


Abstract
Background/Aim: Bacterial resistance to antibiotics has become a serious problem in antibacterial chemotherapy and resistance of bacteria to chemically-unrelated anti-microbial agents can be associated with the over-expression of efflux pumps. The simultaneous therapy with efflux pump inhibitors (EPIs) could be a solution to improve the effectiveness of antibiotics. The response of an organism to an EPI often depends on how that molecule fits a particular site of a protein. Because enantiomers of a given compound rotate plane-polarized light in a solution by the same angle but in opposite directions, the rational drug design should take the chirality into account if there is a difference between the racemic compound and its enantiomers. Materials and Methods: The main goal of the present study was to elucidate the role of chirality of N-hydroxyalkyl-2-aminophenothiazines as effective EPIs by an automated method that uses the general efflux pump substrate ethidium bromide (EB) for the assessment of AcrAB-TolC system of wild-type Escherichia coli K-12 AG100. It has been shown that the most active EPIs among the N-hydroxyalkyl-2-aminophenothiazines were the compounds rac-3i, (+)-3i, and (−)-3i by modulating the AcrAB-TolC efflux pump. Conclusion: Comparison of effects of enantiomeric pairs revealed that their activities were similar to that of racemic derivatives. Moreover, there was no significant difference between the racemic compounds and their enantiomers related to their antibacterial and efflux pump inhibiting effects.
Drug resistance has evidently appeared in response to selective pressures resulting from the excessive use of antibiotics and other anti-microbials. Multidrug efflux systems are responsible for clinically important resistance to chemotherapeutic agents in pathogenic bacteria, fungi, parasites and in human cancer cells. Efflux mechanisms of bacteria, namely pumping of anti-microbial agents out of the cells play an important role in anti-microbial resistance of pathogenic bacteria. Bacterial anti-microbial efflux systems can be divided into five superfamily classes, the major facilitator (MF) superfamily, the ATP-binding cassette (ABC) superfamily, the resistance-nodulation-division (RND) superfamily, the small multidrug resistance (SMR) superfamily and the multidrug and toxic compound extrusion (MATE) superfamily (1, 2).
The use of efflux pump inhibitors (EPIs) has been investigated in order to improve the activity of antibiotics and the goal of the pharmacological chemistry has been the development of compounds that decrease the level of intrinsic resistance, significantly reverse acquired resistance and promote the decrease of emergence of multidrug resistant bacteria (3). Considering the main objective of using EPIs, these compounds should increase the susceptibility of bacteria to one or more antibiotics. The EPIs can address various efflux targets among which the followings can be emphasized: (i) expression of genes that induces multidrug resistance (MDR), (ii) assembly of membrane transporter complex responsible for drug efflux, (iii) energy involved in the activity of transporter proteins and (iv) inhibition of the movement of molecules inside the efflux channel by competition or blocking (4).
The rational drug design should consider the chirality of the molecules if there is a difference between the racemic compound and its enantiomers. In addition, it has been found that the S-enantiomer and R-enantiomer of thioridazine did not differentially affect the growth of Mycobacterium tuberculosis strains (5).
The role of chirality of EPIs was investigated in case of the enantiomers of tricyclic compounds, such as methotrimeprazine and clopenthixol, on the drug efflux of mouse T-lymphoma cells expressing the ABCB1 transporter (P-glycoprotein). It has been found that the stereoselectivity does not have any effect on the inhibition of the efflux pump activity of enantiomers (6). Similar findings have been obtained after the chemical resolution of N-hydroxyalkyl-2-aminophenothiazines, namely the enantiomers were shown to have similar activity on the inhibition of the ABCB1 transporter of rat hepatocytes (7).
The main question of this work was whether the chirality of resolved phenothiazine derivatives can play a role in the inhibition of the AcrAB-TolC efflux pump system of a wild-type Gram-negative bacterium of pathogenic interest. For this reason we used the AcrAB-TolC system of E. coli K-12 AG100 (wild-type, genetically defined strain) and racemic phenothiazines and their (+) −and (−)− enantiomers. Representative efflux pumps such as the E. coli AcrAB-TolC have been used to screen and characterize possible EPIs. An automated real-time screening method has been developed by Viveiros et al., which can be used routinely for the screening of EPIs (8) under physiologically relevant conditions (i.e. pH). This screening method was used to elucidate the activity of the different phenothiazine enantiomers on the bacterial AcrAB-TolC system.
Materials and Methods
Bacterial strains. Wild-type E. coli K-12 AG100 strain [argE3 thi-1 rpsL xyl mtl Δ(gal-uvrB) supE44] expressing the AcrAB-TolC efflux pump at its basal level. The strain was kindly provided by Hiroshi Nikaido (Department of Molecular and Cell Biology and Chemistry, University of California, Berkeley, CA, USA).
