscus, grown only in the Chios island of Greece. Due to its unique shape and diverse efficacy, mastic has been called “the tear drop of Christ”. This extract has been utilized for as long as 5,000 years as a food for the stimulation of health and the treatment of digestive diseases (1). Mastic has shown therapeutic effects on stomach and duodenal ulcers (2-4) and has also shown antibacterial activity against Helicobacter pylori (5, 6). Recently, major triterpenic acids in the acid extract have been suggested to be responsible for anti-H. pylori activity (7). Mastic chewing gum has also shown antiplaque activity in the oral cavity (8). Mastic has been reported to inhibit the growth of colon cancer (9), prostate cancer (10), and erythroleukemia cell line (11). However, the comparative cytotoxicity of mastic against both normal and tumor cell lines has not been well characterized. We investigated here the antibacterial spectrum, anti-HIV activity and cytotoxic activity of mastic (Figure 1) against a total of thirteen types of human normal and tumor cells.
Materials and Methods
Chemicals. The following chemicals and reagents were obtained from the indicated companies: mastic (solid form, liquid form; Figure 1) (Nakamura Chiro Association, Tokyo, Japan); Dulbecco's modified Eagle's medium (DMEM, GIBCO BRL, Grand Island, NY, USA); fetal bovine serum (FBS; JRH, Bioscience, Lenexa, KS, USA), RPMI-1640, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), all-trans retinoic acid (ATRA), nitroblue tetrazolium (NBT), azidothymidine (AZT), 2′,3′-dideoxycytidine (ddC) (Sigma Chem. Co., St Louis, MO, USA); dimethyl sulfoxide (DMSO) (Wako Pure Chem. Ind., Osaka, Japan), human recombinant granulocyte-macrophage colony-stimulating factor (rHuGM-CSF) (Sandoz, Wien, Austria).
Antibacterial activity. Streptococcus mutans and Streptococcus gordonii were grown anaerobically at 37°C in Todd Hewitt broth (Difco Laboratories, Detroit, MI, USA). Actinomyces viscosus, Capnocytophaga ochracea, Fusobacterium nucleatum, Prevotella intermedia, Prevotella melaninogenica and Porphyromonas gingivalis were cultivated anaerobically at 37°C in Brain Heart Infusion broth (Difco Laboratories) supplemented with hemin (5 μg/ml) and menadione (50 μg/ml). Candida albicans, Escherichia coli and Staphylococcus aureus were grown aerobically at 37°C in nutrient broth (Eiken Chemical Co., LTD., Tokyo, Japan). Each bacterium was cultivated for 2 days with or without 2% mastic and the antibacterial activity was determined by their growth in each medium (12).
Assay for anti-HIV activity. Human T-cell leukemia MT-4 cells were infected with HIV-1IIIB at a multiplicity of infection (m.o.i.) of 0.01. HIV- and mock-infected (control) MT-4 cells were incubated for 5 days with a range of concentrations of test samples (solid mastic, 0-200 μg/ml; liquid mastic, 0-0.0006%; AZT, 0-1000 μM; ddC, 0-10000 μM), and the relative viable cell number was determined by MTT assay. The 50% cytotoxic concentration (CC50) and 50% effective concentration (EC50) were determined from the dose-response curve with mock-infected and HIV-infected cells, respectively. All data represent the mean values of triplicate measurements. The anti-HIV activity was evaluated by the selectivity index (SI), which was calculated by the following equation: SI=CC50/EC50 (13).
