


Abstract
Background/Aim: The mouse vagina exhibits stratified squamous epithelium, which is comprised of multiple cell layers. We previously showed that erbB signaling, induced by epithelial estrogen receptor 1 (ESR1), is required for the initial differentiation of the epithelium. However, the downstream effector that mediates terminal differentiation in the apical layers remains elusive. The contribution of fibroblast growth factor (FGF) to vaginal epithelial cell differentiation was investigated. Materials and Methods: Vaginas from wild-type or epithelium-specific Esr1 conditional knockout (cKO) mice were analyzed using immunohistochemistry and quantitative real-time RT-PCR. Results: Of the FGF ligands examined, Fgf22 mRNA was significantly induced following estrogen treatment. Furthermore, FGF downstream signaling, phosphorylated FRS2 and ERK1/2 were exclusively expressed in the apical layers of the vaginal epithelium. No changes in such expression were observed in the Esr1 cKO mice. Conclusion: FGF-ERK/MAPK pathway may be a main inducer of terminal differentiation in the mouse vaginal epithelium.
Estrogens play important roles in regulating mammalian organ homeostasis. In particular, cell proliferation and differentiation of female reproductive organs, such as mammary gland, uterus and vagina, are tightly regulated by estrogens (1). In vagina, estrogens induce epithelial cell proliferation in the basal layer (2). The suprabasal cells are no longer mitogenic and differentiate while moving up through the epithelium (2). The apical cells exhibit keratinization, and they lose their nuclei and cytoplasm, which are eventually shed from the surface (2). The fully stratified and keratinized epithelium in the vagina resembles epidermis in the skin; however, unique and comprehensive process of this epithelial cell differentiation in mouse vagina, which is again regulated by estrogens, remains elusive.
Estrogens activate their target gene expression mainly through binding to the nuclear estrogen receptors α and β (ESR1 and ESR2) (1). Classical tissue recombination experiments have shown that epithelial cell proliferation is mediated through stromal ESR1, whereas epithelial cell differentiation requires epithelial ESR1 (3-5). Our in vivo study with vaginal epithelium-specific Esr1 cKO mice has also revealed that epithelial ESR1 is required for cell differentiation; the vagina of cKO mice did not express cytokeratin 1 (CK1) and filaggrin, early and terminal differentiation maker proteins, respectively (6). We also found that amphiregulin (Areg) is a mediator for CK1 expression in mouse vagina in response to estrogen. Areg is, however, insufficient for full expression of filaggrin and terminal differentiation (6). Therefore, additional downstream factors are necessary for full activation of estrogen effects and terminal differentiation in the epithelium of the mouse vagina.
Fibroblast growth factor (FGF) plays crucial roles for embryonic development and tissue homeostasis by regulating cell proliferation, migration and differentiation (7). The FGF family is composed of 22 secreted proteins, which are divided into 7 subfamilies (7). They interact with tyrosine kinase FGF receptors (FGFRs). Binding of FGFs facilitates dimerization of FGFRs and intracellular conformational changes, which activate tyrosine kinase domains and phosphorylates intermediate docking proteins, such as FGFR substrate 2 (FRS2) (8-11). The phosphorylated FRS2 (pFRS2) recruits the adaptor protein, growth factor receptor-bound 2 (GRB2), and the exchange factor, son of sevenless (SOS) (8-11). As a result, SOS activates the RAS-GTPase, which induces activation of the mitogen-activated protein kinase (MAPK) pathway, including the phosphorylation of extracellular signal-regulated kinases 1 and 2 (pERK1/2) (8-11). During the mouse vaginal development, the FGF-ERK/MAPK pathway is one of the key mediators of lineage determination of the neonatal Müllerian duct epithelium to the vaginal epithelium (12-14); however, the contribution of FGF signaling to the homeostasis of adult vaginal epithelium is still unclear.
Sequences of gene primer's for qRT-PCR.
