


Abstract
Fibrosarcoma is an uncommon soft tissue tumor with a complex cell microenvironment, particularly rich in glycosaminoglycans/proteoglycans (GAGs/PGs). Chondroitin sulfate proteoglycans (CSPGs) participate in the modulation of various cellular functions, including adhesion and migration. The role of chondroitin sulfate (CS) chains on adhesion, chemotaxis and migration of poorly differentiated fibrosarcoma B6FS cell was studied, utilizing exogenous CS treatment and chondroitinase digestions as well as specific modulators of CS synthesis. Cleavage of cell-associated CS chains and specific inhibition of endogenous CS production severely impaired these fibrosarcoma cell functions. These results show that the reduction of endogenous CSPG expression as well as cleavage of the CS chain inhibited fibrosarcoma cell motility, migration and adhesion. Treatment with free CS chains enhanced cell chemotaxis and migration, whereas adhesion was inhibited. CS chains were found to upregulate cell motility through the MAPK pathway, specifically through JNK, whereas CS-induced migration was found to require tyrosine kinase dependent pathways. This study suggests a new role of CS on tumor cell adhesion, chemotaxis and migration.
The interactions between cells and the surrounding extracellular matrix (ECM) play crucial roles in various processes such as cell adhesion, migration and tumor growth. Cellular adhesion to ECM is a complex, tightly regulated process in which glycosaminoglycans/proteoglycans (GAGs/PGs) can participate in the formation of the specific adhesion sites. Cell motility and migration are cycling processes that require continuous assembly and disassembly of these specific cell matrix adhesions (1). Matrix adhesion signals activate specific downstream pathways, including a number of signaling molecules such as Rho, MEK, JNK, p38, FAK and Scr, which regulate assembly and disassembly of the actin cytoskeleton and the associated integrin adhesion complexes, thus affecting the ability of the cells to adhere (2, 3).
GAGs are present in all mammalian organs and tissues (4). The major GAG constituent of human serum is chondroitin sulfate (CS), accounting for 80% of the total serum GAG content (5-7). The microenvironment of the soft tissue tumor, fibrosarcoma, is rich in ECM components and particularly in GAGs/PGs, although the expression, cellular distribution and the turnover of PGs were determined to differ among cell lines of fibroblastic origin (8). Studies evaluating malignant cell motility link cell surface chondroitin sulfate proteoglycans (CSPG) with cell adhesion and migration. Melanoma cells synthesize increased levels of CSPGs on cell surface microdomains called microspikes, facilitating the initial contact of the cell to the ECM (9). Furthermore, CSPGs are suggested to coordinate the regulation of the integrin function in cell adhesion and migration (9-12). Recent studies propose that CSPGs may regulate cell activation by affecting different signal transduction pathways (9, 11, 13-15). It has been previously demonstrated that CS chains stimulate fibrosarcoma cell proliferation through tyrosine kinase-dependent signaling pathways (16), while normal fibroblast proliferation, motility and chemotaxis were inhibited (17).
The purpose of the present study was to examine the effects and the main downstream pathways of both endogenously synthesized and exogenously added chondroitin sulfate on fibrosarcoma cell adhesion, motility and migration. The results show that the reduction of endogenous CSPGs expression as well as cleavage of the CS chain inhibited fibrosarcoma cell motility, migration and adhesion. Treatment with exogenously added CS chains dose-dependently stimulate fibrosarcoma cell motility and migration through JNK and tyrosine kinase dependent pathways, introducing a new role of CS chains on these tumor cell functions.
Materials and Methods
Materials. Chondroitin sulfate type A (CSA), sodium chlorate, β-D-xyloside and genistein were obtained from Sigma Chemical Co. (St Louis, MO, USA). Chondroitinase AC was purchased from Seikagaku Kogyo Co (Tokyo, Japan). Cell culture reagents were obtained from Gibco, USA.