Cell culture. Normal human oral cells (gingival fibroblast HGF, pulp cell HPC, periodontal ligament fibroblast HPLF) were prepared from the periodontal tissues, according to the guideline of Meikai University Ethics Committee (No. 0808), after obtaining informed consent from healthy volunteers. Since normal oral cells have a limited lifespan, all of them ceasing proliferation at the 20 population doubling level (PDL) (14), these cells were used at 7-12 PDL in the present study. Human squamous cell carcinoma cell lines [HSC-2, HSC-4 (supplied by Professor M. Nagumo, Showa University), HSC-3 (supplied by Professpr Y. Ohmori, Meikai University)], human hepatocellular carcinoma (HepG2) (supplied by Professor T. Tobe, Showa University), human glioblastoma cell lines (T98G, U87MG) (supplied by Dr. M. Iida, Showa University) and human normal cells (HGF, HPC, HPLF) were cultured in DMEM supplemented with 10% heat-inactivated FBS in a humidified 5% CO2 atmosphere. Human promyelocytic leukemia HL-60 and human erythroleukemia K-562 cells (supplied by Professor K. Nakaya, Showa University) and human myeloblastic leukemia (ML-1, KG-1) cell lines (supplied by Professor K. Takeda, Tokyo University of Science) were cultured in suspension in RPMI-1640 medium supplemented with 10% heat-inactivated FBS. The normal cells were harvested by detaching with 0.25% trypsin-0.025% EDTA-2Na in phosphate-buffered saline without Mg and Ca [PBS(-)] and subcultured at a 1:4 split ratio once a week, with one medium change in between. The adherent tumor cell lines were similarly trypsinized and subcultured twice a week.
Preparation of polymorphonuclear leukocytes from the oral cavity (OPMNs). OPMNs were prepared by the modification of a previous report (15). In brief, OPMNs from 2 to 3 healthy volunteers were collected by gargling with 25 ml PBS (-) eight times. All operations were performed at 4°C. The cells were collected by centrifugation at 640 xg for 5 minutes and passed through 50 μm nylon mesh (Tokyo Screen, Tokyo, Japan) to remove the oral mucoepithelial cells and bacterial mass. The cells were resuspended with 3 ml PBS(-), and overlayered onto 1 ml of Mono-Poly resolving medium® (Dainippon Pharm Co. Tokyo, Japan). After centrifugation at 100 xg for 10 minutes and then at 1,000 xg for 5 minutes, OPMNs were collected from the interface between PBS and Mono-Poly resolving medium®. OPMNs were then washed once and resuspended at 1×106/ml in PBS(-).
Assay for cytotoxic activity. Near-confluent cells were treated for 24 hours with 0-200 μg/ml (solid mastic) or 0-2 (v/v)% of liquid mastic. The number of viable adherent cells was then determined by the MTT method. In brief, the cells were washed once with PBS(-), and incubated for 4 hours with 0.2 mg/ml of MTT in the culture medium. After removing the medium, the cells were lysed with 100 μl DMSO and the absorbance at 540 nm of the cell lysate (the relative viable number) was measured by a microplate reader (Labsystems Multiskan; Biochromatic Labsystem, Osaka, Japan). The number of viable non-adherent cells (HL-60, ML-1, KG-1, K-562) was determined by cell count with a hemocytometer after staining with 0.15% trypan blue. The CC50 was determined from the dose-response curve.
Assay for DNA fragmentation. Cells (5×105/ml, 1 ml) were treated for 6 or 24 hours with 0-160 μg/ml solid mastic. The cells were washed once with PBS(-) and lysed with 50 μl lysate buffer [50 mM Tris-HCl (pH 7.8), 10 mM EDTA, 0.5% (w/v) sodium N-lauroylsarcosinate]. The solution was incubated with 0.4 mg/ml RNase A and 0.8 mg/ml proteinase K for 2 hours at 50°C and then mixed with 50 μl NaI solution [40 mM Tris-HCl (pH 8.0), 7.6 M NaI, 20 mM EDTA-2Na] and then 250 μl of ethanol. After centrifugation for 20 minutes at 20,000 xg, the precipitate was washed with 1 ml of 70% ethanol and dissolved in TE buffer [10 mM Tris-HCl (pH 8.0), 1 mM EDTA-2Na]. Each sample (10 μl) was applied to 2% agarose gel electrophoresis in TBE buffer (89 mM Tris-HCl, 89 mM boric acid, 2 mM EDTA-2Na). After staining with ethidium bromide, the DNA was visualized as described above (16). As a positive control for internucleosomal DNA fragmentation, apoptotic HL-60 cells induced by UV irradiation were prepared as described elsewhere (17).
