


Abstract
In the present study it was shown that resveratrol (3,4,5-trihydroxystilbene), an efficient light-absorbing molecule, during its transition from trans to cis configuration under UV light, transfers its energy of excitation to triplet oxygen to produce singlet oxygen (1O2). This transition is prevented by Trolox, a quencher of singlet oxygen. In the presence of a stable amount of nitrosoglutathione, UV-irradiated resveratrol reacts with nitric oxide (NO) originating from the nitrosoglutathione to produce peroxynitrite (ONOO−). Beta-carotene, acting as a quencher of 1O2, prevents the transition of resveratrol from trans to cis. Beta-carotene also prevents DNA damage induced by the 1O2. NO synthase (NOS) activity in synaptosomes isolated from rabbit brain increased approximately three-fold by resveratrol and the NO released was converted to ONOO−. Resveratrol increased the lipid fluidity of synaptosomal plasma membranes. These changes suggest that the incorporation of resveratrol into synaptosomal plasma membranes causes an up-regulation of NO synthase. On the other hand, the simultaneous ONOO− and 1O2 formation may cause disturbances in transmembrane signal transduction leading to neurotoxicity. The present study concerning the behavior of resveratrol with respect to its structure and potential prooxidant-antioxidant function provides important new clues as to the role of this fascinating molecule in pathophysiology.
- Nitric oxide
- peroxynitrite
- singlet oxygen
- resveratrol
- ultraviolet C radiation
- rabbit brain synaptosomes
- neurotoxicity
Abbreviations: NO, Nitric oxide; ONOO−, peroxynitrite; SOD, superoxide dismutase; UVC, ultraviolet C; L-NMMA, N-monomethyl-L-arginine; TMA-DPH, 1,4-trimethylamino-phenyl-6-1,3,5,hexatriene.
Trans-resveratrol (3,4,5-trihydroxy-stilbene) is a phytorelaxin, highly abundant in skins of red grapes and moderately abundant in peanuts and blueberries. Resveratrol exists as cis and trans isomers. Trans-resveratrol is the preferred steric form and is relatively stable if it is protected from light. It is of interest for its reported beneficial properties such as anticancer, anti-aging, antiviral, cardiovascular and neuroprotective effects. Literature has underscored the high degree of diversity in terms of the signaling networks and cellular effector mechanisms impacted by resveratrol. Among them are cell surface and nuclear receptors, gene transcription and metabolic pathways (1, 2).
The molecular mechanism of action of resveratrol is not entirely clear. In endothelial cells, it has been proposed to act through membrane-bound structures, for example, estrogen receptors, as well as intracellular targets, such as AMP-activated protein kinase. Many of the protective properties of resveratrol have been attributed to its ability to reduce oxidative stress. Studies have shown an increase in antioxidant enzyme activities following exposure to resveratrol, including up-regulation of mitochondrial superoxide dismutase (SOD), an enzyme that is capable of reducing both oxidative stress and cell death (3). Resveratrol also increases nitric oxide synthase (NOS) expression in various types of endothelial cells, an effect dependent on the estrogen receptors (4). Recent studies underlie peroxynitrite (ONOO−)-mediated apoptosis in endothelial cells and potential mechanisms responsible for resveratrol cytoprotective action through the prevention of ONOO−-induced caspase-3 and -9 activation (5).
Biomolecules which can exist in the ground state and in the first two (singlet and triplet) excited states, are efficient light-absorbing molecule (photosensitizer). The energy released is transferred to molecular oxygen to produce singlet oxygen (1O2), its first excited electronic state, distinguished from the triplet ground state of oxygen. 1O2 is a short-lived species known to react with a host of molecules, including many of biological significance, to yield a variety of oxygenated derivatives. These reactions can have many effects, ranging from the degradation of organic materials to cell death (6).
In the present study, it was investigated whether resveratrol, under UV light, transfers its energy of excitation to triplet oxygen to produce singlet oxygen (1O2) and thus could activate synaptosomal plasma membrane NO synthase.
Materials and Methods
Materials. Ficoll-purified synaptosomes isolated from rabbit brain cortex were prepared as previously described (6). Compared with the original homogenate from which they were prepared, they showed a 7- to 10-fold increase in the (Na+ + K+)-stimulated ATPase activity (EC 3.6.1.3). Protein concentration was measured by the Bradford method (7).