Cell culture. B6FS human fibrosarcoma cells (18) were grown in RPMI-Glutamax (RPMI) (Gibco, USA) supplemented with 10% FBS and gentamicine (0.04 mg/ml; Gibco, USA) at 37°C and 5% CO2. The cells were starved for 24 hours in serum-free RPMI. Before all assays treatments the cells were pretreated respectively with chondroitinase AC (0.1 U/ml; Seikagaku, Japan) used for cell-associated CSA removal; heat-inactivated chondroitinase AC (AC) (0.1 U/ml) used as a control; sodium chlorate (25 mM; Sigma) used as an inhibitor of proteoglycan sulfation; β-D-xyloside (50 μg/ml; Sigma) used as an inhibitor for CSPG biosynthesis; SP600125 (50 mM; Sigma) used as JNK inhibitor; U0126 (30 mM; Cell Signalling Technology, USA) used as MEK1/2 inhibitor; SB-230580 (5 mM; CalBiochem, Germany) used as p38 inhibitor; and genistein (10 μg/ml Sigma) used as tyrosine kinase inhibitor for 1 hour.
Cell adhesion. Human fibrosarcoma cells were cultured in RPMI supplemented with 10 % FBS in a 75 cm flask. The cells were starved for 24 hours in serum-free RPMI. After serum starvation, B6FS cells were pretreated as described in cell culture section during 1 hour. Cells were detached using PBS/EDTA 5 mM and 5,000 cells were seeded in a 96-well plate in the presence of chondroitinase AC and AC- (0.1 U/ml), sodium chlorate (25 mM) and β-D-xyloside (50 μg/ml) and CSA (30 μg/ml). After 2 min incubation time, the floating cells were removed with PBS and adhered cells dislodged using Cell Detachment Solution (Chemicon, USA), lysed using Cell Lysis Buffer (Chemicon) and their cell number measured using CyQuant GR Dye (Invitrogen, USA) on a fluorometer (Biotek) using 480/520 nm filter set. The experiments were performed in triplicate.
Wound healing. Human fibrosarcoma cells were seeded in 24-well culture plates and cultured in RPMI supplemented with 10% FBS until confluent. The cells were starved for 24 hours in serum-free RPMI. After serum starvation B6FS cells were pretreated as described in the cell culture section during 1 hour. The pretreated cell layer was wounded by scratching with a sterile 10 μl pipette tip. Detached cells were removed by washing two times with serum-free media. The cell motility was monitored during 6 hours in the presence of CSA (30 μg/ml), chondroitinase AC and AC- (0.1 U/ml), sodium chlorate (25 mM), β-D-xyloside (50 μg/ml) SP600125 (50 mM), U0126 (30 mM), SB-230580 (5 mM), and genistein (10 μg/ml). The wound closure was monitored using a digital image processor connected to a microscope, at six different positions across the wound. The cell motility was quantified by image analysis (ImageJ 1.4.3.67; Launcher Symmetry Software). The experiments were performed in triplicate.
Chemotaxis assay. B6FS cell chemotactic migration was assessed using 24-transwell plates (Corning Inc., USA). After serum starvation, B6FS cells were pretreated as described in the cell culture section during 1 hour. The pretreated cells were harvested with PBS/EDTA (5 mM). A total of 50,000 B6FS cells were loaded into the upper well of the chamber with serum-free media in the presence of chondroitinase AC and AC- (0.1 U/ml), sodium chlorate (25 mM), β-D-xyloside (50 μg/ml), SP600125 (50 mM), U0126 (30 mM), SB-230580 (5 mM), and genistein (10 μg/ml). Chemoatractant (CSA, 100 μg/ml) in 0% RPMI was placed in the bottom chamber. After 6 hours' incubation time, the migrated cells under the bottom of the insert were dislodged using Cell Detachment Solution (Chemicon), lysed using Cell Lysis Buffer (Chemicon) and their cell number measured using CyQuant GR Dye (Invitrogen) on a fluorometer (Biotek) using a 480/520 nm filter set. The experiments were performed in triplicate.