In this study, we focused on the FGF signaling in order to understand the molecular pathway of the estrogen-mediated cell differentiation in the adult mouse vagina. Our results suggest that the activation of FGF-ERK/MAPK pathway is involved in a series of ESR1-mediating events, particularly in the terminal differentiation of the vaginal epithelium.
Materials and Methods
Mouse and treatment. All experimental procedures and protocols were approved by the Animal Research Committee of the Wakayama Medical University (Approval Number: 28-2 and 28-9). The mouse strains used were C57BL/6J (CLEA, Tokyo, Japan), K5-Cre (15), Esr1-null and Esr1-floxed (16). All lines were maintained in the C57BL/6J background. K5-Cre mice and Esr1-null mice were crossed to obtain male K5-Cre/+; Esr1+/− mice. Esr1 cKO mice (K5-Cre/+; Esr1flox/−) were obtained by crossing K5-Cre/+; Esr1+/− male mice with Esr1flox/flox female mice. For control mice, K5Cre/−; Esr1flox/+ littermates were used. Mice were ovariectomized (OVX) at 6 weeks of age and were culled at 8 weeks of age. To test the effects of estrogen, a single daily injection of 0.1 μg 17β-estradiol (E2, Sigma, St Louis, MO, USA) dissolved in sesame oil (Kanto Chemical, Tokyo, Japan) was given to OVX mice for 3 days and sacrificed 24 hours following the last injection. Unless otherwise mentioned, materials were purchased from Wako Pure Chemical, Osaka, Japan.
Immunohistochemistry. Eight weeks mice vaginas were fixed overnight in 4% paraformaldehyde (Sigma) with phosphate-buffered saline (PBS), embedded in paraffin, and sectioned on 8 μm. Antigen retrieval was performed by incubating with 0.1 mM citrate buffer (pH 6.0) in a microwave (200 W) for 10 min. Endogenous peroxidase activity was inactivated in methanol containing 0.3% H2O2 for 30 minutes. Following 1 hour incubation with 1.5% fetal bovine serum (FBS, Thermo Fisher Scientific, Waltham, MA, USA) in PBS as a blocking buffer, the following primary antibodies were used on the slides: anti-pERK1/2 (1:200, Cell Signaling, Danvers, MA, USA), anti-pFRS2 (1:200, R&D Systems, Minneapolis, MN, USA) and anti-pAKT (1:200, D9E, Cell Signaling). The sections were incubated with the primary antibody diluted in blocking buffer at 4°C for overnight. Final visualization was performed with the Vectastain ABC Kit (Vector Laboratories, Burlingame, CA, USA).
Quantitative real-time RT-PCR (qRT-PCR). Gene expression changes were quantified using the StepOnePlus Real-Time PCR System (Thermo Fisher Scientific), according to the manufacturer's instructions. Total RNA (2 μg) was isolated from whole mouse vaginas at 8 weeks of age using an Isogen RNA extraction kit (Nippongene, Toyama, Japan). Synthesis of cDNA was performed with SuperScript III reverse transcriptase (Thermo Fisher Scientific) and the resulting cDNA was mixed with the SYBR premix Ex Taq (Takara, Ohtsu, Japan). Three pools of samples per group were assayed in triplicates, and three samples were assayed. The relative RNA equivalents for each sample were determined by a comparison to the levels of the normalized standard ribosomal protein L8 (L8), and then calculated as the fold change compared to the normalized standard. The sequences of gene primers are shown in Table I (17, 18). Statistical analysis was performed by Tukey-Kramer test followed by two-way ANOVA. Differences with p<0.05 were considered significant. Error bars represent the standard error of the mean (SEM).