Luminol (5′amino-2,3-dihydro-3,4-pthalazinedione) and lucigenin (bis-N-methylacridinium nitrate) were purchased from Sigma Chemical Company (St. Louis, MO, USA). A stock solution (10 mM) of resveratrol was prepared in dimethylsulfoxide (DMSO). 6-Hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid (Trolox), 2-phenyl-4,4,5,5,-tetramethyl-imidazoline-1-oxyl-3-oxide (PTIO) and DNA isolated from salmon testes were purchased from Sigma. Hank's balanced salt solution (HBSS), phosphate-buffered saline (PBS) were purchased from GIBCO (Grand Island, NY, USA). N-monomethyl-L-arginine monoacetate (L-NMMA) and SOD were obtained from Calbiochem (Switzerland). Nitric oxide (NO) (99.99% pure) was obtained from Messer Griesheim (Germany).
UVC irradiation. Resveratrol dissolved in DMSO and diluted in PBS was irradiated with UVC light for 30-60 min from a distance of 10 cm, using a UVC lamp (VL-6C, 6 W-254 nm Tube, Power: 12 W (Vilber Lourmat).
High pressure liquid chromatographic (HPLC) estimation of trans-cis conversion of resveratrol. HPLC analysis of resveratrol was carried out using a VYDAC 5 μ C-18 column (250×4.6 mm) Merk Ltd. Mainz, Germany. The eluant was CH3CN (20%), CH3COOH (5%) and H2O (75%) at a flow rate of 1 ml/min and detection was by a fluorescence detector (Hewlet Packard FLD 1100 series) set at excitation at 298 nm and emission at 385 nm.
Chemiluminescence measurements (CL). Reactive oxygen species were measured by luminol-amplified chemiluminescence as previously described (8). All the light (photons) emitted were measured using a Berthold AutoLumat LB953 Luminometer (Berthold GmbH & Co, D-7547 Wildbad). A stock solution (1 mM) of luminol in DMSO was diluted in Hans' Balanced Salt Solution (HBSS) to a final concentration of 10 μM just before use. Luminol was directly injected in the tubes by the Luminometer. Light emission was recorded by the computer interface and reported as the integrated light emission for a total period of 10 to 20 s. The results were calculated at counts per min (cpm).
CL from DNA oxidized by 1O2. The 1O2 produced by UVC-irradiated resveratrol (dissolved in Phosphate Buffered Saline (PBS) pH 7.4) was detected from the CL resulted from the oxidative damage to DNA induced by 1O2. Into the 1O2 generation system, 100 μl of a series of concentrations (20-100 μg) of DNA and the antioxidant beta-carotene (100 μM), for the evaluation of its protective effect, were added. The final volume of the reaction mixture in the tubes was kept at 0.5 ml in all assays by adjusting the buffer solution volume.

Time-dependent release of oxygen free radicals from UVC irradiated resveratrol. (A) Control resveratrol and (B) resveratrol plus Trolox.
Fluorometric measurements of resveratrol. The fluorometric spectra of UVC irradiated resveratrol were obtained on a Hitachi F-2000 Fluorescence Spectrophotometer, with an excitation at 330 nm and emission at 374 nm. The scavenging activity of beta-carotene on 1O2 release from resveratrol was estimated.
Assay of NO synthase activity. Synaptosomal NO synthase isolated from rabbit brain was measured by monitoring the NO production. Reactions (1 ml sample volumes) were carried out for 1 min at 37°C. Standard reaction mixtures contained: HBSS-EDTA (1 mM) (pH 7.4); synaptosomes (10 mg of protein); L-arginine (100 μM); NADPH (100 μM); tetrahydrobiopterin (5 μM) and calmodulin (1 μM). NO determination was carried out by mixing the incubates with 100 μl of H2O2. The chemiluminescence response was obtained using the kinetics protocol of the AutoLumat Luminometer, allowing the estimation of ONOO− from the reaction between NO and H2O2. L-NMMA (1 mM) was added to the incubation mixture to inhibit NO synthase activity.
Membrane fluidity. Membrane fluidity changes in synaptosomes due to resveratrol treatment were estimated in steady-state fluorescence polarization studies of 1,4-trimethylamino-phenyl-6-1,3,5,hexatriene (TMA-DPH) according to Shinitzky and Barenholz (9) as previously described (10). Resveratrol-treated synaptosomes were mixed with TMA-DPH (10 μl) (1 mg/10 ml cyclohexane) for 15 min. Measurements were performed in a fluorophotometer equipped with polarizers for polarization studies, at excitation of 360 nm and emission of 430 nm. The fluorescence anisotropy is expressed as r=IVV-IVH/IVV+2IVH where IVV and IVH are the intensities of the emitted light oriented, parallel and perpendicularly to the plane of the exiting beam, respectively.