Statistical analysis. The statistical significance was evaluated using the t-test and the one way completely randomized variance analysis (ANOVA) using Microcal Origin (version 5.0) software.
Results
Effect of endogenous chondroitin sulfate chains on fibrosarcoma cell adhesion, chemotaxis and migration. The human B6FS fibrosarcoma cell line, predominantly consisting of fibroblastoid cells (B6FS) with a high metastatic capacity was utilized in this study (18). Sodium chlorate (SC), a known inhibitor of GAG sulfation, is used to effectively reduce the sulfation of all PG classes, resulting in the production of undersulfated proteoglycans whose secretion is partially prevented by the quality control mechanisms of the cell. SC acts by competing with sulfate in synthesis of 3′-phosphoadenosine 5′-phosphosulfate, the sulfate donor for glycosaminoglycan sulfation, with no significant effect on glycosaminoglycan and protein synthesis or on cell viability (19). As shown in Figure 1, the reduction of total PG sulphation inhibited the basal level of fibrosarcoma cell adhesion, chemotaxis and migration. In order to investigate the specific role of CSPGs on the tested fibrosarcoma cell functions, β-D-xyloside was utilized. β-D-Xyloside acts by competitively disabling the coupling of CS chains to the core protein which inhibits CSPG biosynthesis, while simultaneously enhancing the secretion of low molecular weight CS (20). The results demonstrate that the reduced expression of cell-associated CSPGs after β-D-xyloside treatment severely inhibited the basal level of fibrosarcoma cell adhesion, chemotaxis and motility. The reduction of CSPGs by β-D-xyloside had a similar effect on the basal level of fibrosarcoma cell adhesion, motility and chemotactic migration as compared to that caused by the reduction of total sulphated PG levels, suggesting that CSPGs are the main class of PGs that regulate these fibrosarcoma cell functions (Figure 1A, B and C).