Results
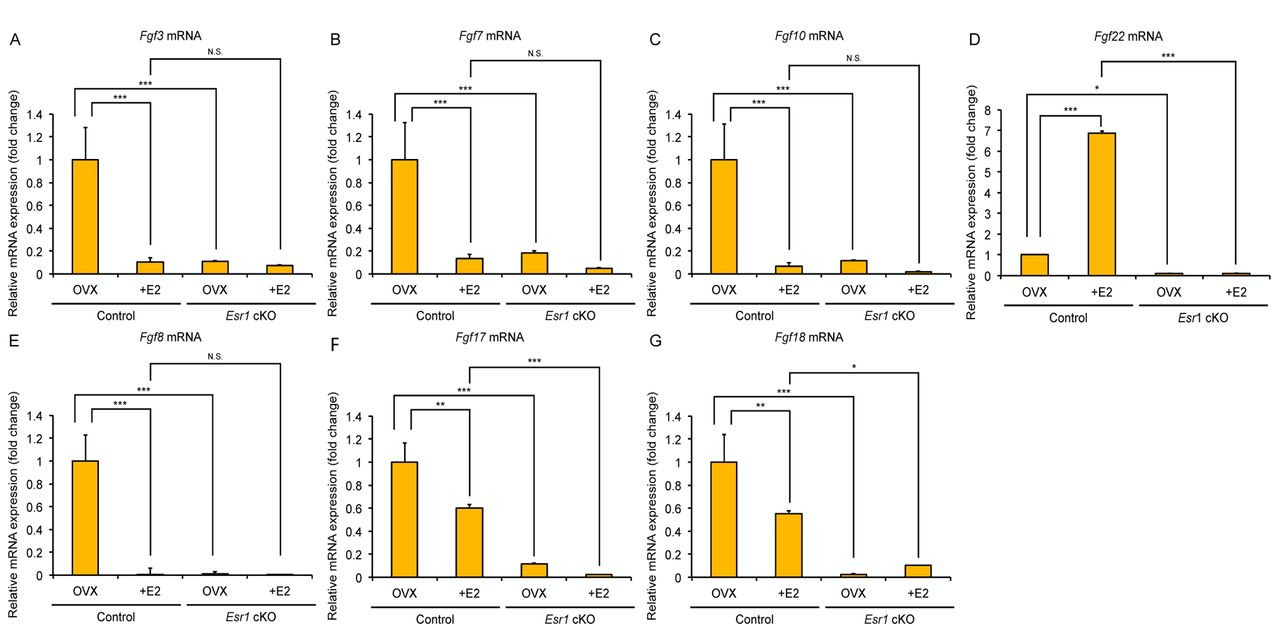
Expression patterns of Fgf ligands in the adult vagina. The signaling mechanism mediated through secreted growth factors, enables the amplification and systematic coordination of estrogen-induced epithelial cell proliferation and differentiation in the female reproductive organs. In this study, we focused on the FGF signaling, one of the major families of secreted growth factors. We have previously found that Fgf22 and Fgf18 expression are significantly decreased in the vagina of Esr1 cKO mice, which display failure of keratinized differentiation, suggesting that these FGF members are possibly involved in the epithelial cell differentiation (6). We first performed quantitative real-time RT-PCR (qRT-PCR) analysis for several FGF ligands expression in the Esr1 cKO mouse vagina. To remove any confounding biases of the hypothalamic-pituitary-gonadal axis and to simplify the analysis of estrogen effects, mice were ovariectomized and treated with 17β-estradiol (E2) for 3 days, which provides sufficient time for inducing stratified and adequately keratinized epithelium in the mouse vagina (6, 19). We tested several Fgf ligands: i) Fgf22, ii) Fgf3, iii) Fgf7, iv) Fgf10 (FGF7 subfamily), v) Fgf18, vi) Fgf8, vii) Fgf17 (FGF8 subfamily). In the control mice, E2-treatment led to the downregulation of most Fgfs mRNA expression, except for Fgf22, which was significantly augmented upon E2 stimulation (Figure 1 and Table II). In the Esr1 cKO mice, such upregulation was not observed even following E2 treatment.