Results
In Figure 1, a racemic mixture of trans and cis resveratrol (10−6 M) in PBS at pH 9.2, irradiated with UVC, gave a luminol CL response at ~40 min with a peak at ~8×108 cpm. In the presence of the antioxidant Trolox (0.1 mM) the chemiluminescence spectrum is shifted to the right at ~80 min, with the peak depressed to ~4×105 cpm, clearly showing the prooxidant activity of resveratrol.
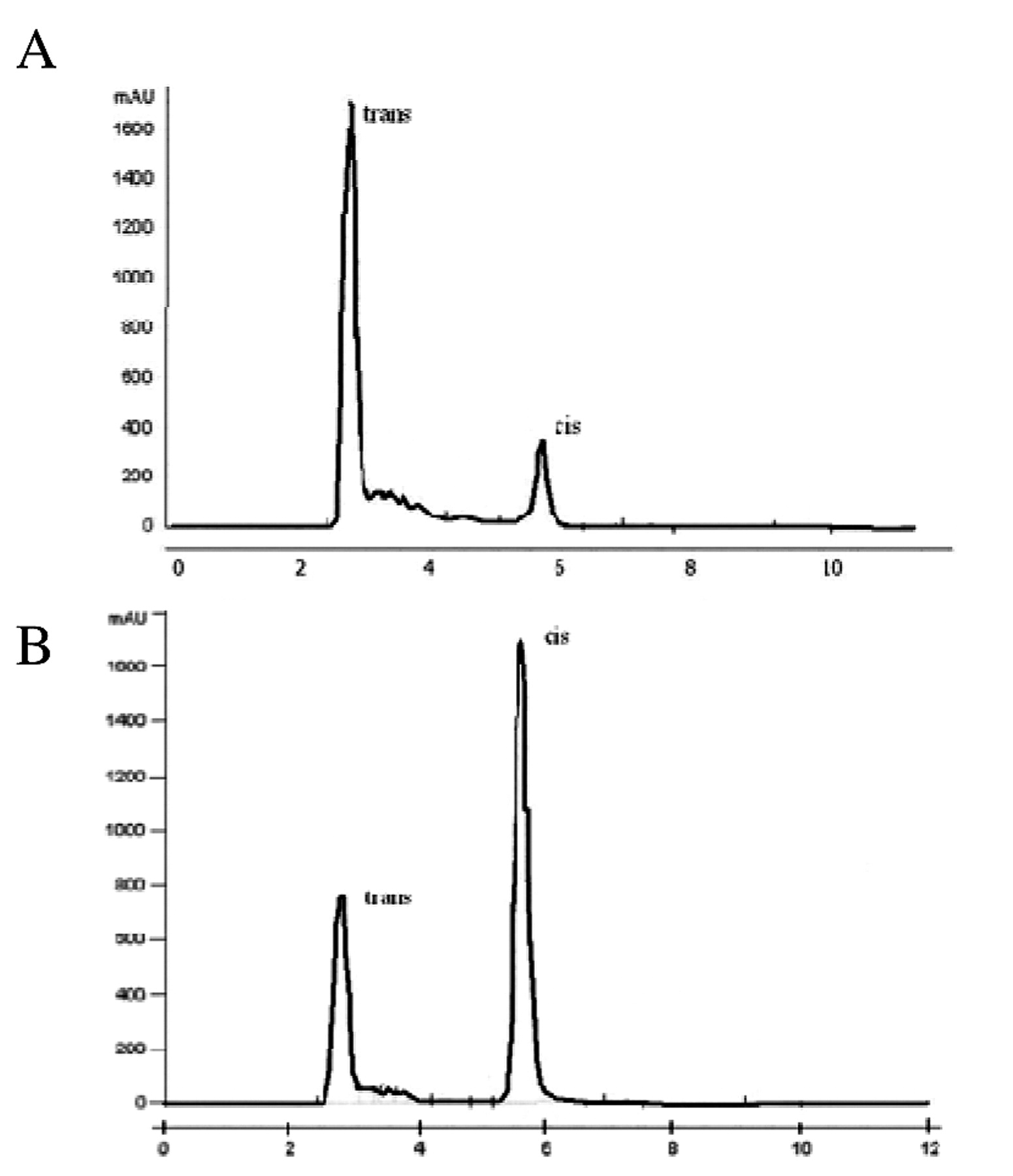
HPLC chromatograms of trans-resveratrol before (A) and (B) after exposure to UVC for 60 min.