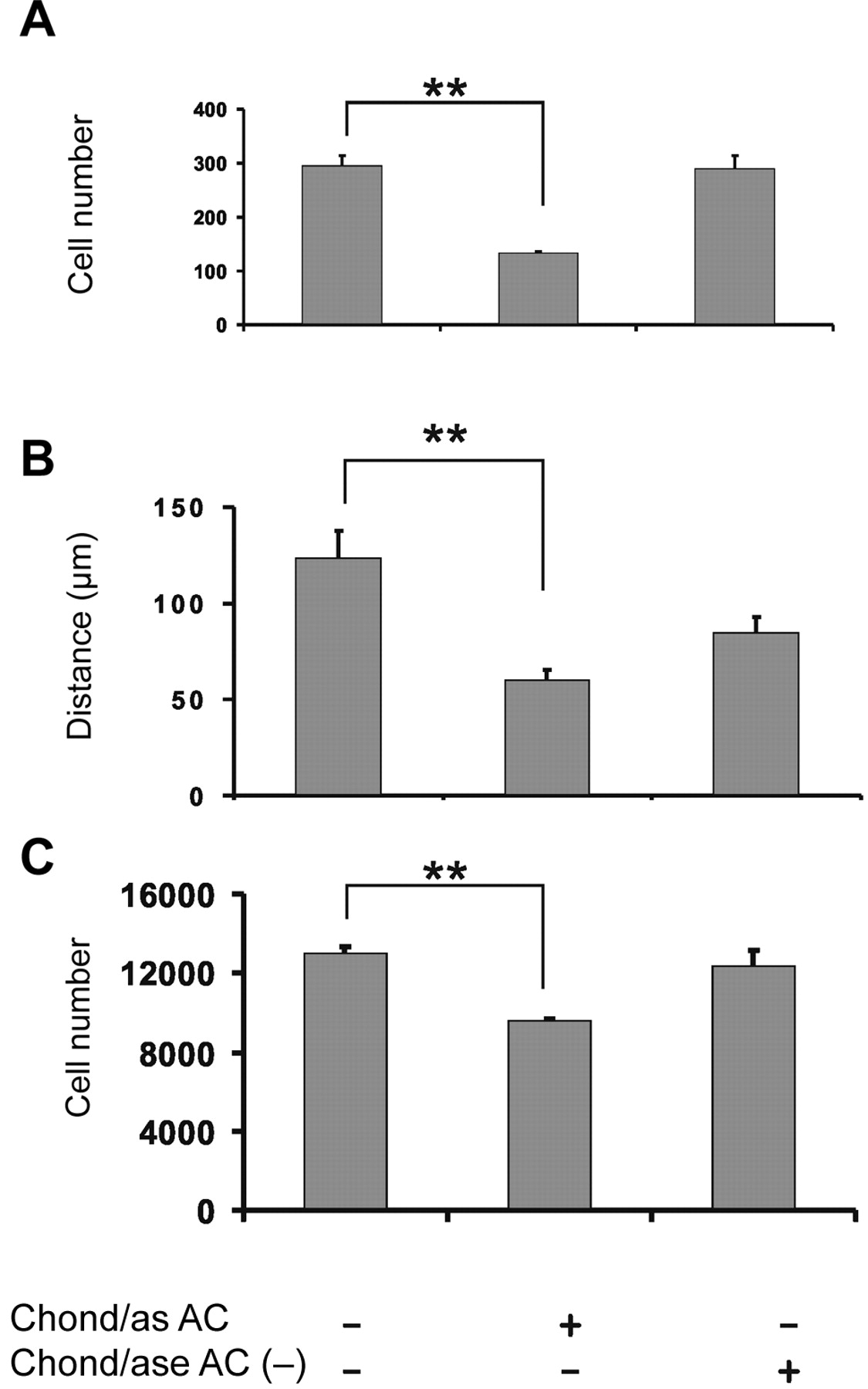
To further investigate the possible participation of cell-associated CS chains on these fibrosarcoma cell functions, B6FS cells were treated with chondroitinase AC. The enzymic treatment of the cells with chondroitinase AC targeted the removal of surface bound CS (A and C) chains. The removal of cell-associated CS chains had a severely inhibitory effect on fibrosarcoma cell adhesion, chemotaxis and motility (Figure 2A, B and C).

Inhibition of endogenous CS chain synthesis. Serum-starved B6FS cells were treated with sodium chlorate (25 mM) and β-D xyloside (50 μg/ml). B6FS cell adhesion (A), motility (B) and migration (C) were monitored. Means±S.E.M. are plotted; n=5. Statistical significance: **p<0.001.
Exogenous CS chains regulate fibrosarcoma cell adhesion, chemotaxis and migration. In order to evaluate the specific role of the exogenous CS chains on the migratory capabilities of fibrosarcoma cells, B6FS cells were treated with 10-100 μg/ml of purified CS. The CSA used in this study was obtained from Sigma and has previously been subjected to an extensive biochemical characterization (21, 22). The utilized concentration range was selected according to data from previous studies which had demonstrated that the CS concentration in human blood serum ranges between 5-30 μg/ml (23). Exogenous CSA chains altered fibrosarcoma cell adhesion, motility and chemotaxis at high concentrations (30 and 100 μg/ml) (Figure 3A). Cell motility (Figure 3B) and chemotaxis (Figure 3C) were enhanced by CS chains, while cell adhesion (Figure 3A) was inhibited.

Selective removal of surface bound CS chains. Serum-starved B6FS cells were treated with chondroitinase AC (0.1 U/ml) and heat-inactivated AC-. B6FS cell adhesion (A), motility (B) and migration (C) were monitored. Means ± S.E.M. are plotted; n=5. Statistical significance: **p<0.001.