Materials. Mueller Hinton (MH) powder (Sigma-Aldrich, St. Louis, MO, USA) was used for the preparation of MH broth. E. coli cultures were grown on solid (1.5% agar) for isolation or liquid Luria Bertani (LB) medium purchased from Difco, Detroit, MI, USA. Ethidium bromide (EB) was purchased from Sigma-Aldrich.
Compounds (EPIs). The synthesis and chemical resolution of the phenothiazines derivatives were described previously (7); the structures are depicted in Table I.
4-[2-(4-Methoxyphenyl)-2H-tetrazol-5-yl]-1-(2-morpholin-4-yl-10H-phenothiazin-10-yl)butan-2-ol (rac-3h)
4-[2-(4-Methoxyphenyl)-2H-tetrazol-5-yl]-1-[2-(4-methylpiperazin-1-yl)-10H-phenothiazin-10-yl]butan-2-ol (rac-3i)
1-(2-Chloro-10H-phenothiazin-10-yl)-4-[2-(4-methoxyphenyl)-2H-tetrazol-5-yl]butan-2-ol (rac-5).
The structure and the chemical formula of the phenothiazines used in the study.
Minimum inhibitory concentration (MIC) determination. The MIC of phenothiazine derivatives was tested according to Clinical and Laboratory Standard Institute (CLSI) guidelines for dilution anti-microbial susceptibility tests (9). All assays were repeated three times.
Real-time EB accumulation assay. The activity of compounds on the real-time accumulation of EB was assessed by the automated EB method (8, 10), using a Rotor-Gene 3000™ thermocycler with real-time analysis software (Corbett Research; Sydney, Australia). Briefly, an aliquot of an overnight culture of the strain in LB medium was transferred to fresh LB medium and incubated until it reached an optical density (OD) of 0.6 at 600 nm, washed with phosphate buffered saline (PBS pH 7.4) and centrifuged at 13,000 × g for 3 minutes. The pellets were re-suspended in PBS (pH 7.4) and the OD adjusted to 0.6 at 600 nm. Aliquots of 45 μl of the cell suspension were distributed to 0.2 ml tubes. The compounds were added individually at the concentration of 100 μM in 5 μl volumes of their stock solutions and, finally, 45 μl of EB were added to yield a final concentration of 1 mg/l in PBS with and without glucose 0.4%. The tubes were placed into the Rotor Gene 3000™ thermocycler that monitors the excitation wavelength (535 nm) and the emission wavelength (585 nm). The fluorescence associated with retained EB was monitored on a real-time basis.
If an agent is to be studied for activity against an efflux pump system it must be applied at concentrations that do not affect the replication or viability of the cell (8). Because the MICs of the phenothiazine derivatives exceeded the concentration of 400 μM, the sub-inhibitory concentrations for the accumulation assays were employed at 100 μM.

The activity of the most effective N-hydroxyalkyl-2-aminophenothiazine derivatives (rac-3i, (+)-3i, (−)-3i) at 100 μM on the accumulation of EB by E. coli AG100, in the presence and absence of glucose 0.4%, after 30 min. Legend: — solvent control; ● rac-3i without glucose; ▴ (+)-3i without glucose; ▪ (−)-3i without glucose; --- solvent control with glucose; ○ rac-3i with glucose, ▵ (+)-3i with glucose, □ (−)-3i with glucose.
From the real-time data, the activity of the compound, namely the relative fluorescence index (RFI) of the last time point (minute 60) of the EB accumulation assay, was calculated according to the formula:
 where RFtreated is the relative fluorescence (RF) at the last time point of EB retention curve in the presence of an inhibitor; and RFuntreated is the relative fluorescence at the last time point of the EB retention curve of the untreated control having the solvent control (DMSO) (11). The greater the difference between RFtreated and RFuntreated the greater the degree of the EB accumulated and, therefore, the greater the degree of inhibition of the efflux pump system of the bacterium by the agent. The experiments were repeated three times and the RFI values presented are the average of three independent assays.
where RFtreated is the relative fluorescence (RF) at the last time point of EB retention curve in the presence of an inhibitor; and RFuntreated is the relative fluorescence at the last time point of the EB retention curve of the untreated control having the solvent control (DMSO) (11). The greater the difference between RFtreated and RFuntreated the greater the degree of the EB accumulated and, therefore, the greater the degree of inhibition of the efflux pump system of the bacterium by the agent. The experiments were repeated three times and the RFI values presented are the average of three independent assays.
Statistical analysis. Statistical analyses were conducted using a one-way analysis of variance (ANOVA) complemented by the Bonferroni post-test for multiple comparisons. The level of significance was p<0.05. The analyses were performed using the GraphPad Prism version 5.00 for Windows, GraphPad Software (San Diego, CA, USA, www.graphpad.com).