Expression profile of FGF ligands in control and Esr1 cKO mouse vagina. Expression of Fgf3 (A), Fgf7 (B), Fgf10 (C), Fgf22 (D), Fgf8 (E), Fgf17 (F), Fgf18 (G) in mouse vagina are measured with quantitative real-time RT-PCR. Of these ligands, Fgf22 mRNA is significantly induced following E2 treatment in the control mouse. (*p<0.05, **p<0.01, ***p<0.001, N.S.: Not significant) (n=3).
FGF-MAPK signals were detected in the apical layer. We next examined the phosphorylation of FRS2 and ERK1/2, presumptive downstream targets of Fgf signaling. The vaginal epithelia of control and Esr1 cKO ovariectomized (OVX) mice were form two or three cell layers (Figure 2A and B). In control mice, E2 treatment induced epithelial stratification and keratinization, and pFRS2 was detected in the apical layer of the vaginal epithelium (Figure 2C). By contrast, vaginal epithelium of Esr1 cKO mice failed in keratinization and did not express pFRS2 (Figure 2D). Similar to pFRS, pERK1/2 expression was detected in the apical layers of epithelium in E2-treated control vaginas, but not in the Esr1 cKO vaginas (Figure 2E, F, G and H).
It has been reported that FGF and erbB signaling activate other cellular signal transduction pathway, such as phosphatidylinositol 3-kinase (PI3K) and AKT (20-26). Phosphorylated-AKT (pAKT) was expressed in the suprabasal (middle) layer rather than in apical layers in response to estrogen (Figure 2I, J, K and L), suggesting that PI3K/AKT signaling is not a main inducer of terminal differentiation in mouse vagina. These data also suggest that FGF-ERK/MAPK pathway activated by ESR1 mainly regulates the vaginal epithelial terminal differentiation.
Discussion
Estrogen tightly regulates cell proliferation and differentiation in the female reproductive organs. Previous tissue recombination experiments and conditional knockout mice studies have revealed that these events are mediated through complicated tissue-tissue interactions (3-6, 19). As in the case of many stratified epithelia, vaginal epithelium is maintained through the proliferation of basal cells, which requires stromal ESR1 expression and secretion of the proliferative factors (3-5). By contrast, epithelial cell differentiation is directly mediated by epithelial ESR1 (6). Suprabasal cells exit the cell cycle and enter a squamous cell differentiation processes, however, in the absence of epithelial ESR1, keratinized differentiation fails (6). In the mouse vaginal epithelium, interactions among multiple cell layers coordinated through various secretory signaling factors regulate epithelial cell homeostasis (6, 19). This is essential for the amplification of estrogen stimulation in the female reproductive organs. We have found that the erbB ligand, AREG, is expressed specifically in the epithelium, and is dependent on epithelial ESR1 expression (6, 17). Organ culture experiments have revealed that AREG compensates for the loss of epithelial ESR1 and induces CK1 expression, an early cell differentiation marker, even in the absence of estrogens. Nevertheless, AREG is insufficient for the full expression of filaggrin and terminal differentiation (6). Thus, other factors are likely to be involved in further epithelial cell differentiation.