Exogenous CS chains regulate cell motility mainly through JNK signaling pathway, whereas chemotaxis is controlled mainly through tyrosine kinase-dependent signaling pathways. To elucidate the signaling pathways that mediate the CS-induced fibrosarcoma cell motility and migration, inhibitors of the main cellular signaling pathways which regulate actin cytoskeleton rearrangements were utilized. There is accumulating evidence that mitogen activated-protein kinases (MAPK), including Jun N-terminus kinase (JNK), MEK kinase and p38 play important roles in cell spreading, laminopodia extension and tail retraction during cell migration. JNK inhibitor (SP600125), MEK1/2 inhibitor (UO126) and p38 inhibitor (SB-230580) were utilized. Moreover, a general tyrosine kinase (PTK) inhibitor, genistein, was used in order to examine the possible implication of CS chains in tyrosine kinase initiated signaling pathways (24, 25). The results demonstrated that CS chains upregulates cell motility through the MAPK pathway, mainly through JNK (Figure 4A) but not MEK1/2, whereas CS-induced cell migration requires tyrosine kinase-dependent pathways (Figure 4B).

Effect of exogenous CS chains on B6FS human fibrosarcoma cell adhesion (A) motility (B) and chemotactic migration (C). B6FS cells were starved overnight and treated with different CS (1, 10, 30, 100 μg/mL) concentrations for 2 min in the case of cell adhesion, and 6 hours for both cell motility and migration. Means±S.E.M. plotted Means±S.E.M. plotted; n=5. Statistical significance: *p<0.01, **p<0.001; ns: non-significant.

Effect of main signaling pathways inhibitors. Serum starved B6FS cells were treated with SP600125 (5 mM), UO126 (30 mM), SB-230580 (5 mM) and genistein (10 μg/ml). Cell motility (A) and migration (B) were measured after 6 hours of treatment. Means±S.E.M. are plotted n=5. Statistical significance: **p<0.001.
Discussion
The complex interactions of cells with ECM components play a significant role in regulating signaling pathways that contribute to tumor progression. Previous reports correlate cell surface CSPGs with tumor cell adhesion, motility and migration (2, 10), however the data regarding the role of CS chains on these fibrosarcoma cellular functions are limited. The present study demonstrates a dose-dependent effect of CS chains on fibrosarcoma cell motility and migration which is mediated by JNK and tyrosine kinase-dependent signaling pathways.
During cell adhesion, information from the extracellular to the intracellular compartments is mainly conveyed through the integrin family members (26). Integrin-mediated cell adhesion and/or migration results in the recruiting of the focal adhesion kinase (FAK) to the focal adhesion points and its subsequent tyrosine phoshorylation (27). Various studies have demonstrated the obligatory role of membrane CSPGs in the formation of focal adhesion complexes (28, 29). Melanoma cells during cytoskeleton rearrangements synthesize increased levels of CSPGs, located on microspikes at the cell surface (9). CD44, a chondroitin sulfate PG, is suggested to be involved on endothelial and melanoma cell migration, while neurocan and NG2 are proposed to play important roles in migration and glial-neuron interactions (10, 12, 30, 31).
Previous studies utilizing inhibitors or enzymes which degrade CS chains indicate that CS plays an important role in the processes of tumor growth and metastasis including migration and adhesion (14, 15, 32). In this study, the reduction of endogenous CSPG or the removal of cell-bound CS chains showed an inhibitory effect on fibrosarcoma cell adhesion, motility and chemotaxis, indicating a critical role of CSPGs/CS chains on these cellular functions. Similarly with the presented results, chondroitin sulfate A and C were found to specifically regulate integrin-mediated cell adhesion (11, 13, 30). It has also been reported that chondroitinase AC exerts antitumor effects by inhibiting melanoma and endothelial cell migration and invasion (13).