HPLC studies allowed the quick and efficient separation of trans- and cis resveratrol (Figure 2A). Trans- resveratrol was eluted first with a retention time of 2.8 min, followed by cis-resveratrol at 5.6 min, which indicates a noticeable difference in the hydrophobicity of these two compounds. The highest peak at the retention time of 2.8 min was obtained from solutions of pure trans-resveratrol. After UVC irradiation of resveratrol (Figure 2B), the highest peak was obtained at the retention time of 2.8 min corresponding to cis resveratrol.
Figure 3A shows the ability of carboxy-PTIO to scavenge the NO released from nitrosoglutathione. A standard amount of nitrosoglutathione (10−6 M) at PBS pH 9.2 was mixed with various amounts of carboxy-PTIO, which reacts rapidly with NO to yield the corresponding imidazoli-neoxyl and free NO2 radical. The decrease in the CL follows a hyperbolic curve. However, as shown in Figure 3B, when carboxy-PTIO was replaced by resveratrol, a sigmoidal increase in chemiluminescence was observed due to its prooxidant activity. Our present experiments have also shown that resveratrol is capable of producing free radicals that react with NO released from nitrosoglutathione to form peroxynitrite (ONOO−) (Figure 3B).
Figure 4 shows the 1O2 production by UVC irradiated resveratrol (dissolved in PBS pH 7.4). Tmax is the time at which the peak chemiluminescence was obtained when 1O2 reacted with DNA to form reactive species. The CL response (expressed as Tmax) due to the oxidative damage caused to DNA by 1O2 was obtained from a 1O2 generation system consisting of UVC-irradiated resveratrol (10 μM) and different concentrations of DNA (20, 50 and 100 μg) in the presence and absence of the antioxidant beta-carotene. Mannitol (10 mM) and SOD (100 U) were ineffective at protecting the DNA oxidized by 1O2, suggesting that hydroxyl radicals and/or superoxide were not involved in these oxidative reactions.

A) Scavenging activity of carboxy-PTIO on NO release from nitrosoglutathione. B) Dose-dependent increase of NO release from nitrosoglutathione in the presence of resveratrol.
1O2 reacted with DNA to form reactive species which then turned into secondary radicals, especially alcoxyl radicals (oxyl and peroxyl). The secondary radicals formed triplet carbonyl compounds again by their recombination giving the chemiluminescence response.
Membrane lipid fluidity determines the optimal membrane functions due to the physical constrains imposed by lipids in membrane proteins (for example enzymes, pumps, channels). Studies on the effects of various substances on membrane fluidity using the fluorescence polarization (r) of diphenylhexatriene (DPH) showed that an increase in fluidity produces concomitant diminution in the r values. In the present study, the r value of TMA-DPH incorporated in rabbit brain synaptosomes decreased in the presence of resveratrol (10 μM) from 0.205±0.025 (controls) to 0.157±0.015, indicating an increase in membrane fluidity.
The fluorometric spectra of UVC-irradiated resveratrol (10 μM) dissolved in PBS pH 7.4 (ex. 330 nm, em. 374 nm) are shown in Figure 5. The scavenging activity of beta-carotene (100 μM) on 1O2 release from resveratrol was idendified.

Dose-dependent chemiluminescence response of DNA oxidized by 1O2 produced from UVC irradiated resveratrol (Resv). A, Resv+DNA (20 μg); B, Resv+DNA (50 μg); C, Resv+DNA (100 μg); D, Resv+DNA (100 μg)+beta-carotene (100 μM).
Figure 6 shows NO release from rabbit brain synaptosomes (1 mg of synaptosomal protein) treated with UVC irradiated resveratrol (10 μM). The NO production was increased approximately 3-fold by resveratrol, which was inhibited by approximately 70% in the presence of L-NMMA (1 mM).