Assay for caspase activation. Cells were treated for 4 or 4.5 hours with 0-160 μg/ml solid mastic. Cells were washed with PBS(-) and lysed in lysis solution [50 mM Tris-HCl (pH 7.5), 0.3% NP-40, 1 mM DTT]. After standing for 10 minutes on ice and centrifugation for 5 minutes at 21,000 xg, the supernatant was collected. Lysate (50 μl, equivalent to 200 μg protein) was mixed with 50 μl 2x lysis solution containing substrates for caspase-3 [DEVD-pNA (p-nitroanilide)], caspase-8 (IETD-pNA) or caspase-9 (LEHD-pNA). After incubation for 2 hours at 37°C, the absorbance at 405 nm of the liberated chromophore pNA was measured by microplate reader (16).
Effect of mastic on bacteria. Mastic clearly has a selective antibacterial action against P. gingivalis and P. melaninogenica.
Assay for differentiation induction. Cells (5×105/ml, 225 μl) were inoculated on 96-microwell plates and 25 μl mastic solution (final concentration: CC50, 1/2CC50, or 1/5CC50) were added. After incubation for 24 hours, NBT reagent (50 μl/well) was added and the cells were further incubated for 30 minutes at 37°C. To stop the reaction, 3 N HCl (50 μl/well) was added and stood on ice for 30 minutes. The NBT reagent was removed and then 200 μl of DMSO were added. The absorbance at 595 nm was then measured by plate reader (18).
Determination of polyamines. The cells were washed twice with PBS(-) and extracted with 10% trichloroacetic acid. After centrifugation for 5 minutes at 10,000 xg, the deproteinized supernatant was collected. The polyamines in the supernatant were determined by HPLC, after Dansyl-derivatization, as described elsewhere (16).
Radical scavenging activity. The radical intensity of solid mastic was determined at 25°C, using electron-spin resonance (ESR) spectroscopy (JEOL JES REIX, X-band, 100 kHz modulation frequency) (19). For the determination of hydroxyl radical [in the form of 5,5-dimethyl-1-pyrroline-N-oxide (DMPO)-OH], produced by the Fenton reaction (200 μl)[1 mM FeSO4 (containing 0.2 mM diethylenetriamine-N,N,N′,N“,N″′-pentaacetic acid) 50 μl, 0.1 M phosphate buffer (pH 7.4) 50 μl, 92 mM DMPO 20 μl, sample 50 μl in water, 1 mM H2O2, 30 μl], the gain was changed to 400 (20). Mastic was suspended in water, since DMSO quenched the hydroxyl radical (data not shown).
NO-scavenging activity was assessed by nitrate/nitrite colorimetric assay. NaNO2 solution (100 μM) was added to a range of concentrations of mastic (0-3 mg/ml), reacted with Griess reagent and the absorbance at 540 nm/620 nm measured (19).
Statistical analysis. Differences between treated and untreated cells were statistically treated by using an unpaired Student's t-test.
Cytotoxicity of mastic towards MT-4 HIV-infected cells. Mastic showed no anti-HIV activity.
Results
Antibacterial spectrum. Liquid type of mastic (2%) showed selective antibacterial action against Porphyromonas gingivalis and Prevotella melaninogenica, as compared with that against the growth of Actinomyces viscosus, Streptococcus gordonii, Streptococcus mutans, Capnocytophaga ochracea, Fusobacterium nucleatum, Prevotella intermedia, Staphylococcus aureus, Escherichia coli and Candida albicans (Table I).
Anti-HIV activity. Neither solid nor liquid mastic had any anti-HIV activity (SI<1.0), whereas popular anti-HIV agents, AZT and ddC, showed much higher anti-HIV activity (SI=11567 and 1320, respectively) (Table II).
Cytotoxic activity. Solid mastic showed highest cytotoxicity against human leukemic cell lines (promyelocytic leukemia HL-60, myeloblastic leukemia ML-1, KG-1, erythroleukemia K-562) [CC50=33-77 μg/ml (high cell density, HCD); 19-30 μg/ml (low cell density, LCD)]; followed by human oral squamous cell carcinoma cell lines (HSC-2, HSC-3, HSC-4) (110-126; 81-90 μg/ml) and human hepatocellular carcinoma cell line (HepG2) (144; 48 μg/ml). Human glioblastoma cell lines (T98G, U87MG) (143-158; 127-138 μg/ml) and human normal oral cells (gingival fibroblast HGF, pulp cell HPC, periodontal ligament fibroblast HPLF) (136-166; 93-155 μg/ml) were the most resistant to mastic (Table III).