To elucidate the functions of matrix CS chains on fibrosarcoma cell adhesion, motility and migration, B6FS cells were treated with exogenously added purified CS chains. CS chains enhanced B6FS cell motility and migration, while their cell adhesion was inhibited in a dose-dependent manner. These effects were not observed after the xyloside treatment, which is known to promote a secretion of CS chains, probably due to insufficient amounts, as compared to the 30-100 μg/ml exogenously added CS. A central role of CS in α4β1 integrin-mediated cell migration through the enhancement of FAK phosphorylation, has been suggested (33). Thus, the observed dose-dependent stimulatory effect of free CS chains on fibrosarcoma cell motility and chemotactic migration might be due to the participation of CS in an integrin-dependent signaling cascade. Recent studies on cellular adhesion to plastic have proposed a sulfation pattern-specific role for CS, showing that chondroitin-6-sulfate increases while chondroitin-4-sulfate reduces cellular adhesion (34). Our results demonstrate a dose-dependent inhibitory effect of free CS chains, characterized to be mostly chondroitin-4-sulfate (21), on fibrosarcoma cell adhesion, which is in agreement with studies on palatal fibroblasts and leukemia cells (34, 35). The observed inhibitory action of free CS chains on fibrosarcoma cell adhesion might be due to the competition with the corresponding cell bound CS for the available binding sites on the plastic surface.
To clarify the signaling pathways that contribute to CS-induced fibrosarcoma cell motility and migration, inhibitors of the main cellular signaling pathways which regulate actin cytoskeleton rearrangements were utilized. It has been suggested that mitogen activated-protein kinases (MAPK), including Jun N-terminus kinase (JNK), MEK kinase and p38 play crucial roles in cell spreading, laminopodia extension and tail retraction during cell migration (36-40). JNK inhibitor (SP600125), MEK1/2 inhibitor (UO126) and p38 inhibitor (SB-230580) were utilized. The presented results demonstrated that CS chains stimulate cell motility through the MAPK pathway, and more specifically, mainly through JNK. Recent studies have emphasized the general role of MAPK and have especially clarified the role of JNK on motility (2, 41). It has been shown that the signaling molecules that activate JNK are essential for cell motility (3, 38, 41). Morever, inhibition of JNK by either the chemical inhibitor SP600125 or the dominant negative mutant JNK1AF, significantly impairs the rate of motility in melanoma cells (2,41).
Focal adhesion formation at the cytoplasmic face of the cell membrane requires the connection of the adaptor proteins to the actin cytoskeleton as well as of integrins with receptor tyrosine kinases (42). Different MAPK pathways may be initiated by tyrosine kinases receptors (42). In the present study, a general tyrosine kinase (PTK) inhibitor, genistein, was used in order to examine the possible implication of CS chains in tyrosine kinase-dependent signaling pathways. Genistein, a protein tyrosine kinase (PTK) inhibitor is suggested to have an inhibitory effect in carcinogenesis (43), cell invasion and metastasis (43-45). Both random and CS-induced fibrosarcoma cell migration was severely inhibited by the action of genistein, suggesting the role of tyrosine kinases in fibrosarcoma cell migration.
The co-stimulatory effect of CS chains on PDGF-induced fibrosarcoma cell proliferation through tyrosine kinase-dependent pathways has previously been demonstrated, suggesting a new role of CS in activating signaling pathways that affect fibrosarcoma tumor growth (16). In the present study, the dose-dependent effect of free CS chains on fibrosarcoma cell motility and chemotaxis mediated by JNK and tyrosine kinase-dependent signaling pathways was elucidated. Further investigation is needed in order to clarify the mechanisms of CS action in fibrosarcoma tumor growth.
Acknowledgements
This project was financially supported by the Greek ministry of Education (25%) and European Community (75%) programme EPEAK II, PYTHAGORAS II, Grant Number: KA2089.
- Received July 18, 2008.
- Revision received November 28, 2008.
- Accepted December 2, 2008.
- Copyright © 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}