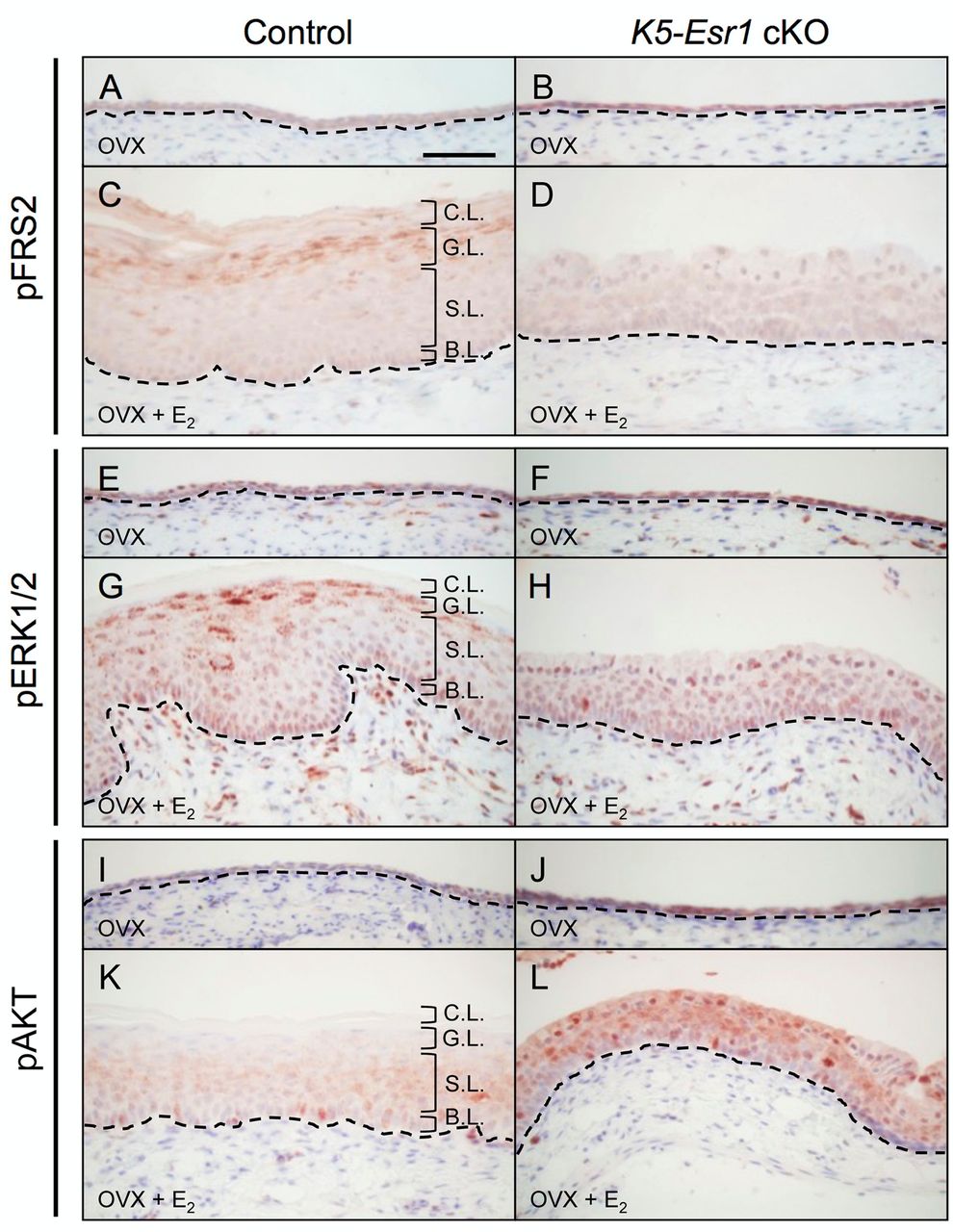
Expression pattern of FGF-MAPK and AKT signaling proteins in control and Esr1 cKO mouse vagina. The vaginal epithelia from OVX control mice show two or three cell layers (A, B, E, F, I and J). Expression of pFRS2 and pERK1/2 are detected in E2-treated vaginal epithelia of control mice, particularly in the apical layer (C and G). The vaginal epithelia from Esr1 cKO mice show failure of cell differentiation, stratification and phosphorylation of pFRS2 and pERK1/2 (D and H). pAKT is expressed in the middle layer of E2-treated vaginal epithelia of control mice and is augmented in the whole epithelium of E2-treated Esr1 cKO vagina (K and L). Dotted lines indicate the basal layer of the epithelium. Scale bar=100 μm. (n=3). B.L.: Basal Layer; S.L.: Spinous Layer (Suprabasal Layer); G.L.: Granular Layer (Apical Layer); C.L.: Cornified Layer.
The results of qRT-PCR for Fgf ligands.