Similarly, the liquid mastic showed the highest cytotoxicity againt leukemic cell lines (CC50=0.036-0.18% at HCD; 0.010-0.11% at LCD), followed by squamous cell carcinoma cell lines (0.26-2.0; 0.24-1.9%). Glioblastoma, hepatocellular carcinoma cell lines and normal oral cells were much more resistant (CC50>2.0%). Liquid mastic showed higher specificity of antileukemic activity than did solid mastic (Table III). Solid mastic stored for more than 2 years turned yellowish in color, but showed comparable cytotoxicity with fresh mastic (Figure 2).
Selective cytotoxicity of mastic against human leukemic cell lines.
Induction of apoptosis in human leukemic cells. We next investigated whether the selective cytotoxicity of mastic against human leukemic cell lines is due to the induction of differentiation or that of apoptosis. We found that mastic, both solid and liquid at the CC50, 1/2 CC50 and 1/5 CC50 failed to induce differentiation of HL-60 and ML-1 cells, whereas all-trans retinoic acid (ATRA) (1 μM), or granulocyte-macrophage colony-stimulating factor (GM-CSF) (0.1 ng/ml) together with ATRA (0.1 μM) (18) induced differentiation (accompanied by elevated NBT-reducing activity) (Figure 3).
We found that solid mastic induced apoptotic cell death, characterized by the decline of proliferation, decline of intracellular putrescine level and induction of internucleosomal DNA fragmentation and caspase-3 activation, in HL-60 cells (Figure 4). Mastic induced internucleosomal DNA fragmentation (Figure 5) and caspase-3 activation (Figure 6) in three leukemic cell lines (HL-60, ML-1, KG-1).