Results
The phenothiazine derivatives had no prominent antibacterial effect on E. coli AG100 expressing the AcrAB-TolC efflux pump, the MICs were above 400 μM and the DMSO as solvent had no antibacterial effect at the concentration applied in the assays (5 % v/v). There was no difference between the (+)- and (−)-enantiomers and the racemic compound in their antibacterial effect; all of them had an MIC above the concentration of 400 μM.
After determining the MICs, the RFI was calculated for each derivative. As shown in Figure 1, the most active EPIs for the modulation of the AcrAB-TolC pump were compounds rac-3i, (+)-3i, and (−)-3i.
The difference between the enantiomers in terms of RFI was evaluated by one-way analysis of variance (ANOVA) with Bonferroni post-test for comparing all pairs of results by using the GraphPad Prism software. A 95% confidence interval has been chosen for the analysis and the means of the triplicate assays were compared. Comparison of RFI values for the enantiomeric derivatives revealed that their activities were comparable to that of the racemic derivatives. The statistical analysis confirmed that the mean of the RFI values for the racemic and enantiomeric derivatives were not significantly different (p>0.05) (Table II).
Since the racemic phenothiazines and their corresponding enantiomers exhibited equivalent efflux pump-inhibiting activity on the AcrAB-TolC efflux pump system of E. coli AG100, the racemic phenothiazines and their corresponding enantiomers have been evaluated as identical compounds in the following analysis concerning the energy supply of the AcrAB-TolC efflux pump system.
Because RND efflux pumps obtain their energy for activity from the proton motive force (PMF) and the PMF results from metabolic activity of the cell, the EPI effect of phenothiazines on the AcrAB-TolC efflux pump system was assessed in the presence and absence of 0.4% of glucose at pH 7.4. As expected, the RFI values of the N-hydroxyalkyl-2-aminophenothiazines are significantly lower (p<0.05) if glucose is present in the system, thereby indicating that glucose plays an important role in reducing the amount of EB accumulated by E. coli AG100 in the presence of N-hydroxyalkyl-2-aminophenothiazines at pH 7.4 (example: N-hydroxyalkyl-2-aminophenothiazine rac-3i, Figure 2). The phenothiazines rac- 3h and rac-5 showed significantly less activity in the presence of the indirect energy source glucose (Figure 2).
Statistical analyses of the relative final fluorescence index (RFI) of racemic phenothiazines (rac-3h, rac-3i, rac-5) and their (+)- and (−)-enantiomers using a one-way analysis of variance (ANOVA) complemented by the Bonferroni post-test for multiple comparisons. R2 is the fraction of the overall variance attributable to differences among the group means (R2=0.279), the F ratio is the ratio of two mean square values (F=0.8705).
Discussion
RND proteins are part of tripartite pumps and they mediate transport across the inner and outer bacterial membrane. The conformational changes and the dynamic transition of the different configurations of the AcrB protein have to be elucidated in order to design effective EPIs (12). The discovery of the inhibitor-bound structures of AcrB described by Nakashima et al. in 2013 contributed enormously to the EPI-discovery strategies related to the RND transporters (13).
From the observations presented, it can be concluded that the chirality of resolved N-hydroxyalkyl-2-aminophenothiazines does not influence the EPI activity of the derivatives on the bacterial AcrAB-TolC efflux pump system of E. coli AG100. It can be suggested that there is no direct interaction between the phenothiazine derivatives and the AcrB protein. Furthermore, they may influence the expression of genes regulating the AcrAB-TolC system (14). It was previously shown that promethazine might form a complex with two guanine–cytosine (G–C) rich regions of DNA (15). In addition, phenothiazine derivatives can cause unspecific reactions with membrane lipids because of the high degree of their lipophilicity (16).