In this study, we found that FGF signaling is a possible candidate involved in the terminal differentiation in mouse vagina. Of the Fgf ligands examined, Fgf22 was unique to be induced following E2 treatment in the control mouse vagina. FGF22 is classified into the same group as FGF7 (7), and is called keratinocyte growth factor (KGF), regulating the cell differentiation and regeneration of various epithelial tissues, such as esophagus, lung, trachea and skin (27-32). KGF treatment in the neonatal or adult mouse vagina results in fully stratified and keratinized epithelium, suggesting a relationship between estrogen signaling and activation of FGF signaling (33-35). KGF and FGF22 bind to FGFR2IIIb (36, 37), which is predominantly expressed in epithelial cells, and is essential for epithelial cell proliferation, differentiation and homeostasis (38-40). From these data, we hypothesized that Fgf22 is involved in the terminal differentiation of the mouse vagina as a downstream factor of estrogens, although it is unclear whether ESR1 directly regulates Fgf22 expression. By contrast, Fgf18 was not induced following E2 treatment in the current study. In the mouse uterus, it has been reported that FGF18 is induced by stromal ESR1 and possibly mediates epithelial cell proliferation (41, 42). Thus, we cannot rule out a possible role of FGF18 on the vaginal epithelial cell homeostasis. Further studies are needed in order to elucidate the regulatory mechanisms of FGF signaling by estrogens in the female reproductive tracts with a clear view of the differences between uterus and vagina.
pFRS2 and pERK1/2 were co-expressed in the apical layers of mouse vagina, suggesting that the FGF-ERK/MAPK pathway can be a main inducer of terminal differentiation. By contrast, pAKT was detected in the suprabasal layers of the mouse vagina, where early differentiating cells are localized. Thus, the PI3K/AKT pathway is possibly activated by erbB signaling in the mouse vagina (22-26). Akt phosphorylated by PI3K can lead to the stabilization of the cell cycle inhibitor, p21 (43). It is, thus, suggested that PI3K/AKT might promote cell differentiation by attenuating cell proliferation in the mouse vaginal epithelium. In fact, p21 is expressed in the suprabasal cells of the mouse vaginal epithelium upon estrogen treatment (44). Intriguingly, pAKT was found to be augmented in the whole epithelial layers of E2-treated Esr1 cKO vaginas. In these mice, the suprabasal layers of the epithelium have been shown to exhibit aberrant and ectopic cell proliferation, which never happens in the normal stratified epithelium (6). Considering the results, this augmented pAKT promotes ectopic cell proliferation in the whole epithelium of Esr1 cKO vagina.
Taken together, our results suggest that FGF-ERK/MAPK signaling is involved in the vaginal epithelial terminal differentiation. Overall, we unraveled the mechanism of estrogen signaling in the mouse vagina, which is key to the elucidation of estrogen-mediated events in the female reproductive organs, including disorders such as hormone-induced tumorigenesis.
Acknowledgements
The Authors thank Drs. P. Chambon and J. Takeda for providing Esr1-floxed and K5-Cre mice respectively, and Drs. D. Matsumaru, A.Murashima, D. Kajioka, R.Yatsu, K. Toyota for their intellectual input. They would also like to thank Mr. H. Saeki and Ms. T.I. Iba for their assistance. This work was supported by Japan Society for the Promotion of Science KAKENHI Grant Number 17H06432, 18H02474, 18K06837, 18K06938.
Footnotes
Authors' Contribution
Y.H., T.I. and S.M. designed the research. Y.H. and T.I. collected tissue samples and performed experiments. Y.H., K.S. and G.Y. analyzed data. Y.H. and S.M. supervised the research and wrote the manuscript.
This article is freely accessible online.
Conflicts of Interest
None of the Authors has any potential conflicts of interest associated with this research.
- Received May 26, 2019.
- Revision received June 30, 2019.
- Accepted July 3, 2019.
- Copyright© 2019, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.