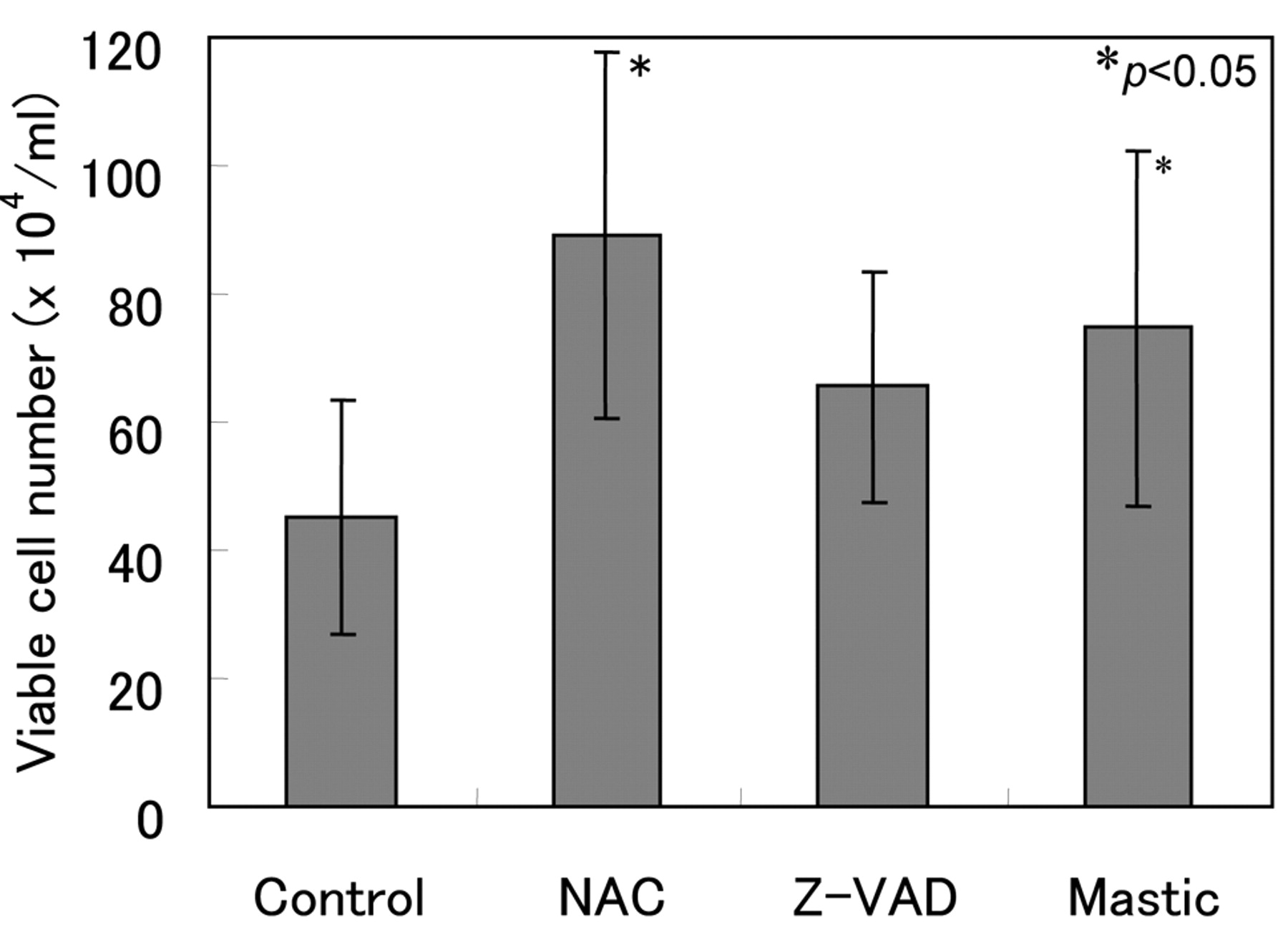
Inhibition of spontaneous apoptosis of OPMNs. We have previously reported that OPMNs undergo spontaneous apoptosis characterized by internucleosomal DNA fragmentation, and that L-cysteine, reduced glutathione (GSH) and herbimycin A, a protein tyrosine kinase inhibitor, suppressed the activation of caspase-3 and the apoptosis of OPMNs (21, 22). Moreover, N-acetyl-Asp-Glu-Val-Asp-aldehyde (Ac-DEVD-CHO), an inhibitor of caspase-3, inhibited the fragmentation of DNA. These results suggested that oxidative stress and/or tyrosine kinase-dependent pathway(s) activated caspase-3 in OPMNs, thereby inducing their apoptosis (21, 22). We found that mastic, as well as a popular antioxidant (N-acetyl-l-cysteine, NAC) and caspase inhibitor (Z-VAD) inhibited the spontaneous cell death of OPMNs (Figure 7). Mastic failed to scavenge the NO produced by NOC-7, but effectively scavenged the hydroxyl radical generated by the Fenton reaction (Figure 8). This result suggests that mastic inhibits the spontaneous apoptosis of OPMN by its antioxidant activity.

Solid and liquid types of mastic used in the present study.

Effect of storage of mastic on its cytotoxicity against HL-60 cells. HL-60 cells were incubated for 48 hours with the indicated concentrations of fresh solid mastic (new) or that stored for 2 years (old), and the viable cell number was determined by trypan blue exclusion method. Each value represents the mean±S.D. from triplicate assays.
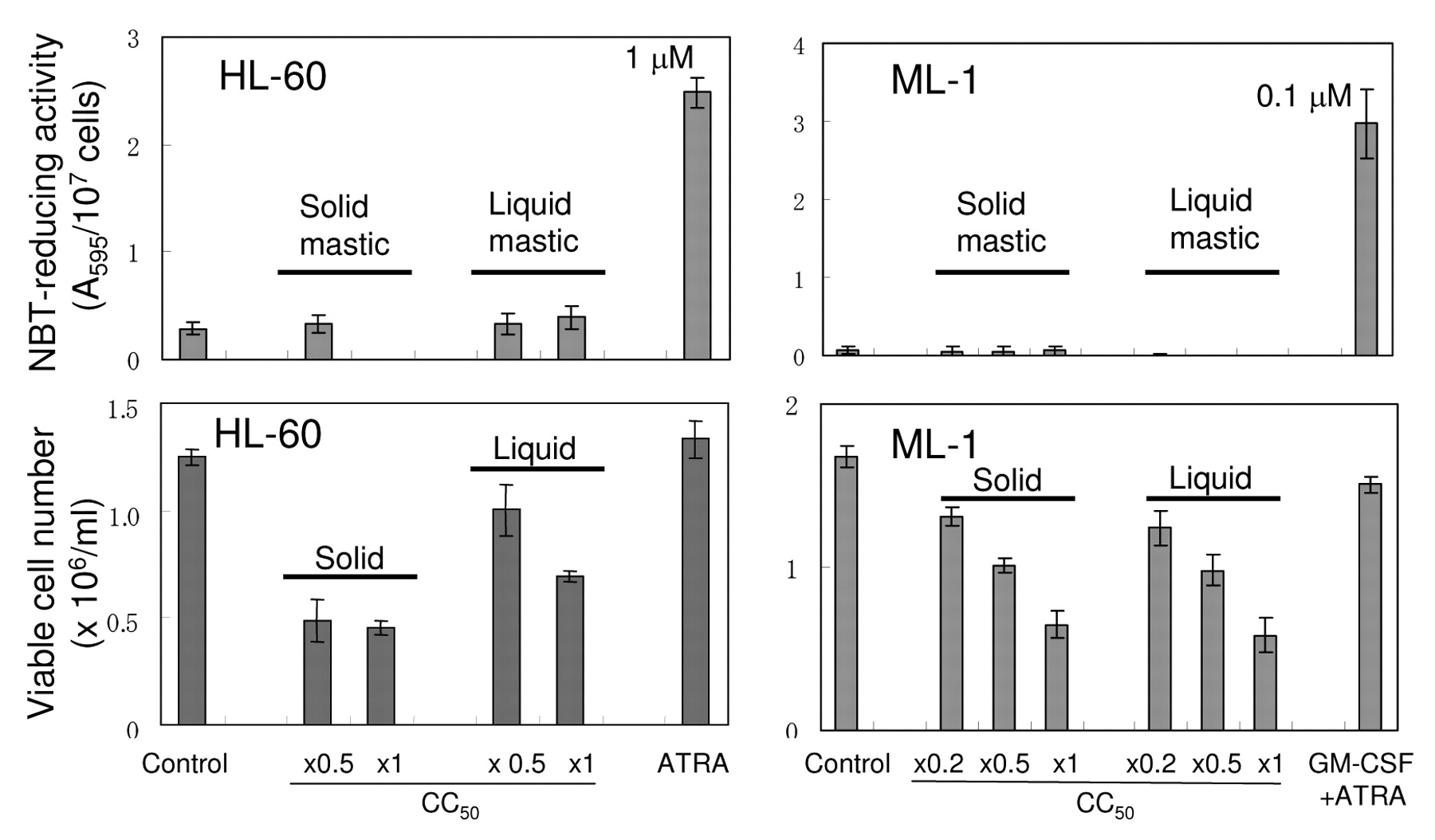