Discussion
In the present study it was shown that resveratrol (3,4,5-trihydroxystilbene) which can exist in the ground state and in the first two (singlet and triplet) excited states, is an efficient light-absorbing molecule (photosensitizer). During its transition from the trans to the cis configuration, under UVC light, resveratrol transfers its energy of excitation to triplet oxygen to produce oxygen free radicals (Figures 1, 2 and 3). More specifically, after exposure to doses of UVC energy, it evokes singlet oxygen (1O2) formation that is sustained for more than 60 min (Figure 4). These results indicate that resveratrol, which is known to be an effective antioxidant agent (11), may be amenable to suitable chemical modifications by appropriate charge density changes in specific sites and thus to became important in determining signaling cascades and intracellular signals. Although the majority of studies of pharmacological effects have been concerned with trans resveratrol it has also been shown that the cis isomer significantly attenuates several components of the macrophage response to proinflammatory stimuli (production of superoxide, NO and PGE2) (12).
The release of free radicals from UVC-irradiated trans resveratrol was demonstrated by the ability of antioxidants such as Trolox to quench luminol chemiluminescence intensity. The chemiluminescence spectrum obtained from UVC-treated resveratrol was enhanced compared to the non-treated control, indicating its enhanced capability for causing a higher oxidative stress to the molecules with which it interacts (Figure 1). This ability of resveratrol to generate free radicals has been suggested to be responsible for its cytotoxic effects. The inhibitory effect of free Cu(II) metal-chelators on such reactions indicates a role for Cu(II) in this process of oxygen-free radical production. The latter has been postulated to require the redox cycle of Cu(II)-peroxide complex capable of binding DNA to form a DNA-resveratrol-Cu(II) ternary complex (13). In our present study, in addition to the generation of 1O2 by UVC-irradiated resveratrol, nitric-oxide (NO) synthase activation was evoked with NO formation in rabbit brain synaptosomes. Resveratrol activated NO synthase leading to the oxidation of NO to ONOO− (Figure 6). ONOO− binds to heme centers and forms the basis of NO-mediated signaling, as occurs through soluble guanylate cyclase, and the nitrosyl-heme may then act as an antioxidant to retard damage induced by free radicals produced after UVC radiation (14).
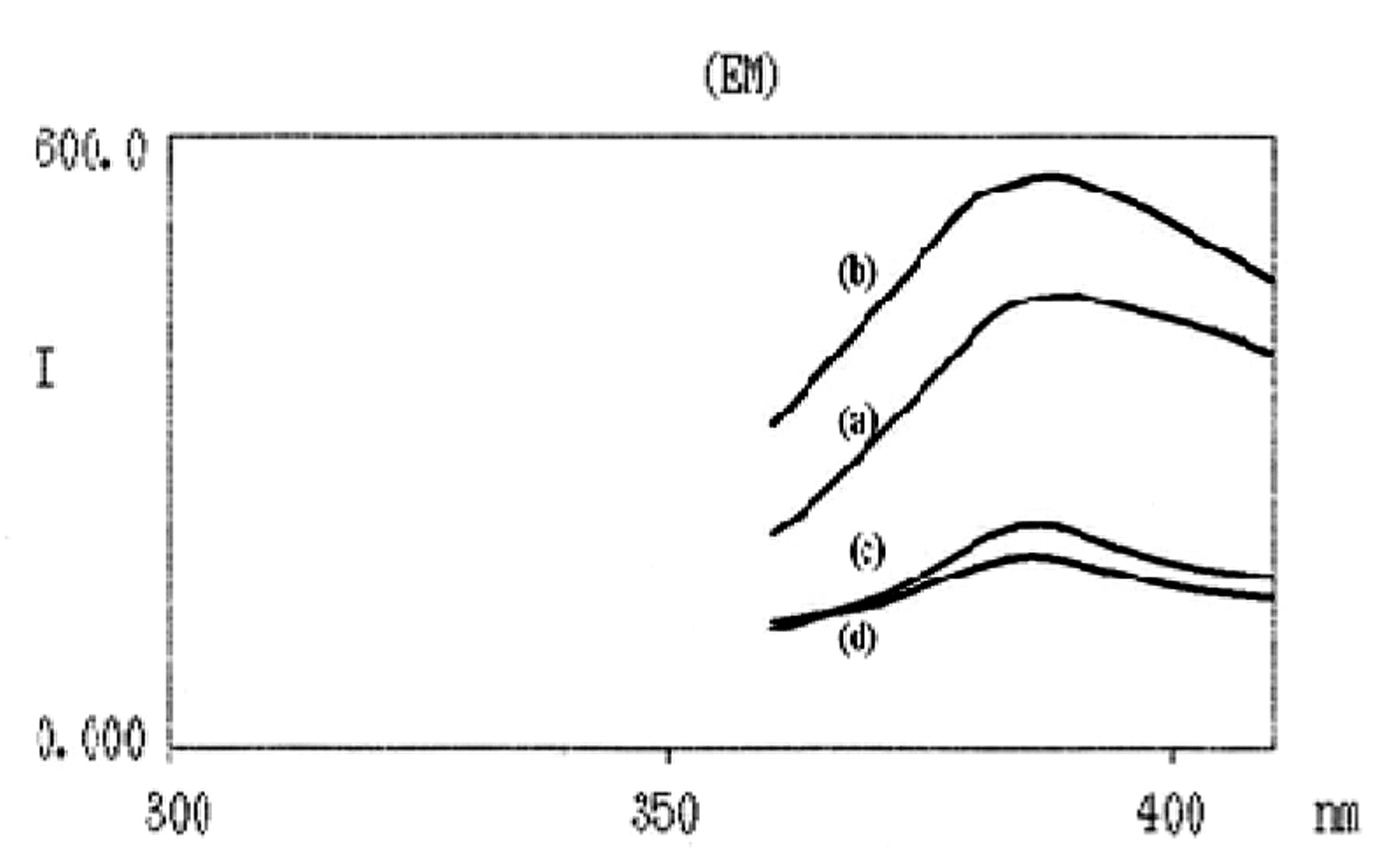
Fluorescence spectrum of UVC irradiated resveratrol: (a) 30 min after irradiation, (b) 60 min after irradiation, (c) 30 min after irradiation plus beta-carotene and (d) 60 min after irradiation plus beta-carotene.

Chemiluminescence response of NO release from (a) rabbit brain synaptosomes treated with UVC irradiated resveratrol and (b) control rabbit brain synaptosomes.