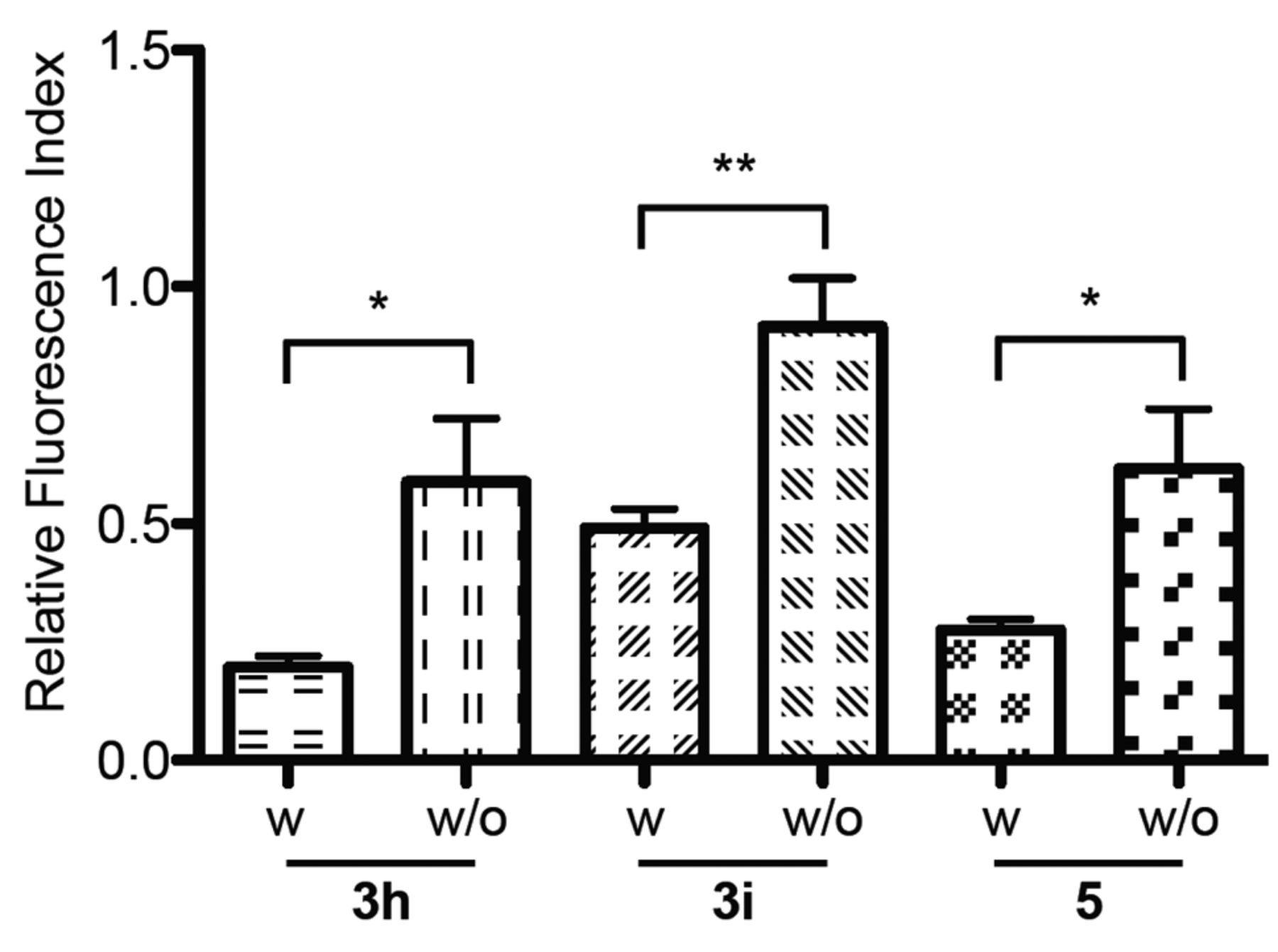
The effect of glucose on RFI index in the presence of N-hydroxyalkyl-2-aminophenothiazines: the activity of racemic phenothiazines (rac-3h, rac-3i, rac-5) and their enantiomers at 100 μM on the accumulation of EB by E. coli AG100 in the presence and absence of glucose 0.4%. Legend: w, with glucose 0.4%; w/o, without glucose 0.4%; *the correlation is significant: p<0.05; **the correlation is very significant: p<0.01.
However, if glucose is present in the system, the activity of the N-hydroxyalkyl-2-aminophenothiazines will be diminished demonstrating the role of energy supply on the activity of the bacterial AcrAB-TolC efflux pump system.
Acknowledgements
Gabriella Spengler was supported by the Campus Hungary Teaching Staff Short Term Mobility action of the Social Renewal Operational Program (TÁMOP) of Hungary.
The Authors thank Prof. Dr. Miguel Viveiros for supporting the research carried out at the Institute of Hygiene and Tropical Medicine (Lisbon, Portugal). The authors wish to thank Elisabete Junqueira for the technical help in the laboratory work. The authors gratefully acknowledge Dr. Attila Hunyadi (Institute of Pharmacognosy, University of Szeged, Szeged, Hungary) for the valuable scientific discussion.
Footnotes
-
Funding
TÁMOP-4.2.2.A-11/1/KONV-2012-0035; TÁMOP 4.2.4.B/2-11/1 2012-0001 (Hungary).
This research was supported by the European Union and the State of Hungary, co-financed by the European Social Fund in the framework of TÁMOP 4.2.4.A/2-11-1-2012-0001 ‘National Excellence Program’.
- Received July 21, 2014.
- Revision received September 19, 2014.
- Accepted September 26, 2014.
- Copyright © 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.