Failure of mastic to induce the differentiation of human myelogenous leukemic cell lines. HL-60 and ML-1 were treated for 24 hours with mastic or, all-trans retinoic acid (ATRA) in the presence or absence of granulocyte-macrophage colony-stimulating factor (GM-CSF) (0.1 ng/ml), and viable cell number and NBT-reducing activity were then determined. Similar results were obtained in KG-1 and K-562 cells (data not shown). Each value represents the mean±S.D. from triplicate assays.
Discussion
The present study demonstrated that mastic preferentially inhibited the growth of P. gingivalis and P. melaninogenica. This finding further supports the antibacterial activity of mastic chewing gum previously reported (8).
We also found that mastic inhibited the spontaneous apoptosis of OPMNs, possibly by its antioxidant activity. PMNs play crucial roles in protecting hosts against invading microbes and in the pathogenesis of inflammatory tissue injury. We previously reported that PMNs in the oral cavity are more fragile as compared with circulating PMNs (CPMNs), that they spontaneously generate superoxide radical and nitric oxide (NO) in the absence of any stimuli, and undergo spontaneously apoptosis (21, 22). The present finding demonstrated that mastic stimulated the survival of OPMNs and thus may contribute in the potentiation of natural immunity such as the elimination of bacteria from the oral cavity. Both the antibacterial and OPMN survival-promoting activities of mastic may improve oral health, thus leading to the reduction of oral malodor (23) (Figure 9).