Recent data on the role of NO in disease has stimulated efforts to develop therapeutic strategies include NO donors that directly or indirectly release NO and agents that increase NO bioactivity. Direct NO donors include NO gas, which is useful in respiratory disorders, and the more recent classes of diazeniumdiolates, sydnonimines and S-nitrosothiols (15). 1O2 formed progressively during UVC irradiation of resveratrol can serve as a store of prooxidants with a concordant activation of intracellular signaling pathways (16). Therefore it is apparent that despite its well-known antioxidant properties, the resveratrol molecule has the potential to act as a cytotoxic agent.
Membrane-bound enzymes and carriers can be markedly affected by membrane lipid composition and fluidity, confirming that the physical state of the lipids surrounding various membrane proteins can control protein conformation and regulates protein function (17). Thus, optimal membrane function seems to depend on the fluidity of the constituent membrane components, especially of the membrane lipids. The effects of fluidizers on the fluorescence polarization (r) of DPH in cell membranes were demonstrated by the diminution in the r value indicating an increase in the membrane fluidity. Due to the structural similarity between resveratrol and estrogens (18), experiments were conducted to study possible effects of resveratrol on membrane fluidity. An increase of the r value was observed when rabbit brain synaptosomes were treated with UVC-irradiated resveratrol supporting the assumption that resveratrol can itself be incorporated in the lipid bilayer thus increasing membrane fluidity despite the fact that the 1O2 formation has the potential to decrease it due to lipid peroxidation and malonyldialdehyde (MDA) production (19). Membrane lipid bilayer fluidity has a crucial role in signal transduction for a variety of biologically active molecules which activate cellular functions, cell differentiation and proliferation (20).
The present data suggest that resveratrol, which is an efficient light-absorbing molecule under UV light, transfers its energy of excitation to triplet oxygen to produce singlet oxygen (1O2). The DNA damage induced by the 1O2, which was prevented by beta-carotene, further support the suggestion that resveratrol after UVC irradiation forms 1O2. Furthermore, resveratrol caused an up-regulation of the rabbit brain synaptosomal NO-synthase with a simultaneous ONOO− and 1O2 formation. In conclusion, resveratrol could act as a prooxidant by producing 1O2, with subsequent lipid peroxidation and alterations in membrane lipid fluidity, causing disturbances in extracellular signal transduction. The combination of NO with 1O2 may also form ONOO− that could cause extensive oxidative damage in brain cells leading to neurotoxicity.
Footnotes
- Received July 26, 2009.
- Revision received October 2, 2009.
- Accepted October 12, 2009.
- Copyright © 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.