Induction of apoptosis by mastic in HL-60 cells. HL-60 cells were incubated for 3, 4.5, 6 or 24 hours with the indicated concentrations of mastic, or 1 μg/ml actinomycin D, or subjected to UV irradiation (1 minute) followed by 6 hours' incubation, and then assayed for growth, DNA fragmentation, caspase activity and intracellular polyamine concentrations.

Induction of internucleosomal DNA fragmentation by mastic in human leukemic cell lines. Human leukemic cell lines were incubated for 6 or 24 hours with the indicated concentrations of mastic and then assayed for DNA fragmentation. UV, DNA from apoptotic HL-60 cells induced by UV irradiation.
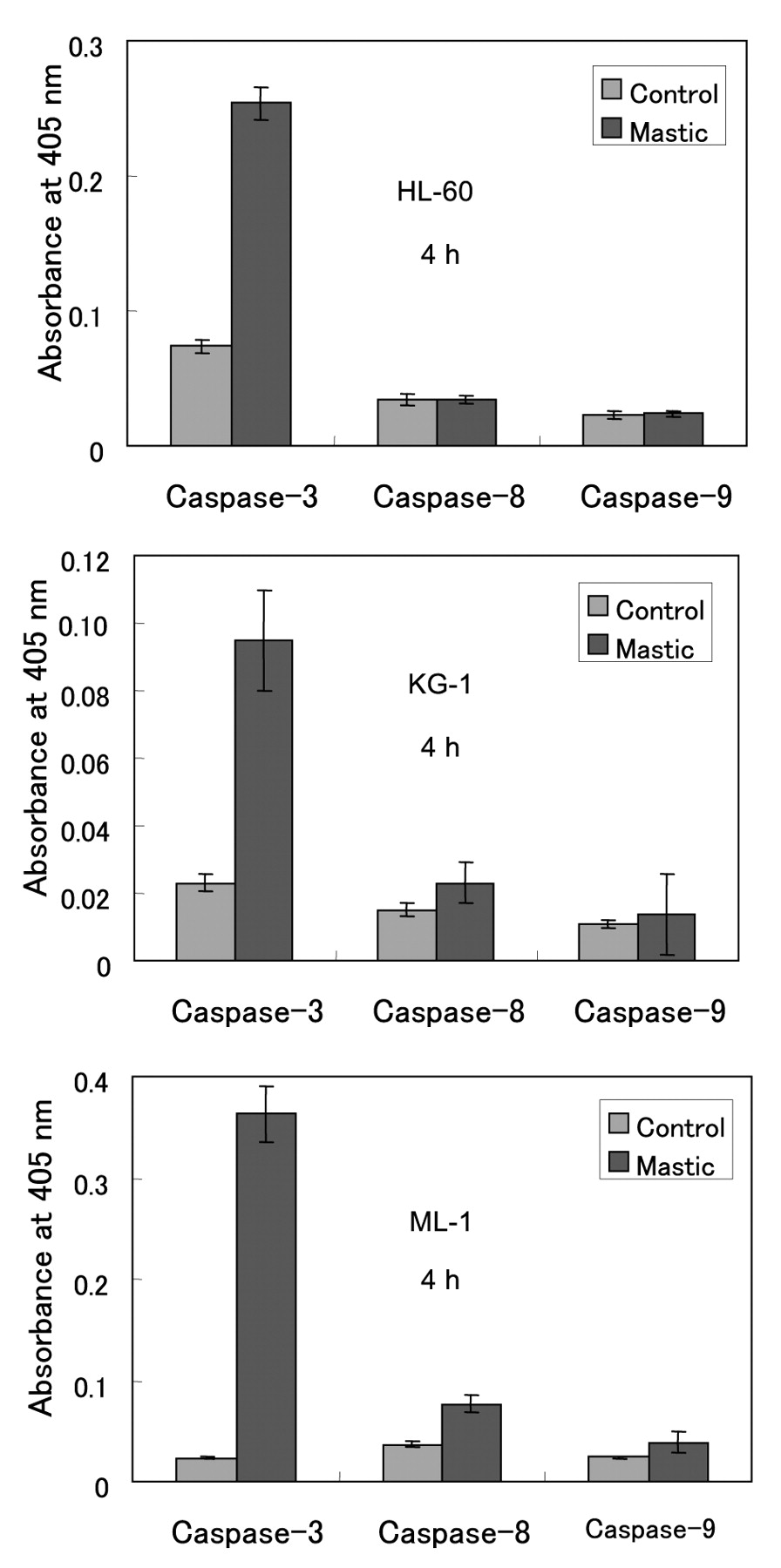
Induction of caspase-3 activity by mastic in human leukemic cell lines. Cells were incubated for 4 hours with or without mastic (80 μg/ml for HL-60 and KG-1 cells; 160 μg/ml for ML-1 cells), and then caspase activity was measured. Each value represents the mean±S.D. from four independent experiments.

Inhibition of cell death of OPMNs by mastic, antioxidant and caspase inhibitor. OPMNs (1×106/ml) were incubated at 4°C for 60 minutes without (control), or with N-acetyl-l-cysteine (NAC) (5 mM), or caspase inhibitor (Z-VAD) (0.15 mM), or with mastic (1 mg/ml), then incubated at 37°C for 60 min and the viable cell number was then counted. Each value represents the mean±S.D. from triplicate assays.

Mastic scavenged hydroxyl radical (*OH), but not nitric oxide (NO) radical. NO concentration was determined by the Griess method. Hydroxyl radical intensity was determined by electron-spin resonance. The radical intensity was expressed as a % that of the control.
Reactive oxygen species (ROS) have been of interest for many years in all areas of biology because of their importance in cell growth, differentiation and death by initiating cellular signaling pathways. Being a secondary messenger in cellular signal transduction, ROS alter the numbers of redox-sensitive enzymes which are critical in cellular activities such as mitogenesis, cell adhesion, oncogenic transformation and apoptosis (24). ROS are closely involved in various pathological events such as aging, carcinogenesis and inflammation. ROS have been detected in the oral cavity and thus it is attractive to use natural antioxidants such as mastic to reduce ROS in the oral cavity.

Hypothetical scheme of points of action of mastic.
The present study demonstrated for the first time that mastic selectively inhibited the growth of human leukemic cell lines and this was due to the induction of apoptosis, not to the induction of differentiation. We found that putrescine declined with mastic treatment. This is consistent with our previous finding that putrescine, but not spermidine and spermine, declined during the apoptosis induction of HL-60 cells (25, 26). Mastic has also been reported to induce apoptosis in colon cancer cells (9) and target the NF-κB signal in prostate cancer cells (10).
The cytotoxicity of mastic against leukemic cells was much more pronounced when the liquid form of mastic was used, rather than the solid form. The higher specificity of the liquid form may be due to its higher solubility when added to the culture medium. At present, the active principle for the cytotoxicity is unknown, although gas chromatography-mass spectroscopic analysis of penta- and tetracyclic triterpenes of the mastic resin demonstrated the presence of isomasticadienonic acid, masticadienonic acid and 28-norolean-17-en-3-one (27). Further fractionation of mastic may provide information to help elucidate the mechanism of antileukemic action of mastic. Among tumor cell lines, human oral squamous cell carcinoma cell lines (HSC-2, HSC-3, HSC-4) showed higher sensitivity than human glioblastoma cell lines (T98G, U87MG) and hepatocellular carcinoma cell line (HepG2). The expression of MDR proteins that play an important role in pumping out the drugs among these different types of cancer cell lines remains to be investigated.
In conclusion, mastic may improve the oral environment by its selective antibacterial activity and the prevention of OPMN apoptosis (Figure 9); it also appears to have antileukemia cell activity.
Acknowledgements
This study was supported in part by a Grant-in-Aid from the Ministry of Education, Science, Sports and Culture of Japan (Sakagami, No. 19592156; Takahashi, No. 19592210).
- Received July 29, 2008.
- Revision received December 2, 2008.
- Accepted January 9, 2009.
- Copyright © 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Data Mining a Medieval Medical Text Reveals Patterns in Ingredient Choice That Reflect Biological Activity against Infectious Agents
- Evaluation of Biological Activity of Mastic Extracts Based on Chemotherapeutic Indices
- Chios Mastic Gum: A Plant-produced Resin Exhibiting Numerous Diverse Pharmaceutical and Biomedical Properties
- Re-evaluation of Anti-inflammatory Activity of Mastic Using Activated Macrophages